一种提高实验重复性的大豆胞囊线虫抗性鉴定方法
2022-10-10练云周扬雷晨芳武永康李海朝王仕伟王金社卢为国
练云, 周扬,2, 雷晨芳, 武永康, 李海朝, 王仕伟, 王金社, 卢为国*
(1.河南省作物分子育种研究院国家大豆改良中心郑州分中心 农业农村部黄淮海油料作物重点实验室河南省油料作物遗传改良重点实验室,河南 郑州 450002; 2.漯河市农业科学院,河南 漯河 462000)
大豆是我国主要的经济作物之一,随着社会经济的发展和人们生活水平的提高,我国大豆需求量近年急剧增加。近年来,国内大豆胞囊线虫病(soybean cyst nematode,SCN)、症青、根腐病等病害对大豆造成严重减产、甚至绝产[1-3],因此,培育抗病、优质、高产大豆品种迫在眉睫。大豆胞囊线虫病是一种世界性大豆病害,世界范围内各个大豆产区均有发生,伴随近年来关于大豆产区SCN优势生理小种致病性的升级[4-6],具有超强致病力的大豆胞囊线虫新小种被发现的报道[7]越来越多,应对SCN病害措施的相关研究有待加强。SCN相关研究中的抗性资源鉴定、抗病品种培育、生理小种鉴定为提出应对SCN策略提供技术支撑[1,8-10],而以上研究均是以胞囊指数为判定依据,因此,准确、可靠的抗性鉴定方法尤其重要。尽管SCN抗性鉴定相关研究工作多是在温室内开展,但在实际操作过程中,由于胞囊受温度、湿度影响较大,不同重复间胞囊数目差异大仍是十分普遍的现象。基于通过控制小环境提高大豆胞囊线虫侵染效率的抗性鉴定对SCN研究工作具有重要意义。
已有研究表明,大豆胞囊线虫优势生理小种出现了致病能力升级的现象,比如在2016年报道的中国大豆主产区—黄淮地区和美国密苏里大豆产区,优势生理小种均由1号生理小种(1号生理小种仅能侵染Riggs鉴定模式中的PI 88788)升级到了2号生理小种(2号生理小种能侵染Riggs鉴定模式中的Peking、Pickett和PI88788 3个寄主)[11];在中国,前期还发现了具有超强致病力的新小种X12(X12小种能够侵染目前常用的所有抗病资源)[7,12],且大豆胞囊线虫仍在扩散[13]。因此,亟需一套稳定的SCN抗性资源鉴定方法,筛选新抗源并将筛选到的抗源有效用于生产实践。近年来,在SCN表型鉴定方面,取得了一定的研究进展,尤其是在计数大豆胞囊的方法方面。