长链非编码RNA肝癌高表达转录本通过微小RNA-134-5p促进宫颈癌细胞恶性生物学行为的机制研究
2022-09-22谭建福饶丽娟王秀
谭建福,饶丽娟,王秀
宫颈癌是常见的妇科恶性肿瘤之一,在全世界女性癌症死亡原因占很大比例。据报道,全球每年有超过520 000名新增宫颈癌病人,每年因宫颈癌死亡人数超过270 000[1]。由于巴氏涂片筛查试验的广泛实施,宫颈癌的发病率和死亡率在过去30年内有所下降,但晚期宫颈癌病人的预后仍较差,5年生存率约为15%[2]。因此,迫切需要深入了解宫颈癌发生和发展的分子机制,为确定新的诊断和预后标志物提供基础。长链非编码RNA(long noncoding RNAs,lncRNAs)是一种不具有编码蛋白质能力的RNA,长度>200个核苷酸,可通过转录调控,转录后调控,在基因表达和细胞生物学功能中发挥作用[3-4]。大量研究表明lncRNAs与包括宫颈癌在内的多种癌症的发生和发展密切相关[5-6]。然而,目前关于宫颈癌发病和进展的LncRNAs较匮乏,以往报道显示,lncRNA肝癌高表达转录本(highly upregulated in liver cancer,HULC)可促进胃癌、肝癌和骨肉瘤等肿瘤恶性进展,发挥致癌基因的功能[7-9]。近期研究发现,lncRNA HULC在宫颈癌组织中表达量较高,但其是否参与宫颈癌细胞恶性生物学行为,目前未见相关报道[10]。目前研究发现,HULC与下游微小RNA(microRNAs,miRNA)存在作用靶点[11],本研究前期通过生物信息学软件预测发现,HULC与miR-134-5p间存在相似结合位点。miR-134-5p可调控宫颈癌细胞增殖过程,具有一定抑癌作用。故本研究推测HULC可通过抑制miR-134-5p的表达参与宫颈癌的进展和预后。因此,本研究于2019年10月至2020年5月,探究HULC和miR-134-5p间的调控关系及对宫颈癌细胞生物学行为的影响,以期为宫颈癌的诊断提供潜在的标志物。
1 材料与方法
1.1 材料人正常宫颈上皮细胞Ect1/E6E7及宫颈癌细胞HeLa、Caski、SiHa、C-33A购自美国ATCC;DMEM培养基购自美国Gibco公司;特异性靶向HULC siRNA和非特异性siRNA、miR-134-5p模拟物(miR-134-5p mimics)、模拟物对照(mimics control)、miR-134-5p抑制剂(miR-134-5p inhibitors)、抑制剂物对照(inhibitors control)均购自广州锐博;Trizol试剂、M-MLV逆转录试剂盒购自大连宝生物工程有限公司;脂质体转染试剂Lipofectamine 2000购自美国Invitrogen公司;SYBR Green PCR荧光检测试剂盒购自武汉塞维尔生物科技有限公司;双荧光素酶报告基因检测试剂盒购自美国Promega公司。Transwell小室购自美国Coring公司;Matrigel基质胶购自美国BD公司;Annexin V-FITC/PI细胞凋亡检测试剂盒购自日本同仁化学研究所。本研究符合《世界医学协会赫尔辛基宣言》相关要求。
1.2 细胞培养及分组将宫颈癌C-33A细胞培养在含10%胎牛血清(FBS)、100 U/mL青-链霉素的DMEM培养基中,放置在体积分数为5%二氧化碳、37℃恒温培养箱中孵育,孵育24 h后更换培养基,稳定孵育48 h,细胞贴壁生长汇合度达80%以上时,用胰蛋白酶消化传代,取对数增殖期的细胞进行后续转染实验。将生长状态良好的C-33A细胞接种到96孔板中,继续孵育24 h,待细胞生长汇合约50%时,将特异性HULC siRNA和非特异性siRNA用Lipofectamine 2000转染至C-33A细胞中,分别命名为si-HULC组和NC组,以未转染的C-33A细胞命名为Control组,转染后的细胞放置在37℃培养箱继续培养。特异性HULC siRNA序列:正向-5’-CCGGAAUAUUCUUUGUUUAUU-3’;反 向-5’-UAAACAAAGAAUAUUCCGGUU-3’;非 特异性siRNA序列:正向-5’-CCUUAUAUGUUCUGGAAUUUU-3’;反向-5’-UAAAACGAAUGGAAUUCACUU-3’。
