3种陆生植物叶片浸提液对东海原甲藻生长及光合生理的影响
2022-09-22陈雯雯沈盎绿
向 舒,陈雯雯,沈盎绿
(上海海洋大学海洋生态与环境学院,上海 201306)
东海原甲藻(Prorocentrum donghaiense)赤潮在福建、浙江沿海常有发生,该藻已经成为了我国东海海域内引发赤潮次数最多、累计面积最大的肇事甲藻[1]。东海原甲藻赤潮期间造成水体缺氧,降低中华哲水蚤(Calanus sinicus)种群丰度从而影响渔业资源[2-3]。网箱养殖是海水增养殖生产的重要方式之一,赤潮的暴发会对网箱养殖产生严重影响[4]。因此,如何有效防治近岸水产养殖海域有毒有害赤潮成为目前我国赤潮灾害防治的热点和重点之一。
植物化感作用因简单易操作、对环境压力小且无二次污染、经济适用等优点,正逐渐成为当下藻华生物防治的重要手段之一。早期大多涉及陆生植物抑制淡水藻类,其中紫玉兰(Yulania liliiflora)、女贞子(Ligustri lucidi)、石榴(Punica granatum)和银杏(Ginkgo biloba)等陆生木本植物分泌的化感物质对铜绿微囊藻(Microcystis aeruginosa)和丝状藻(Cladophora sp.)等淡水藻类有较好的抑制效果[5-8]。但是在海洋赤潮藻类抑制方面,目前大多数研究聚焦在稻麦秆、玉米(Zea mays)和加拿大一枝黄花(Solidago canadensis)等 陆 生 草 本 植 物[9-10]。盐 角 草(Salicornia europaea)、互 花 米 草(Spartina alterniflora)[11-13]及红树林植物秋茄叶(Kandelia candel)、木榄(Bruguiera gymnorhiza)和海芒果(Cerbera manghas)等湿地植物[14-15]也具有化感抑藻作用。此外,少数研究发现,陆生木本植物对海洋藻类具有较好的抑制作用,比如托里桉(Eucalyptus tarelliana)粉和杉木(Cunninghamia lanceolata)粉能够有效去除水体中的米氏凯伦藻(Karenia mikiotoi)或东海原甲藻等海洋藻类[16-17]。常 见 木 本 植 物 香 樟(Cinnamomum camphora)、夹竹桃(Nerium indicum)和银杏虽被证明其落叶浸出液中化感物质可有效抑制黄丝藻(Tribonema bombycinum)、铜绿微囊藻等淡水藻类的生长,并导致其光合生理发生显著变化[8,18],但作为良好的化感物质候选者,这几种植物对赤潮生物东海原甲藻的抑藻效果却未见报道。
基于脉冲-振幅-调制的叶绿素荧光测量技术(pulse-amplitude-modulation,PAM),因快速且不伤害植物本身而被广泛用于测量植物光合系统Ⅱ(PSⅡ)的生理变化,可提供最大光化学量子效率(Fv/Fm)、有效光化学量子效率(Fv′/Fm′)、最大相对电子传递效率(rETRmax)和光能转化效率(α)等有效光合参数[19]。在植物化感抑藻方面的研究中,这些参数被广泛应用,研究发现荔枝(Litchi chinensis)、银杏和互花米草等植物中存在的化感物质对铜绿微囊藻、东海原甲藻和丝状藻等微藻的光合参数具有显著抑制作用[8,13]。
