鸡皮肤毛囊生长差异基因的生物信息学分析
2022-09-22刘祎轩朱慧媛张朋威雷艳茹高超群李东华康相涛
张 豪,刘祎轩,朱慧媛,张朋威,雷艳茹,高超群,李东华,康相涛
(河南农业大学动物科技学院,农业农村部畜禽资源(家禽)评价利用重点实验室,郑州 450000)
在现代商业生产中,家禽羽毛的数量和质量越来越受到消费者的关注,羽毛生长速度、质量和脱毛方式对高价值家禽胴体的生产具有重要意义。在家禽规模化生产中,15%的鸡存在不同程度的羽毛发育问题,如脱毛和身体覆盖不足等[1-2],这些问题严重影响了鸡的能量代谢、活体外观和胴体质量。羽毛生长不良不仅影响活体外观,而且降低胴体质量和饲料转化效率[3-4]。羽毛是一种起源于外胚层表皮细胞的角质化衍生物[5],其生长发育的调控机制目前仍不清楚。羽毛发育始于皮肤毛囊,它是一个由上皮(表皮)和间充质(真皮)成分组成的复杂微器官,通过上皮和间充质之间的关键分子相互作用调控羽毛的生长、替换及其形态结构的变化[6-7]。在羽毛形态发生前真皮细胞的分化过程并没有被预先决定,胚胎皮肤间充质通过与发育中的表皮相互作用,使表皮基板下的间充质细胞采用出芽或芽间分化的方式形成真皮凝聚物,凝聚物在随后的形态发生过程中再进一步形成毛乳头[8-9]。