1.3 qRT-PCR检测细胞中HULC的表达采用Trizol法从正常宫颈上皮细胞Ect1/E6E7和宫颈癌细胞HeLa、Caski、SiHa、C-33A中提取RNA,si-HULC组和NC组C-33A细胞转染48 h后,同样以Trizol法提取细胞中RNA。使用超微量核酸蛋白检测仪测定RNA的浓度和纯度,选取合格的RNA进行逆转录,合成cDNA,通过SYBR Green PCR荧光检测试剂盒测定细胞中HULC相对表达水平,以18S rRNA作为内部对照,反应条件为:95℃预变性10 min,95℃变性15 s,55℃退火30 s,72℃延伸20 s,采用40个循环。反应结束后分析熔解曲线,得出Ct值,采用2-ΔΔCt公式计算细胞中HULC相对表达量,实验重复3次,取均值。引物:18S rRNA:正向引物5’-AGGATCCATTGGAGGGCAAGT-3’;反向引物5’-TCCAACTACGAGCTTTTTAACTGCA-3’;HULC正向引物5’-TCATGATGGAATTGGAGCCTT-3’;反向引物5′-CTCTTCCTGGCTTGCAGATTG-3’。
1.4 CCK-8检测C-33A细胞的增殖能力将各组C-33A细胞用胰蛋白酶消化,制成单细胞悬液,以1×103个/孔接种到24孔细胞培养板中,每组设置3个平行复孔,置37℃培养箱继续培养48 h,加入20 μL CCK-8,37℃孵育1 h,在酶标仪上吸光度值(OD值),波长设为测定450 nm。
1.5 流式细胞术收集各组C-33A细胞,以PBS洗涤2次,加入适量1×Binding Buffer悬浮细胞并调整细胞至1×106个/毫升,向细胞悬液中添加5 μL Annexin V-FITC和5 μL PI,避光反应15 min,补加1×Binding Buffer缓冲液,1 h内上流式细胞仪检测。
1.6 Transwell实验将Transwell小室置于24孔细胞培养板中,在小室的上室中加入50 μL以不含血清培养基稀释的Matrigel基质胶,待基质胶凝固后备用。Transwell小室的下室添加600 μL含15%胎牛血清的培养基,收集各组细胞计数,取约1×104个细胞加入到Transwell小室的上室(侵袭实验上室基底膜以Matrigel基质胶包被,迁移实验无需Matrigel包被),转移小室于37℃培养箱48 h后取出,拭去上室细胞,用0.1%结晶紫染色15 min,洗去染色,晾干后在显微镜下(随机选5个视野)观察并计数。
1.7 双荧光素酶报告基因实验生物信息学软件LncBase Experimental v.2预 测结果 显示,HULC与miR-134-5p存在相似结合位点,为进一步验证HULC能够靶向作用于miR-134-5p,将miR-134-5p与HULC结合位点的3’UTR扩增并构建到荧光素酶报告基因载体上,记为HULC-Wt,并将miR-134-5p与HULC乱序结合位点的3’UTR构建到荧光素酶报告基因载体上,记为HULC-Mut。将HULC-Wt和HULC-Mut与miR-134-5p mimics或mimics control共同转染到C-33A细胞中,48 h后使用双荧光素酶报告基因检测试剂盒分析各组细胞的相对荧光素酶活性。