2005年报道了一种基于荧光成像系统的胞囊计数方法[14],可将胞囊拍成照片,输入电脑后在电脑上人工计数,此方法不需要在胞囊成熟一周内就完成对胞囊的计数,把人们从统计胞囊的时间限制中解脱了出来,但是不适于高通量计数;2010年报道了一种适于高通量计数胞囊的计数方法[15],利用一种组装、改造的荧光成像系统并配合计数软件,实现了一次拍摄6棵植株上的胞囊并自动计数的高通量统计方法,但该机器功能单一,部分配件需特殊定制,有些配件目前市场上难以采购且市场需求量小,至今在SCN研究领域未得到推广应用;2014年,报道了植物病虫害表型数据采集系统[16],此方法不需要荧光环境,直接把从根系上冲洗下来的胞囊均匀分散在黑色背景布上,利用计数软件对普通相机拍摄的胞囊图像计数,但是根系、沙子、蛭石、胞囊黏连等情况会影响计数结果;2018年报道了褐化大豆胞囊自动计数方法[17],该方法适于统计褐化胞囊数目,但是在抗性鉴定及资源筛选中,多是统计褐化前处于显囊盛期的胞囊数目。以上技术更新,均提升了胞囊数目统计的准确性及效率,已经不同程度地被应用到了大豆胞囊抗性鉴定过程中。
rhg1和Rhg4是抗SCN的2个主效位点[18-20],借助与rhg1和Rhg4紧密连锁的KASP标记[21-22],在现有大豆资源中筛选到了含有SCN抗性位点且农艺性状优良的资源[8]。开发的KASP标记不仅能从资源中筛选出来自rhg1、Rhg4位点的SCN抗性基因型信息,而且能够从绝大部分抗性资源中区分Peking类型和PI88788类型,这些标记用于高通量分子标记辅助育种[1,8,21-22]。筛选SCN抗性种质资源主要依靠在温室内开展表型鉴定,其中最重要的表型依据就是接种一定浓度虫卵后,统计根系上的胞囊数目,并对待鉴定材料抗性水平进行划分。寄生在根系中的线虫容易受到温度、湿度的影响,接种操作不规范、接种浓度不一致以及接种时虫卵活性的干扰等因素都会导致抗性鉴定需要多次重复。由于诸多因素的干扰,在温室内开展的表型抗性鉴定,劳动强度大、运行成本高,导致抗性资源筛选效率仍然比较低。目前已有的改进措施多是基于提高统计胞囊数目的准确性,目前尚未见到在抗性鉴定过程中提高大豆胞囊线虫侵染效率、提升有效重复的表型鉴定技术。
开展SCN相关工作的研究在大环境上多利用培养室进行控温、控湿,但在实际操作中,由于散热性差、线虫侵染效率不一致等小环境的影响,往往重复性差。针对温室内由小环境引起的温度、湿度、侵染效率不一致等引起的误差,我们研究一种通过调控小环境和提高大豆胞囊线虫侵染效率的抗性鉴定方法,提高接种植株间的有效重复。
1 材料与方法
1.1 实验材料与培养条件
感病材料Lee和具有不同抗性水平的大豆材料来自河南省作物分子育种研究院种质资源保存室。温室培养条件为光照16 h/8 h(白天/黑夜),温度(27±1) ℃/(24±1) ℃(白天/黑夜),湿度(70±5)%。
1.2 胞囊的繁殖