在胚胎发育早期,皮肤上皮和间充质可以通过细胞的紧密排列和一系列基因表达的变化来相互作用形成羽毛原基,羽毛原基进一步分化,开始在羽毛芽的不同部位表达不同的信号、生长和细胞黏附分子,然后在毛囊内长成细丝,通过孵化产生羽绒[9]。因此,探究家禽羽毛形态发生发育过程中上皮和间充质的相互作用,了解家禽羽毛生长发育的分子机制,对于促进羽毛生长进而提高家禽养殖业的经济效益具有重要意义。
在鸡胚胎期前6 d,鸡胚皮肤发育不全,表皮层非常薄,很难区分皮肤的表皮和间充质结构[10]。在鸡胚胎期第7天,胚胎背部从肱到腰骶出现羽芽,腰骶水平处有约7排羽芽,腿部出现明显的羽芽,而在尾巴的每个侧边上羽芽较模糊。在鸡胚胎期第9天时,羽芽更加明显,新出现的羽芽靠近中腹线,与胸骨相连,并延伸至腹部两侧[11]。在鸡胚胎发育的第7~9天是鸡皮肤上皮和间充质调控羽毛毛囊发育的关键时期。鸡胚胎期第7天首次产生的背部羽毛在形态上被鉴定,第7~9天突出的毛囊建立了良好毛细血管供应,并渗透到初期毛囊的毛乳头中[12],血管系统的建立对毛囊的后期生长至关重要。本研究通过下载GSE62882芯片表达数据集,经生物信息学分析筛选鸡胚胎期第7~9天影响羽毛生长发育的关键基因及其参与的生物过程和信号通路,为进一步探究皮肤上皮和间充质在毛囊生长发育中的作用提供理论参考。
1 材料与方法
1.1 基因芯片数据分析