1.8 转染miR-135-5p inhibitor逆转实验为进一步验证HULC靶向miR-134-5p调控宫颈癌细胞增殖、侵袭和迁移能力,本研究在抑制HULC表达的C-33A细胞中 转染miR-134-5p inhibitor或inhibitor对照,分别记为si-HULC+anti-NC组和si-HULC+antimiR-134-5p组,采用qRT-PCR技术检测miR-134-5p的表达水平,miR-134-5p正向引物5’-ATCTGTGACTGGTTGACCAGAGG-3’;反 向 引 物5’-TGCAGGGTCCGAGGT-3’;U48正向引物5’-AGTGATGATGACCCCAGGTAACTC-3’;反向引物5′-CTGCGGTGATGGCATCAG-3’。采用CCK-8检测、流式细胞术和Transwell实验分析细胞增殖、凋亡、侵袭和迁移能力,步骤同上。
1.9 统计学方法采用SPSS 21.0进行统计学分析,以上实验均重复3次,取均值,计量资料以表示,两组采用t检验分析相对荧光素酶活性,多组采用单因素方差分析HULC的表达量、细胞活力、凋亡率,以及侵袭和迁移细胞数,多组之间两两比较采用LSD-t检验的方法,P<0.05表示差异有统计学意义。
2 结果
2.1 HULC在宫颈癌细胞中呈高表达qRT-PCR检测结果显示,与Ect1/E6E7细胞1.00±0.11相比,C-33A、Caski、SiHa、HeLa细 胞 中HULC的 表 达 量(3.49±0.40、2.26±0.21、1.77±0.19、1.95±0.24)明显升高(F=39.92,P<0.001),在各宫颈癌细胞系中,C-33A细胞中HULC的表达量最高(P<0.05),因此选取C-33A细胞进行后续实验。
2.2 抑制HULC的表达对C-33A细胞增殖和凋亡的影响qRT-PCR检测转染HULC siRNA 48 h后C-33A细胞中HULC的表达量,结果显示,与Control组和NC组比较,si-HULC组细胞中HULC的表达量明显下调(P<0.05);Control组和NC组间相比,HULC的表达量差异无统计学意义(P>0.05)。CCK-8实验检测细胞增殖能力,与Control组和NC组比较,si-HULC组细胞活力显著降低(P<0.05);流式细胞术检测结果显示,与Control组和NC组比较,si-HULC组细胞凋亡率明显升高(P<0.05);Control组和NC组间相比,细胞OD值差异无统计学意义(P>0.05)。见表1,图1。
表1 各组C-33A细胞中HULC的表达量、细胞活力和凋亡率比较/

表1 各组C-33A细胞中HULC的表达量、细胞活力和凋亡率比较/
注:HULC为肝癌高表达转录本,OD为光密度。①与Control组比,P<0.05。②与NC组比,P<0.05。
组别Control NC si-HULC F值P值重复次数333 HULC 1.01±0.10 0.94±0.08 0.18±0.02①②113.52<0.001 OD值0.64±0.11 0.59±0.08 0.38±0.05①②8.16 0.019凋亡率/%2.88±0.30 3.01±0.34 18.54±2.16①②147.79<0.001
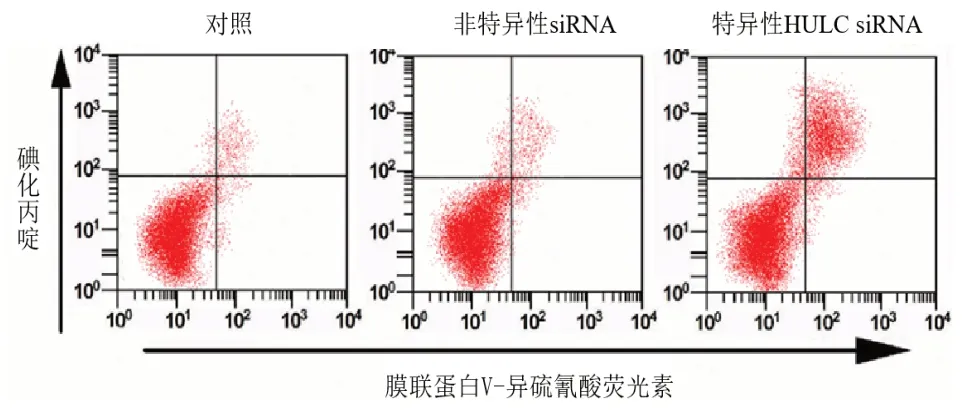
图1 流式细胞术检测细胞凋亡
2.3 抑制HULC的表达对C-33A细胞侵袭和迁移的影响Transwell实验结果显示,见图2,表2。si-HULC组侵袭细胞数和迁移细胞数显著少于Control组和NC组(P<0.05)。
表2 Transwell实验检测细胞侵袭和迁移能力/
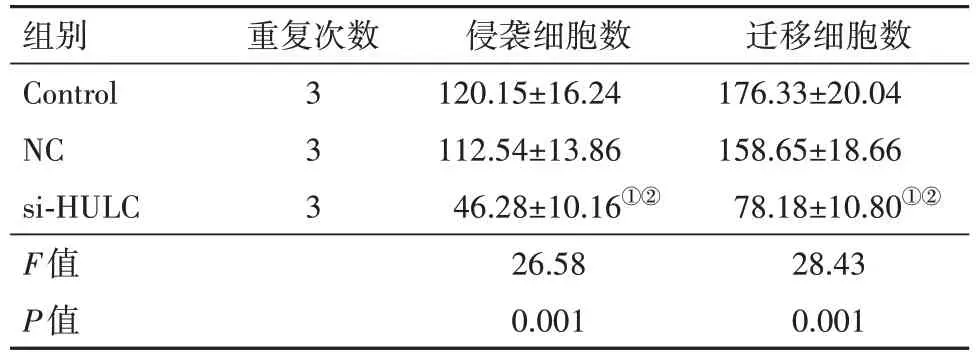
表2 Transwell实验检测细胞侵袭和迁移能力/
注:Control为对照,NC为非特异性siRNA,si-HULC为特异性HULC siRNA。①与Control组比,P<0.05。②与NC组比,P<0.05。