借鉴森林防火隔离带原理,当沿海养殖区面临赤潮威胁时,在网箱养殖区用陆生植物叶片粉末构筑一道赤潮隔离带,可实现保护海水养殖区的目的。本研究在前期筛选有效陆生植物的基础上,选择具有较好化感作用的香樟、夹竹桃和银杏叶片作为研究对象,探究其叶片浸提液对东海原甲藻的抑制效果;在此基础上,应用PAM测定3种植物叶片浸提液对东海原甲藻细胞的Fv′/Fm′、rETRmax和α等光合作用参数,从而综合评价香樟、夹竹桃和银杏这3种植物叶片浸提液对东海原甲藻生长及光系统PSⅡ的调控作用,为东海原甲藻生物防控提供依据。
1 材料与方法
1.1 实验材料
香樟、夹竹桃和银杏叶片于2019年10—11月收集自上海市上海海洋大学校园内。将清洗干净的材料置于60℃烘箱中烘干,粉碎过筛(100目),常温密封保持备用。
东海原甲藻(GY-H40)购自上海光语生物科技有限公司,藻类培养基采用f/2培养基。培养基所用海水采自浙江省舟山市漳州湾近岸(29°54′56″N、122°25′14″E),海水经过孔径为0.22μm的滤膜过滤,然后经过121℃高压蒸汽灭菌20分钟。藻类培养温度为(25±1)℃,光照强度大约为20μmol·m-2·s-1,光暗比为14 h∶10 h。藻类培养和实验均按照以上条件在新苗智能型光照培养箱(GZX-250BSH-Ⅲ,上海)进行。
1.2 实验方法
1.2.1 植物叶片浸提液的制备
称取50.0 g植物粉末,加入500 mL灭菌海水,60℃水浴锅中水溶2 h后取上清液,然后重新加入灭菌海水,多次重复上述操作至上清液无色。上清液再经0.22μm的滤膜过滤,除去颗粒杂质和微生物,得到最终实验浸提母液,质量浓度为50.0 g·L-1。
1.2.2 3种植物叶片浸提液抑藻实验
根据预实验结果,香樟和夹竹桃实验中每组实验设置1个对照组和4个质量浓度梯度为0.1、0.5、1.0和3.0 g·L-1的试验组。银杏实验中每组实验设置1个对照组和5个质量浓度梯度为0.1、0.5、1.0、3.0和5.0 g·L-1的试验组。将处于指数增长期的东海原甲藻接种于100 mL锥形瓶中,同时添加植物叶片浸提液,接种后的总体积为50 mL。香樟、夹竹桃和银杏植物叶片浸提液处理组中东海原甲藻的起始藻密度均为1.1×104个·mL-1。每个浓度3个重复组,培养条件与1.1相同。实验周期为96 h,每天光周期2 h后取样一次,进行藻细胞密度和叶绿素荧光参数的测定。
1.2.3 东海原甲藻叶绿素荧光参数的测定
通过浮游植物分类荧光仪(PHYTO-PAMED,Walz公司,德国)测定东海原甲藻的Fv′/Fm′和rETR,参数测定时光化学设置参见文献[20]。rETR和光照强度(I)之间关系按照EILERS和PEETERS[21]的公式进行拟合,Fv′/Fm′、rETRmax及α计算公式参见文献[20]。本实验光照强度设置为1、32、64、164、264、364、464、564、664、764 μmol photons·m-2·s-1共10个梯度,每个光强下照射时间为20 s。
1.3 数据分析
所有实验数据均以平均值±标准差(SD)表示,Shapiro-Wilk和Levene检验用于检验数据是否正态分布和方差齐性。在数据呈正态分布以及方差相等的前提下采用SPSS软件进行单因素方差分析,组间差异显著的情况下(P<0.05表示差异显著)采用Tukey’s方法进行多重比较。所有图表采用Origin 2015软件进行绘制。藻类抑制率计算方法参照文献[10]。96 h-EC50的计算采用GraphPad Prism 7.0软件[22]。
2 结果与分析
2.1 3种植物叶片浸提液对东海原甲藻生长的影响