GSE62882芯片表达数据集以Affymetrix基因组阵列GPL3213平台为基础,从GEO数据库(https:∥www.ncbi.nlm.nih.gov/geo/)中下载获得,本研究中共有8个Cobb500肉鸡样本数据,其中胚胎期第7天背部皮肤上皮(E7 dorsal skin epithelium,E7ep)、胚胎期第9天背部皮肤上皮(E9 dorsal skin epithelium,E9ep)、胚胎期第7天背部皮肤间充质(E7 dorsal skin mesenchyme,E7me)和胚胎期第9天背部皮肤间充质(E9 dorsal skin mesenchyme,E9me)样本数据各2个(表1)。将原始表达矩阵通过R语言中的limma包进行标准化分析,以log2|FoldChange|>1且P<0.05为筛选条件,筛选出E7ep和E9ep、E7me和E9me的差异基因,并绘制火山图及热图。
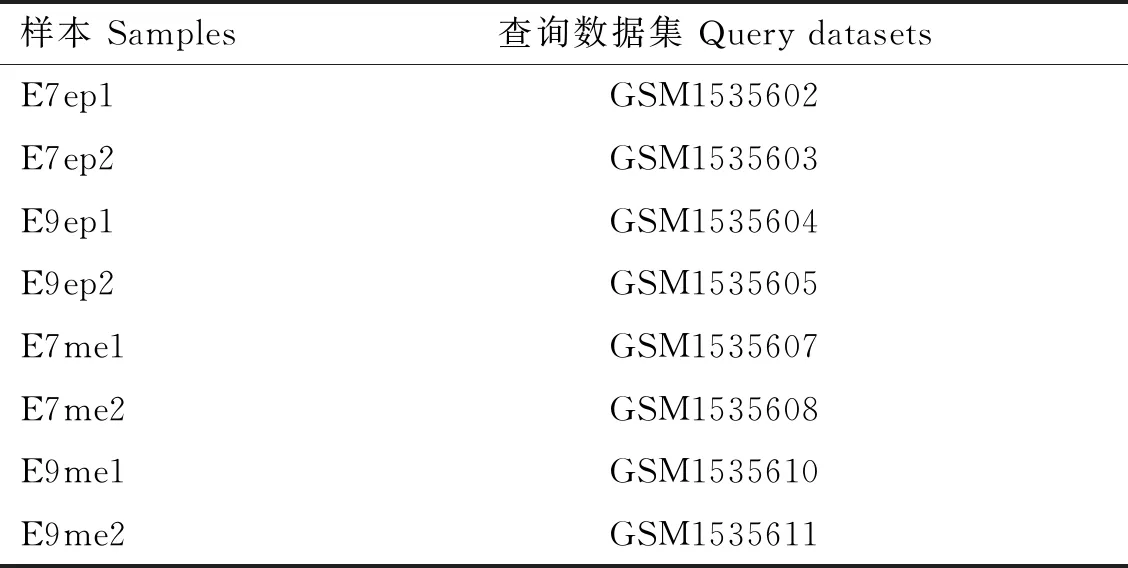
表1 GSE62882样本数据
1.2 关键基因筛选
将E7ep和E9ep、E7me和E9me差异基因,以及E7ep/E9ep、E7me/E9me差异基因中共同上下调基因分别导入STRING version 