胞囊的繁殖参考练云等[5]实验方法,将受大豆胞囊线虫感染的病土分装到一次性塑料杯中,移栽子叶出土期(萌发3~5 d)的Lee幼苗至病土中,每杯1株。温室培养,适时浇水管理。
1.3 控温改善
降低筐高:将规格为638 mm×420 mm×139 mm(内部尺寸长×宽×高)的筐改用为530 mm×341 mm×70 mm(内部尺寸长×宽×高)。加湿:在筐底部垫4 cm高的底滤网,并于光照时间在底滤网里注入0.3~0.5 cm深的水,用于光照时通过蒸发达到局部降温,而在黑夜温度较低时尽量使筐底部干燥,同时防止根系伸长至筐底的水中。
1.4 纯化胞囊
将在病土中生长约25 d的Lee从病土中倒出,采用淘洗、过筛法依次用25目、60目、100目筛网从根系上分离胞囊,并搜集胞囊至100目筛网上。纯化步骤如下:依次用100目、200目、500目筛网,用橡皮塞破碎胞囊,搜集虫卵至500目筛网上,借助洗瓶和漏斗,将虫卵转移至50 mL离心管中,体积控制在20 mL以内;加等体积的40%蔗糖溶液,轻轻颠倒混匀;2 000 r·min-1离心5 min,如看到分层,则吸取中间层,弃掉上清和底部沉淀;若看不到分层,则吸取上清,弃掉底部沉淀,并将吸取的上清液在600目筛网上过滤,用流水冲洗以去除蔗糖,虫卵留在600目筛网上;借助洗瓶和漏斗,将虫卵转移至烧杯中,稀释到一定体积,混匀后取10 μL在显微镜下计数,即为接种液。
1.5 接种孔的孔径改善
紧挨着待鉴定材料根系旁边扎孔,用于注入接种液,原方法使用1 mL枪头扎孔,本研究改用3 mm×75 mm螺丝刀或3 mm直径的竹签扎孔,孔的直径是原来的1/3~1/6,接种分两次进行,每次每株接种约1 250个虫卵,每株两次共接种2 500个虫卵,两次接种间隔24 h。
1.6 统计分析
在条件改善前后,各待鉴定植株上胞囊数目取平均值,利用方差分析不同重复间的数值,统计数值变化区间,比较条件改善前后重复实验效果。
2 结果与分析
2.1 控温改善
传统方法中的筐内温度31.9 ℃、湿度49%(图1),培养架表面温度41.3 ℃、湿度28%,改进后方法(降低了筐的高度,铺设底滤网且加水)的筐内温度27.6 ℃、湿度68%(表1)。胞囊最佳生长条件为白天(27±1)℃,夜晚(24±1)℃,湿度70%±5%,改进后的条件更接近胞囊的最佳生长条件。

A—传统方法的筐内温、湿度;B—培养架表面温、湿度;C—改进后方法的筐内温、湿度。
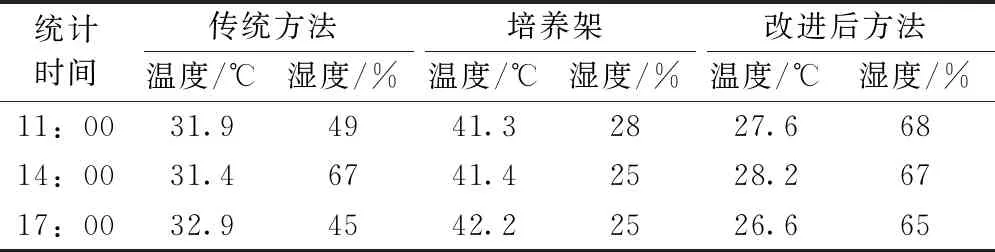
表1 在不同时间点的温、湿度比较
2.2 蔗糖梯度离心纯化接种虫卵