由图1可知,0.1 g·L-1香樟浸提液处理96 h时间内,东海原甲藻藻细胞生长与对照组无显著差异(P>0.05);1.0 g·L-1和3.0 g·L-1浸提液在48 h对东海原甲藻的藻密度具有显著的抑制作用(P<0.05),96 h时藻密度的抑制率分别为57.6%和100.0%。鉴于对东海原甲藻的抑制率已达100%,所以更高浓度(>3.0 g·L-1)试验组在图1中不再显示。经计算,香樟浸提液对东海原甲藻的96 h-EC50值为0.84 g·L-1。
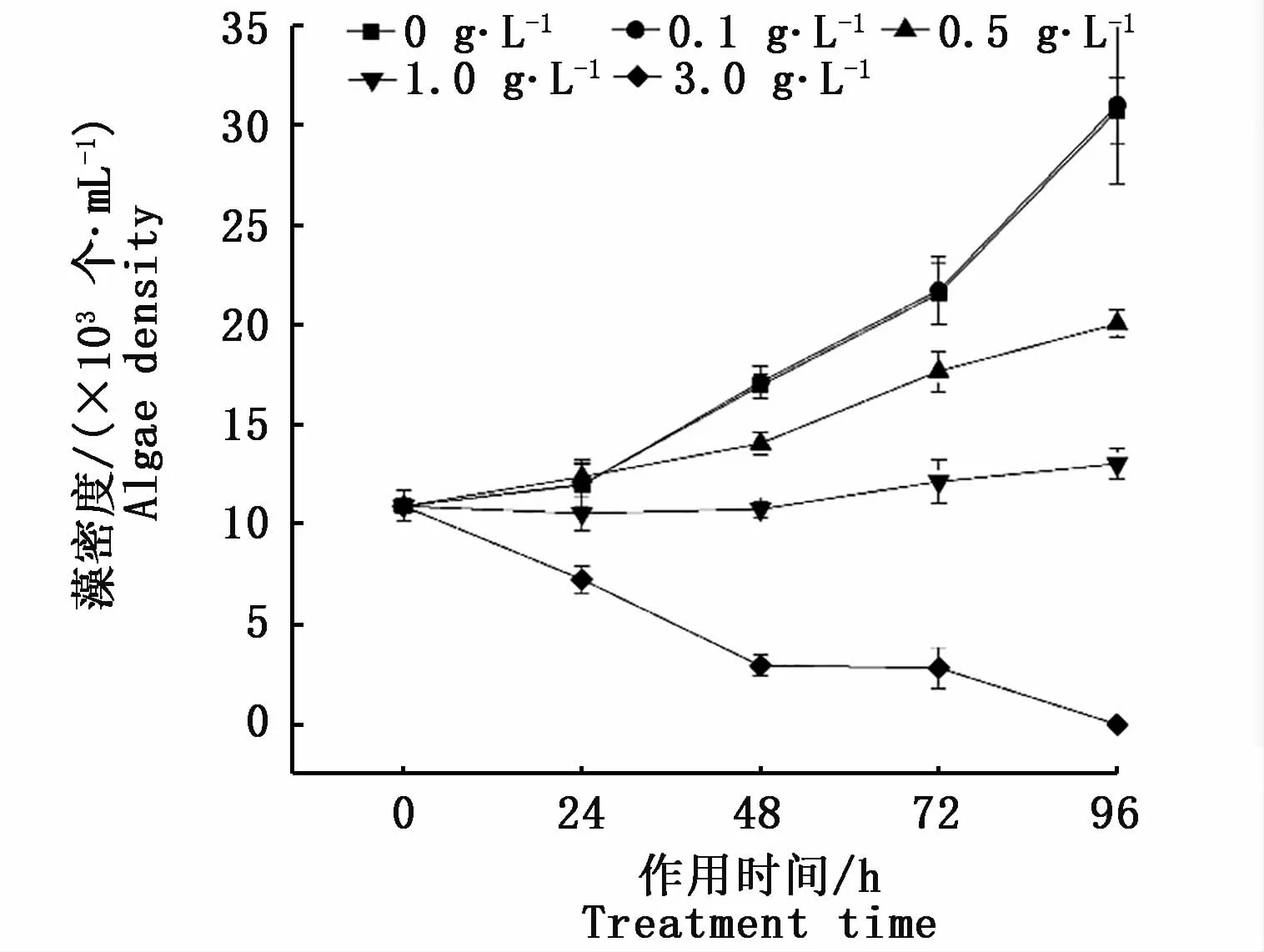
图1 不同梯度浓度香樟叶片浸提液对东海原甲藻藻密度的影响Fig.1 Effects of Cinnamomum camphora leaf extract concentration gradient on algae density of Prorocentrum donghaiense
由图2可知,夹竹桃浸提液对东海原甲藻的抑制效果趋势基本一致,0.1 g·L-1浸提液处理后东海原甲藻藻细胞生长与对照组无显著差异(P>0.05);1.0 g·L-1夹竹桃浸提液处理96 h后东海原甲藻密度与对照组相比下降较为明显,抑制率为38.1%(P<0.05)。高浓度3.0 g·L-1浸提液作用下,藻密度在48 h时出现急剧下跌,96 h藻密度抑制率为100%(P<0.05)。经计算,夹竹桃浸提液对东海原甲藻的96 h-EC50值为1.30 g·L-1。
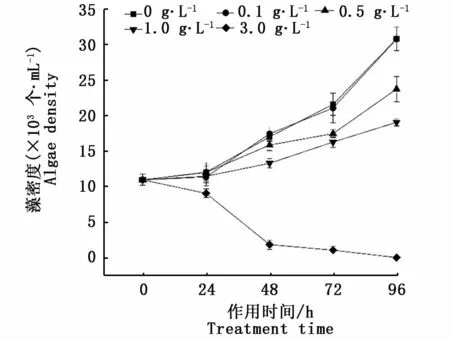
图2 不同梯度浓度夹竹桃叶片浸提液对东海原甲藻藻密度的影响Fig.2 Effects of Nerium indicum leaf extract concentration gradient on algae density of P.donghaiense
由图3可知,0.1 g·L-1银杏浸提液处理下东海原甲藻藻密度与对照组差异不显著(P>0.05),0.5 g·L-1和1.0 g·L-1浸提液处理72 h时藻细胞还能轻微生长。3.0 g·L-1和5.0 g·L-1浸提液处理后东海原甲藻的藻密度48 h时出现显著下降,96 h时抑制率达到74.9%和88.7%(P<0.05)。经计算,银杏浸提液对东海原甲藻的96 h-EC50值为1.74 g·L-1。

图3 不同梯度浓度银杏叶片浸提液对东海原甲藻藻密度的影响Fig.3 Effects of Ginkgo biloba leaf extract concentration gradient on algae densityof P.donghaiense
2.2 3种植物叶片浸提液对东海原甲藻PSⅡ的影响
2.2.1 对东海原甲藻F v′/F m′的影响
如图4-A所示,0.1 g·L-1香樟叶片浸提液处理96 h后东海原甲藻Fv′/Fm′值与对照组相比差异不显著(P>0.05)。而0.5 g·L-1和1.0 g·L-1叶片浸提液处理96 h后藻类Fv′/Fm′抑制率分别为19.4%和29.9%(P<0.05)。3.0 g·L-1浸提液处理下藻细胞的Fv′/Fm′值急剧下降,72 h时藻类Fv′/Fm′抑制率为100.0%(P<0.05)。如图4-B所示,0.1 g·L-1夹竹桃叶片浸提液处理96 h内藻细胞Fv′/Fm′值与对照组无显著差异(P>0.05)。