11.5在线软件(https:∥string-db.org/),得到蛋白互作(protein-protein interaction,PPI)网络,同时将PPI网络导入Cytoscape 3.8.2软件进行可视化。运用STRINGAPP插件计算PPI网络中各节点的拓扑参数,基于Degree选择PPI网络排名前十的关键基因,找出网络中处于中心位置的关键节点,并构建PPI子网络图。
1.3 差异基因GO功能注释
GO功能富集分析是大规模基因功能注释的常用方法,本研究取E7ep和E9ep、E7me和E9me的差异基因运用R语言中clusterProfiler程序包进行功能注释,获取差异基因的GO条目。
1.4 差异基因的信号KEGG通路富集分析
KEGG是一个包含生物化学路径的生物信息学数据库。本研究将E7ep和E9ep、E7me和E9me的差异基因,以及表皮与间充质中共同上下调基因,利用R语言中clusterProfiler程序包进行差异基因KEGG通路富集分析,获取差异基因和共同上下调基因富集的信号通路。
2 结 果
2.1 E7ep和E9ep差异基因的筛选和PPI网络的构建
本研究共筛选出E7ep和E9ep差异基因626个,其中上调基因356个,下调基因270个(图1)。选取前25个差异基因绘制热图,发现E7ep和E9ep差异基因包括SPINK5、LRRC17、GPIHBP1等(图2);将差异基因导入STRING数据库进行PPI网络分析,经Cytoscape 3.8.2软件构建PPI网络的可视化结果,剔除孤立蛋白后,共得到119个节点及189条互作关系(图3),基于Degree选择PPI网络排名前十的核心基因分别为WNT3A、FN1、WNT7A、FZD8、WNT3、WIF1、SHH、WNT9B、WNT4、WNT6(图4)。
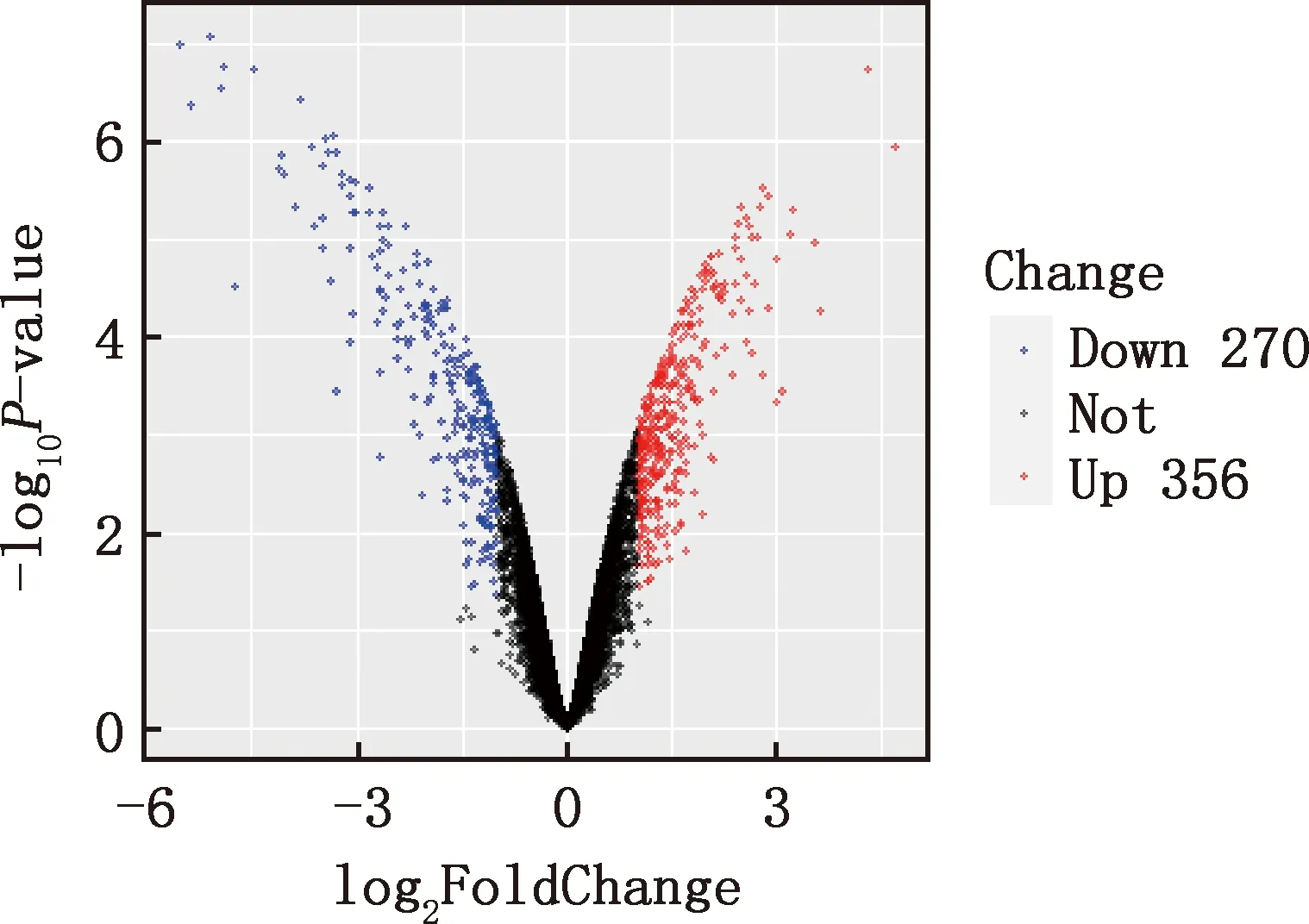
图1 E7ep和E9ep差异基因的火山图
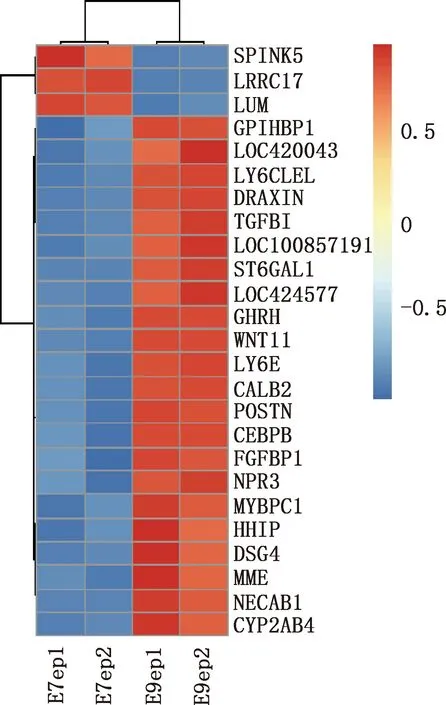
图2 E7ep和E9ep差异基因的热图
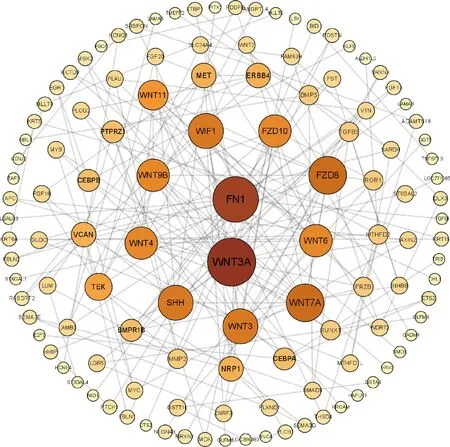
图3 E7ep和E9ep差异基因的PPI网络分析

图4 E7ep和E9ep差异基因的PPI网络枢纽模块分析
2.2 E7ep与E9ep差异基因的GO功能注释和KEGG通路富集分析
对E7ep和E9ep差异基因进行GO功能注释和KEGG通路富集分析,结果显示,差异基因参与细胞分化、解剖结构形态发生、神经系统发育、组织发育、神经形成、神经元的形成、神经分化、细胞黏着、生物黏附、多细胞有机体负调控等GO条目(图5),参与Wnt信号通路、黑色素合成、TGF-β信号通路、mTOR信号通路等信号通路(图6)。