未经蔗糖梯度离心的虫卵溶液中有破碎的根系、沙子、土壤等杂质(图2),影响计数的准确性,经过纯化后的虫卵,在4倍显微镜下杂质明显减少,计数相对准确,接种到每一个待鉴定材料里的虫卵数目一致性更高。
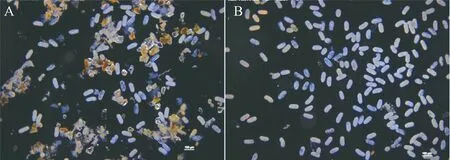
A—未经蔗糖梯度离心的虫卵;B—经过蔗糖梯度离心的虫卵。
2.3 接种孔径改善
传统方法中,使用1 mL的枪头扎孔,3 000个虫卵注入一个孔内,因枪头上部直径较大,扎入土内导致杯内土壤产生裂纹,接种液易扩散到远离根系的地方,导致孵化后的幼虫不能及时接触到寄主;改善后,注入接种液的孔直径是原来的1/3~1/6,由于扎孔的孔径小,不易在扎孔时产生裂纹,而且接种液分两次注入,每次注入一个孔内(图3)。条件改善后有效缩小了虫卵与根系的接触空间,孵化后的幼虫更容易接触到寄主。

A—用直径为5 mm的竹签扎孔;B—用直径为3 mm的螺丝刀扎孔;C—用1 mL移液器枪头扎孔。
传统方法中,每株一次接种3 000个虫卵,条件改善后,每株一次接种1 500个虫卵,接种两次。采用分两次接种可以减少因虫卵活性差异或人为漏接种引起的误差。改用螺丝刀/竹签扎孔后,由于孔径小,不容易注入1 mL接种物,借用1 mL枪头辅助接种。
2.4 分离胞囊并计数
培养25 d后,将大豆植株从病土中倒出,采用淘洗、过筛法从根系中分离胞囊,利用计数软件及体式显微镜,通过计算机扫描、分析,自动计数每张照片上的大豆胞囊数目,计数软件具体使用方法见专利2018SR477328[16]。
通过以上在温湿度、纯化接种虫卵、接种方式等方面的改善,使得每个重复实验结果之间的胞囊数目差异较小,标准差减小(表2)。从条件优化前后接种的大豆材料Lee来看,条件优化后,标准差由133.9减小到25.1,数值变化范围由35~347优化为102~166。条件优化后,不同重复间标准差和数值变化范围均有较明显变化(表1)。
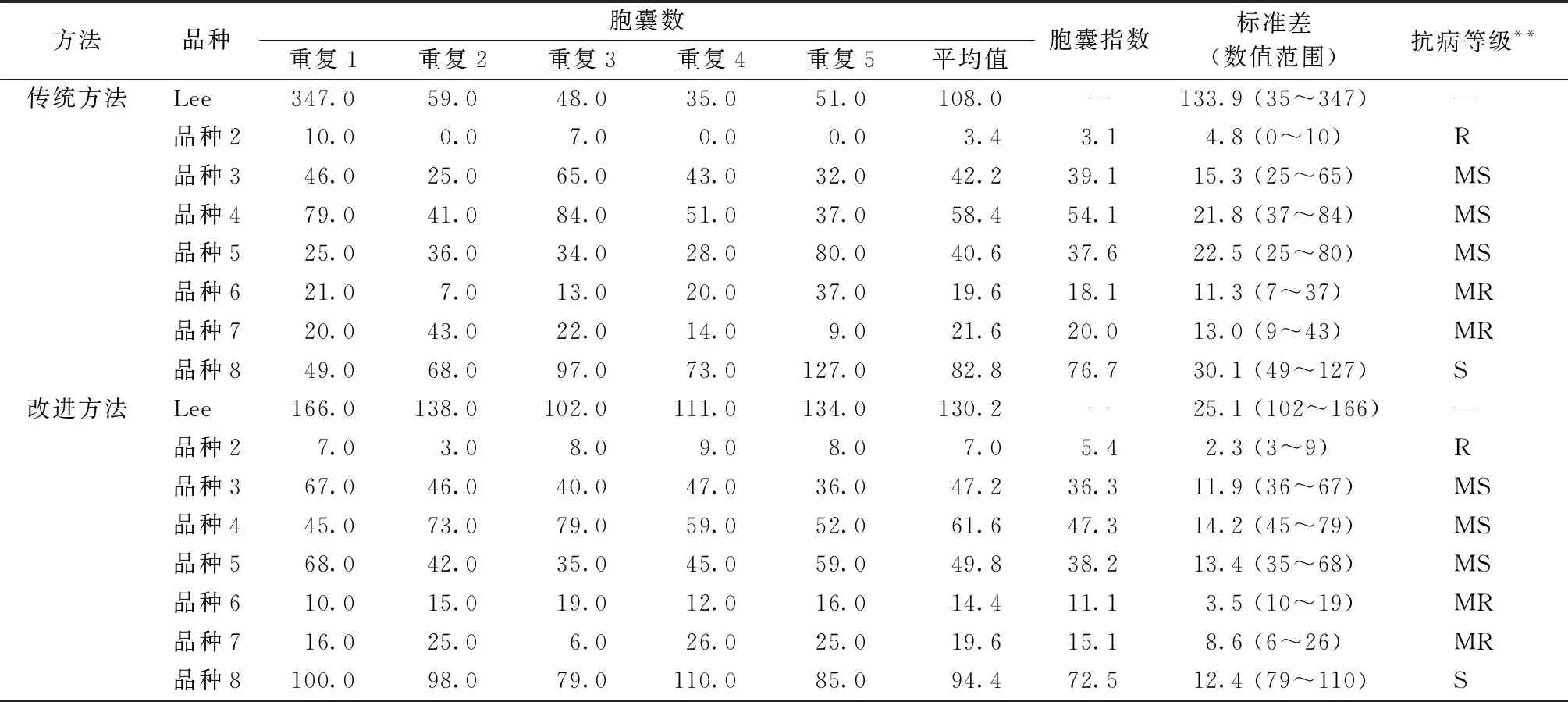
表2 大豆胞囊线虫抗性鉴定结果比较
从表2可以看出,采用改善条件后的方法使得每个重复胞囊数目一致性较好,标准差减小。其次,在鉴定过程中,利用传统的鉴定方法,会遇到初次鉴定为中感或中抗的品种,但在重复鉴定后分别是感病或中感的现象。使用传统方法对鉴定为中抗、中感的品种需要进行2~4次重复,对于鉴定为免疫和抗病的材料,需要进行4次或更多次重复才能下结论。采用改进后的方法,鉴定结果相对稳定,对鉴定为中感的材料一般不再重复,对鉴定为免疫、抗病、中抗的材料,根据实验要求重复1~2次即可下结论。