0.5 g·L-1和1.0 g·L-1叶片浸提液处理后藻细胞Fv′/Fm′值出现小幅度下降,96 h后藻类Fv′/Fm′抑制率分别为11.9%和19.4%(P<0.05)。3.0 g·L-1浸提液处理下藻细胞的Fv′/Fm′值剧烈下降,96 h后藻类Fv′/Fm′抑制率为97.0%(P<0.05)。由图4-C所示,浓度低于1.0 g·L-1银杏叶片浸提液处理时,藻细胞Fv′/Fm′值与对照组无显著差异(P>0.05)。3.0 g·L-1叶片浸提液处理96 h后藻类Fv′/Fm′抑制率为34.9%,5.0 g·L-1叶片浸提液作用24 h后藻类Fv′/Fm′抑制率为50.8%,96 h后藻类Fv′/Fm′抑制率达88.9%(P<0.05)。

图4 不同梯度浓度香樟(A)、夹竹桃(B)、银杏(C)叶片浸提液对东海原甲藻有效光化学量子效率(F v′/F m′)值的影响Fig.4 Effects of C.camphora(A),N.indicum(B)and G.biloba(C)leaf extract concentration gradient on effective photochemical efficiency(F v′/F m′)of P.donghaiense
2.2.2 对东海原甲藻PSⅡ光合活性参数的影响
通过测量藻细胞的快速光曲线得到香樟叶片浸提液对东海原甲藻PSⅡ光合活性参数(图5)。0.1、0.5 g·L-1和1.0 g·L-1香樟叶片浸提液处理72 h内,东海原甲藻的α值与对照组差异不显著(P>0.05),浸提液处理96 h后,0.1 g·L-1和0.5 g·L-1组虽然显著低于对照组和1.0 g·L-1组,但是绝对值相差不大。藻细胞rETRmax值和Ik值在实验期间的变化幅度则大于α值,处理组浓度越高,两者的数值越低(P<0.05)。高浓度3.0 g·L-1浸提液处理下,藻细胞α值、rETRmax值和Ik值从24 h开始就显著低于对照组(P<0.05),并且在72 h时表现出急剧降低,96 h后藻类的PSⅡ光合活性参数均处于极低状态(P<0.05)。

图5 不同梯度浓度香樟叶片浸提液对东海原甲藻光能转化效率α(A)、最大相对电子效率r ETRmax(B)及光饱和系数I k(C)的影响Fig.5 Effects of C.camphora leaf extract concentration gradient on the photosynthesis efficiencyα(A),relative maximal electron transport rates r ETRmax(B)and light saturation coefficient I k(C)of P.donghaiense
如图6所示,夹竹桃叶片浸提液在低浓度范围内(0.1、0.5 g·L-1和1.0 g·L-1)对东海原甲藻PSⅡ光合活性参数的影响与香樟叶片浸提液对其的影响趋势基本一致,东海原甲藻α值差异不显著(P>0.05)。3.0 g·L-1处理组对东海原甲藻α值、rETRmax值和Ik值都在24 h出现骤降,但在96 h后却有所回升,各个参数的藻类抑制率分别为57.4%、94.0%和70.1%(P<0.05),均未达到完全抑制。