图5 E7ep和E9ep差异基因的GO功能注释
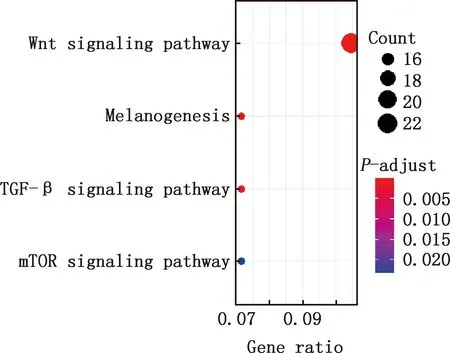
图6 E7ep和E9ep差异基因的KEGG通路富集分析
2.3 E7me和E9me差异基因筛选和蛋白互作网络构建
本研究共筛选出E7me和E9me差异基因690个,其中上调基因269个,下调基因421个(图7)。选取前25个差异基因制备热图(图8),发现E7me和E9me差异基因包括TAC1、FRK、PENK等;将差异基因导入STRING数据库网站进行PPI网络分析,经Cytoscape 3.8.2软件构建PPI网络的可视化结果,剔除孤立蛋白后,共得到206个节点及326条互作关系(图9),基于Degree选择PPI网络排名前十的核心基因分别为FGF2、SDC1、ITGB3、BMP4、EGFR、ITGA9、FLT1、MYL10、RUNX2、PTPN11(图10)。
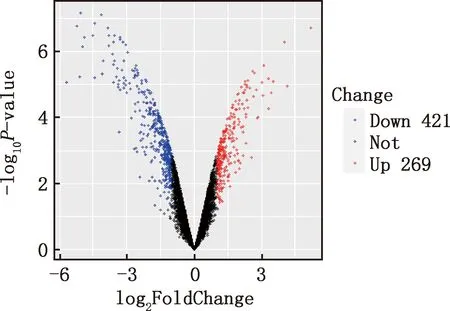
图7 E7me和E9me差异基因的火山图
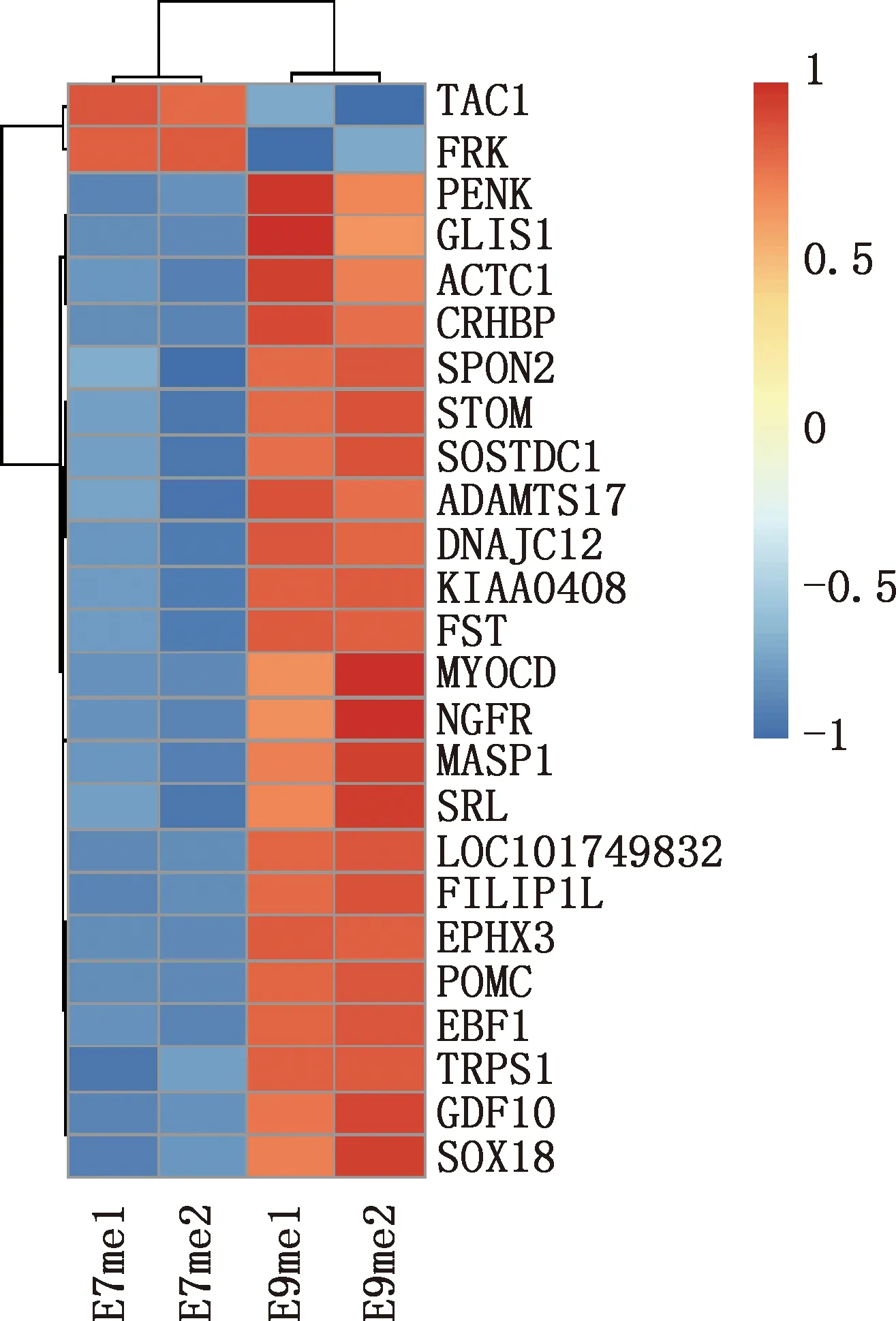
图8 E7me和E9me差异基因的热图