组别Control NC si-HULC F值P值重复次数333侵袭细胞数120.15±16.24 112.54±13.86 46.28±10.16①②26.58 0.001迁移细胞数176.33±20.04 158.65±18.66 78.18±10.80①②28.43 0.001
2.4 HULC抑制miR-134-5p的表达在线软件LncBase Experimental v.2预测 结果 显示,HULC和miR-134-5p有靶向结合位点。双荧光素酶报告基因实验结果显示,在HULC-Wt组中,转染miR-134-5p mimics的细胞相对荧光度酶活性明显低于miRNC组(P<0.05);在HULC-Mut组中,两组细胞相对荧光度酶活性无明显改变(P<0.05)。为进一步验证HULC对miR-134-5p表达的负向调控作用,本研究qRT-PCR检测抑制HULC的表达后对miR-134-5p表达的影响,结果显示,与NC组(1.00±0.09)相比,si-HULC组细胞中miR-134-5p表达(2.45±0.24)显著升高(t=16.37,P<0.05)。见图3,表3。
表3 双荧光素酶报告基因实验检测HULC和miR-134-5p的相互作用/

表3 双荧光素酶报告基因实验检测HULC和miR-134-5p的相互作用/
注:miR-NC为模拟物对照,miR-134-5p为miR-134-5p模拟物,HULC-Wt为HULC野生型载体,HULC-Mut为HULC突变型载体。
组别miR-NC miR-134-5p t值P值重复次数。33 HULC-Wt 1.00±0.10 0.23±0.02 22.65<0.001 HULC-Mut 1.00±0.09 0.99±0.09 0.24 0.817

图3 在线软件LncBase Experimental v.2预测HULC和miR-134-5p结合位点
2.5 抑制miR-134-5p的表达对C-33A细胞增殖、凋亡、侵袭和迁移的影响qRT-PCR检测结果显示,与si-HULC组 和si-HULC+anti-NC组 比,si-HULC+anti-miR-134-5p组 细 胞 中miR-134-5p的 表达水平明显降低(P<0.05)。CCK-8实验结果显示,与si-HULC组和si-HULC+anti-NC组比,si-HULC+anti-miR-134-5p组细胞OD值显著升高(P<0.05)。流式细胞术检测结果显示,与si-HULC组和si-HULC+anti-NC组比,si-HULC+anti-miR-134-5p组细胞凋亡率明显降低(P<0.05)。Transwell实验结果显示,与si-HULC组 和si-HULC+anti-NC组 比,si-HULC+anti-miR-134-5p组侵袭细胞数和迁移细胞数均明显增多(P<0.05)。si-HULC组和si-HULC+anti-NC组相比,以上各指标均差异无统计学意义(P>0.05)。见图4,5;表4。
表4 抑制miR-134-5p对C-33A细胞增殖、侵袭和迁移的影响/

表4 抑制miR-134-5p对C-33A细胞增殖、侵袭和迁移的影响/
注:si-HULC为特异性HULC siRNA,si-HULC+anti-NC为特异性HULC siRNA+抑制剂对照,si-HULC+anti-miR-134-5p为特异性HULC siRNA+miR-134-5p抑制剂,OD为光密度。①与si-HULC组比,P<0.05。②与si-HULC+anti-NC组比,P<0.05。