依据传统方法操作的抗性鉴定,由于参试材料重复间数值差异大,常常需要多次重复才能确定抗性。推测重复间数值差异大的可能原因是同一个品种间接种浓度不一致、温度差异大。因此,为保证鉴定结果的准确性,每一个品种的鉴定都会重复3次以上,尤其是对于抗性材料,需在不同时间多次重复,才能下结论。这也是大豆胞囊线虫抗性鉴定工作进展缓慢的原因之一,而经过对控温、纯化胞囊、接种方式、接种孔隙等关键步骤的优化,有效提高了抗性鉴定接种效率,使得重复间数值差异明显降低。这一技术的改进对推进大豆胞囊线虫相关研究,包括抗性资源筛选、大豆胞囊线虫生理小种鉴定、大豆胞囊线虫抗病基因挖掘、大豆胞囊线虫抗病品种培育及大豆-大豆胞囊线虫互作研究有极大的促进作用。
3 小结与讨论
随着科技进步和科研条件改善,较多单位能够在培养室有效控制温度和湿度,开展SCN相关研究[9,23-26]。但多年抗性鉴定结果表明,由于SCN易受温度、湿度的影响,同一批次重复间的胞囊数目差异较大,导致SCN表型鉴定需要多次重复。Zhou等[27-28]在温室内循环水浴条件下培养大豆胞囊线虫,能够较好地控制温度,并开展大豆胞囊QTL定位研究。目前,国内还未见用循环水浴培养大豆胞囊线虫的报道,多是在温室培养架上进行培养,还有一些抗性鉴定是利用自然温度在田间病圃开展,温度、湿度较难控制。
本研究降低筐的高度,有助于散热,保证杯子间温度一致;垫底滤网并在筐底部加少量的水,有助于散热,温度可控制在光照时间(27±1)℃、夜晚时间(24±1)℃,且能够有效避免根系通过杯子底部的孔伸长到水里,并保证湿度在70%±5%。筐的尺寸改善前,每个筐内可放6×9个杯子,每个种质材料重复3~6次;筐的尺寸改善后,每个筐内可放4×8个杯子,每个种质材料重复4次,使得设计实验间的重复更加方便。
据了解,国内目前在SCN抗性鉴定中,室内接种条件多不经过纯化,将胞囊收集,用研钵轻轻研磨把胞囊破碎后,将释放出来的卵冲洗入烧杯内,用清水将虫卵悬浮,取10 μL在显微镜下计数虫卵密度后,直接用于接种。在直接用于接种的条件下,由于虫卵溶液里含有砂土、蛭石、根系等残留物,接种物比较脏,计数欠准确。经过纯化程序的虫卵,由于经过利用蔗糖梯度离心之后,去除了显微镜下可视的残留物,虫卵比较干净,计数相对准确。另外,孔径减小后,有效缩小了虫卵与根系的接触空间,孵化后的幼虫更容易找到寄主;而且有效减小了扎孔时根系周围出现裂纹的现象,更容易让接种的虫卵聚集在根系周围,采用两次接种可以有效避免因虫卵活性不好或一次鉴定量大时人为漏接种导致的实验误差。本研究结果可以为开展SCN相关研究提供参考依据。