图9 E7me和E9me差异基因的PPI网络分析
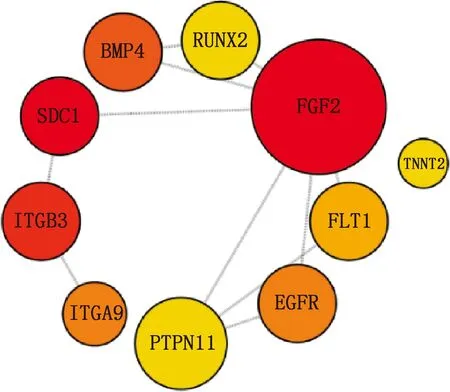
图10 E7me和E9me差异基因的PPI网络枢纽模块分析
2.4 E7me和E9me差异基因的GO功能注释和KEGG通路富集分析
对E7me和E9me差异基因进行GO功能注释和KEGG通路富集分析,结果显示,差异基因参与细胞分化、细胞发育过程、解剖结构的形态发生、动物器官发育、细胞表面受体信号通路、系统进程、循环系统的发展、解剖结构形态发生的调控、心脏发育、肌肉系统过程等GO条目(图11),参与钙信号通路、肌动蛋白骨架调控、EMC受体互作等信号通路(图12)。
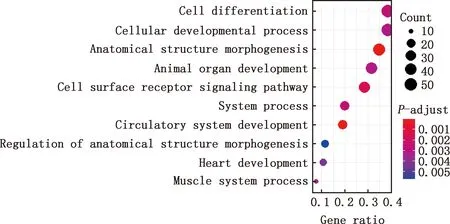
图11 E7me和E9me差异基因的GO功能注释
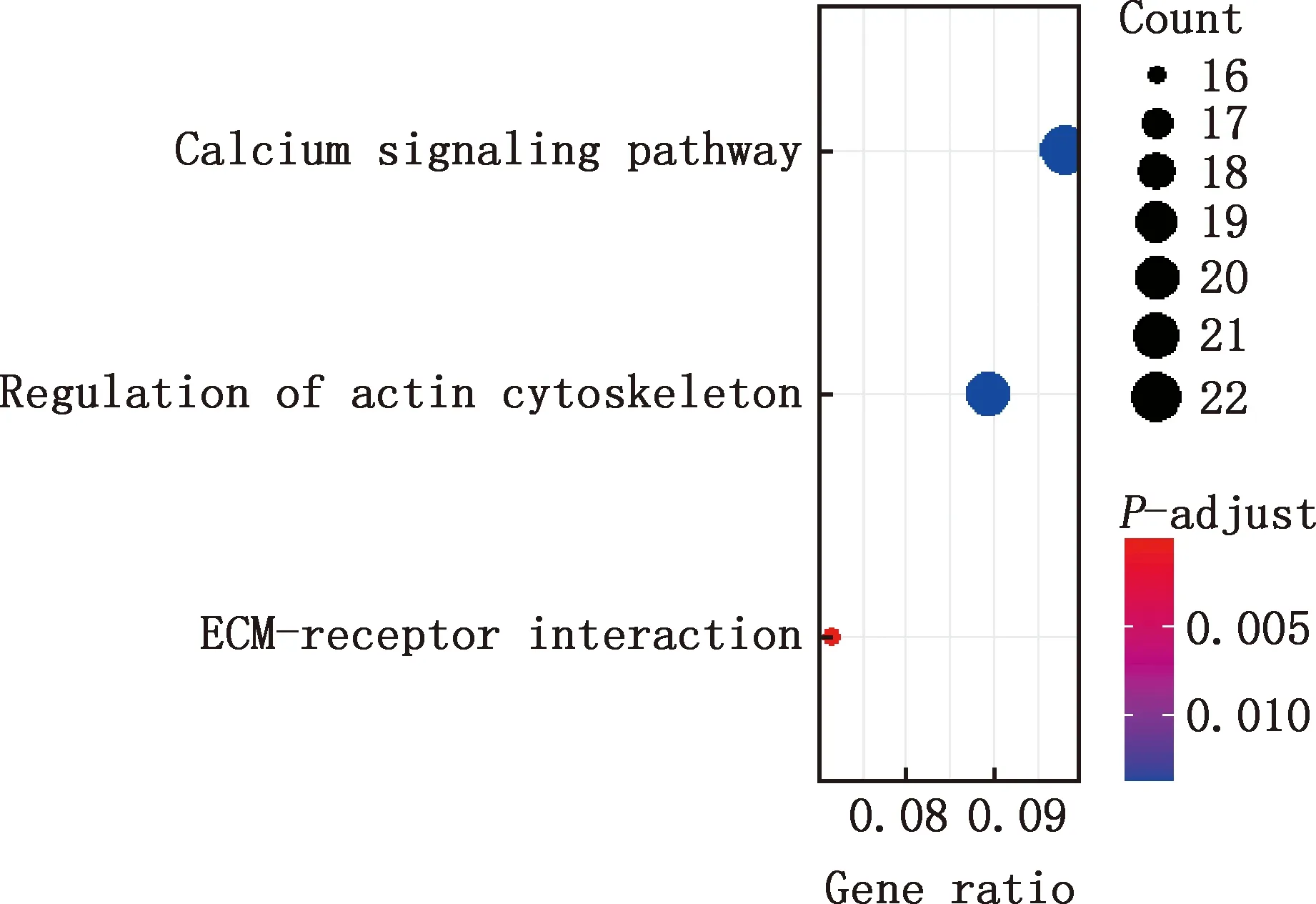
图12 E7me和E9me差异基因的KEGG通路富集分析
2.5 皮肤上皮和间充质中共同上下调基因的生物信息学分析
筛选鸡胚胎从第7和9天发育过程中背部皮肤上皮和间充质共同上下调基因,获得共同上调基因26个(图13A),共同下调基因48个(图13B)。将共同上下调基因导入STRING数据库进行PPI网络分析,通过Cytoscape 3.8.2软件构建PPI网络的可视化结果。