组别si-HULC si-HULC+anti-NC si-HULC+anti-miR-134-5p F值P值重复次数333 miR-134-5p 1.00±0.10 0.99±0.11 0.18±0.02①②88.57<0.001 OD值0.39±0.05 0.41±0.04 0.61±0.06①②17.30 0.003凋亡率18.14±2.25 19.22±2.43 8.27±1.14①②26.72 0.001侵袭细胞数48.02±10.04 47.96±8.64 96.28±12.12①②21.70 0.002迁移细胞数80.20±9.24 82.36±10.58 156.82±16.15①②37.39<0.001

图4 抑制miR-134-5p的表达对C-33A细胞凋亡的影响
3 讨论
宫颈癌是全球常见的妇科恶性肿瘤之一,也是全世界女性癌症死亡的主要原因[12]。探索宫颈癌发生的分子机制有利于开发宫颈癌的新疗法。本研究首次探讨了lncRNA HULC和miR-134-5p在宫颈癌中的表达模式和生物学功能。越来越多的证据表明,LncRNA可以在转录水平或转录后水平调节靶基因,参与机体多种生长发育和病理过程[13-14]。多项研究表明,lncRNA可作为致癌基因或抑癌基因参与肿瘤发生、浸润、转移、预后和诊断[15-16]。在先前的研究中,发现HULC表达上调参与调控肝癌、胃癌、前列腺癌等肿瘤细胞增殖、凋亡、侵袭和迁移[17-19]。以往研究结果表明,HULC可能在肿瘤细胞生物学行为中发挥重要功能。近期发现HULC在宫颈癌中表达亦呈高表达,且与病人预后不良密切相关[10]。但关于HULC与宫颈癌细胞生物学功能的研究鲜有报道,因此,本研究旨在探究HULC对宫颈癌细胞生物学行为的影响及分子机制。本研究结果发现,HULC在宫颈癌细胞中的表达显著高于人正常宫颈上皮细胞Ect1/E6E7。qRT-PCR检测结果显示,在宫颈癌C-33A细胞中转染HULC siRNA后HULC的表达显著下调,提示转染HULC siRNA可有效抑制HULC的表达。此外本研究发现,抑制HULC的表达后C-33A细胞OD值降低,凋亡率升高,侵袭和迁移细胞数减少,表明抑制HULC的表达可抑制宫颈癌C-33A细胞增殖、侵袭和迁移能力,诱导细胞凋亡。因此,推测HULC对宫颈癌的发生和发展具有重要作用。
lncRNA在人类疾病中的重要性可能与它们通过各种机制影响细胞功能的能力有关[20-21]。受到“竞争性内源RNA”调控网络的启发,假设HULC也可作为ceRNA,因此需寻找与HULC相互作用的潜在的miRNA。为支持这一观点,本研究采用生物信息学分析软件预测与HULC相互作用的miRNA,发现miR-134-5p与HULC可形成互补碱基配对。此外通过双荧光素酶报告基因验证了HULC与miR-134-5p相互作用。qRT-PCR检测结果进一步验证了HULC可负向调控miR-134-5p的表达。有研究发现,miR-134-5p可通过靶向整合素β1抑制非小细胞肺癌A549和H1299细胞的侵袭和迁移[22]。汤继英等[23]研究发现,miR-134-5p的过表达不仅可以抑制人宫颈癌细胞体外增殖和细胞周期进程,还可以诱导细胞凋亡。结合本研究结果提示,抑制HULC的表达抑制宫颈癌C-33A细胞增殖、凋亡、侵袭和迁移与上调miR-134-5p的表达有关。为进一步验证该结论,本研究在抑制HULC表达的C-33A细胞中转染miR-134-5p抑制物,检测发现,抑制miR-134-5p的表达后可逆转HULC表达下调导致的C-33A细胞增殖、侵袭和迁移的抑制作用,逆转HULC表达下调诱导的细胞凋亡,表明HULC靶向调控miR-134-5p参与宫颈癌细胞生物学行为的调控。
综上所述,本研究证明HULC在宫颈癌组织和细胞中表达上调,体外抑制宫颈癌C-33A细胞中HULC的表达可抑制细胞增殖、凋亡、侵袭和迁移能力,其作用机制可能与上调miR-134-5p的表达有关。关于miR-134-5p调控下游靶基因的研究本实验暂未涉及,后续研究将进一步探究。随着HULC的深入研究,其可能成为宫颈癌的潜在预后生物标志物和治疗靶点。