图6 不同梯度浓度夹竹桃叶片浸提液对东海原甲藻光能转化效率α(A)、最大相对电子效率r ETRmax(B)及光饱和系I k(C)的影响Fig.6 Effects of N.indicum leaf extract concentration gradient on the photosynthesis efficiencyα(A),relative maximal electron transport rates r ETRmax(B)and light saturation coefficient I k(C)of P.donghaiense
如图7所示,0.1、0.5 g·L-1和1.0 g·L-1银杏叶片浸提液处理下,东海原甲藻的α值、rETRmax值和Ik值在72 h内都与对照组无显著差异(P>0.05),浸提液处理96 h后0.1、0.5和1.0 g·L-1组虽然与对照组有差异,但是绝对值也相差不大。3.0 g·L-1浸提液作用下,96 h内藻细胞α值出现小幅度下降(P<0.05),rETRmax值和Ik值与对照组相比有小幅波动(P>0.05)。5.0 g·L-1浸提液处理后,东海原甲藻α值、rETRmax值和Ik值在实验期间不断下降,96 h后各个参数的藻类抑制率分别达到88.9%、96.1%和62.2%(P<0.05)。
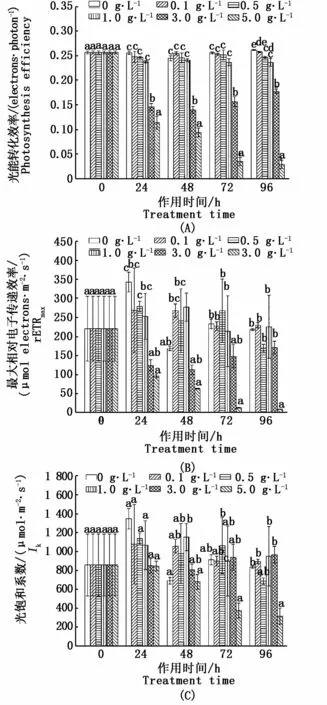
图7 不同梯度浓度银杏叶片浸提液对东海原甲藻光能转化效率α(A)、最大相对电子效率r ETRmax(B)及光饱和系I k(C)的影响Fig.7 Effects of G.biloba leaf extract concentration gradient on the photosynthesis efficiencyα(A),relative maximal electron transport rates r ETRmax(B)and light saturation coefficient I k(C)of P.donghaiense
3 讨论
3.1 不同植物提取物对藻类抑制效果的影响
近年来赤潮发生的频率和危害不断上升,自2000年以来东海海域经常暴发大规模东海原甲藻赤潮[23],所以筛选有效的生物防治方法控制东海原甲藻赤潮成为十分迫切的事情。由于陆生植物种类多、来源广、化感物质更丰富,所以在抑藻剂原材料选择上,陆生植物提供了更多选择[24]。香樟、夹竹桃及银杏是我国常见的绿化树种,其树形高大、枝叶丰满,利用其落叶化感作用抑制藻类生长,不仅可以有效防控赤潮的发生,也能实现落叶的废物利用。
在本研究中,东海原甲藻在对照组中一直处于指数增长期,但在3种植物浸提液处理组中藻类生长受到不同程度的抑制,发现香樟的抑制效果最为显著,其次是夹竹桃,银杏效果最差。香樟[25]、夹竹桃[26]及银杏[27]水提液均含有丰富的黄酮类化合物,这类化合物对藻类生长具有抑制作用。3种植物浸提液抑藻程度差异化的主要原因可能是黄酮化合物种类不同,不同种类黄酮类化合物中羟基个数不同,羟基个数越多抑藻效果越好;其次黄酮分子结构中引入3位亲水基团、A环5位带负电氢键供体基团、B环4’位亲水氢键供体基团及3’、4’位带负电基团有利于化合物抑藻效果的增加,反之,A环6、7位引入带负电羟基供体基团或B环2’、3’、5’及6’位引入氢键供体基团能削弱黄酮化合物的抑藻效果;此外黄酮类化合物分子极性越大、通透性越高及疏水性越强,其抑藻活性也更高[28]。除黄酮类化合物之外,木脂素类化合物是樟属植物中的一类主要化学成分,这类物质被降解产生氧化类多酚可能抑制藻类生长[29]。夹竹桃叶片中特有的夹竹桃苷类[26]、银杏叶片中大量的有机酸类[27]等化合物也是导致抑藻活性差异的重要原因。这3种植物有效物质成分的组成和含量存在差异,后续将通过液相色谱质谱联用仪等仪器定量分析3种植物叶片水提液的各种有效成分的组成和含量,为后期分析相关抑藻作用机制提供更为精确的靶点。
3.2 植物化感作用对藻类光合作用的影响
叶绿素荧光测量技术是测量植物光合作用最经典的手段,因快速、灵敏、无损伤而被用于赤潮藻类光合机制的研究,利用叶绿素荧光分析技术可以有效监控藻细胞PSⅡ的生理状态,光合系统PSⅡ的Fv′/Fm′值、α值和rETRmax值取决于电子传输链的容量或Calvin循环的限制[30-31]。在研究植物化感物质对藻类作用机理中,PSⅡ是被认为最敏感的位点之一,特别是对电子传递链某一区段表现出强烈的抑制作用。本研究中香樟、夹竹桃和银杏植物叶片中都含有丰富的黄酮类物质,3种植物浸提液高浓度试验组会导致东海原甲藻Fv′/Fm′值、α值和rETRmax值显著降低。相似地,黄皓旻[28]发现,黄酮类物质木犀草素会导致铜绿微囊藻的Fv/Fm值、α值和rETRmax值显著下降,破坏铜绿微囊藻的光合作用从而抑制其生长。李超等[32]发现,黄酮类物质槲皮素能胁迫球形棕囊藻(Phaeocystis globsa)的光合系统,导致球形棕囊藻的Fv/Fm、Fv′/Fm′值、α值和rETRmax值呈不同程度降低,对球形棕囊藻光合作用影响明显。
本研究选取的3种陆生植物香樟、夹竹桃和银杏叶片中都含有丰富的黄酮类物质[25-27],因此,这3种植物浸提液中化感物质可能是通过阻断东海原甲藻第一电子受体QA和第二电子受体QB之间电子流,中断藻细胞PSⅡ中电子传递链,从而抑制东原甲藻的光合作用。化感物质往往是抑制藻细胞光合作用的关键。与此观点类似,有学者认为黄酮类化合物,特别是木犀草素,通过破坏光合系统中第二电子受体QB功能和降低有效量子产量,阻断光系统中PSⅡ之间电子流,导致光合作用下降[33]。LEU等[34]发现,穗花狐尾藻(Myriophyllum spicatum)中的特里马素Ⅱ会影响淡水蓝藻及菠菜的光合作用,3μmol·L-1纯特里马素Ⅱ通过阻断光系统中第一电子受体QA和第二电子受体QB之间电子流来抑制其光合作用。HERNÁNDEZ-TERRONSE等[35]发现,从菊科植物中分离出trachyloban-19-oic acid会阻断ATP的合成和PSⅡ中H2O→2,6-二氯靛酚(DCPIP),其抑制位点也可能是QA和QB之间的电子传递链。ZHU等[36]认为,穗花狐尾藻中的化感物质焦性没食子酸和没食子酸能抑制铜绿微囊藻和羊角月牙藻(Selenastrum capricornutum)的光合系统活性和电子传递链,其抑制位点在PSⅡ放氧复合体到QB间。
4 小结
香樟、夹竹桃和银杏3种植物叶片浸提液对东海原甲藻生长具有不同程度的抑制作用,综合分析东海原甲藻藻密度及96 h-EC50发现,香樟的抑制效果最佳,夹竹桃次之,这种抑藻效率的差异可能是3种植物中所含的化感物质种类和含量不同。香樟、银杏和银杏叶片高浓度浸提液(≥3.0 g·L-1)可以大幅度降低东海原甲藻Fv′/Fm′值和藻细胞光合参数α、rETRmax和Ik值,对东海原甲藻的光合作用有显著的抑制作用。因此,这3种植物浸提液中化感物质很有可能通过阻断东海原甲藻第一电子受体QA和第二电子受体QB之间电子流,中断藻细胞PSⅡ中电子传递链,从而抑制东海原甲藻的光合作用。