剔除孤立蛋白后,PPI网络中共得到32个节点及34条相互作用关系,主要基因包括MYC、FGF18、RUNX2、JAG1、MSX2等(图14)。将共同上下调基因进行KEGG通路富集分析,结果发现共同上下调基因主要富集在Wnt信号通路上(图15)。
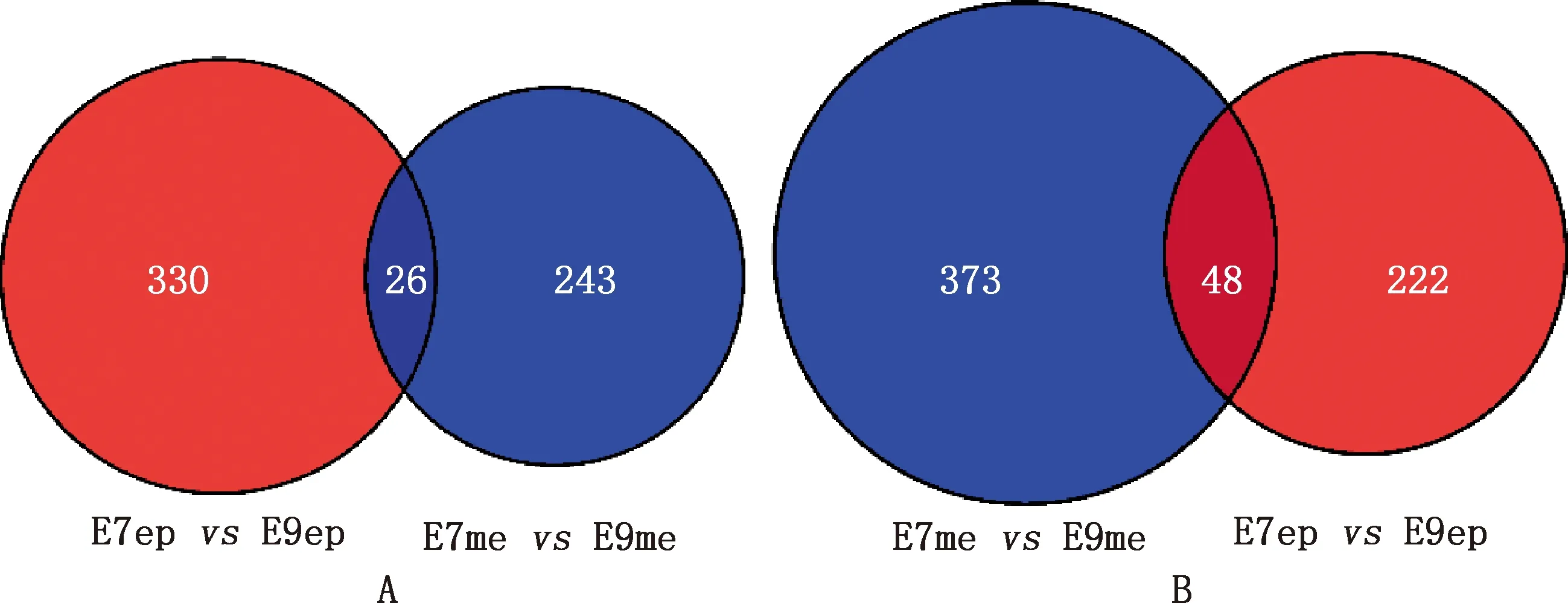
图13 皮肤上皮与间充质共同上调(A)和下调(B)基因

图14 共同上下调基因的PPI网络分析

图15 皮肤上皮与间充质共同上下调基因的KEGG通路富集分析
3 讨 论
羽毛是鸡的一个重要的外观性状,成鸡羽毛的发育呈现从生长期、退行期到休止期的周期性生长,羽毛的生长性状能够反映鸡的发育状况,羽毛的正常生长和再生对提高家禽的福利和经济价值具有重要意义[13-14]。羽毛是由皮肤上皮和间充质之间的一系列信号通路相互作用而形成的,上皮组织中的毛囊干细胞接收来自真皮间充质细胞的信号,并将信号传回真皮间充质细胞,从而影响皮肤附属物毛囊的生长发育[13]。目前关于鸡胚胎皮肤毛囊发育的研究已有大量报道,但对早期毛囊发育过程中上皮和间充质间相互作用调控毛囊的生长发育的研究较少,本研究分析GSE62882数据集中胚胎期第7、9天的背部皮肤上皮和间充质的差异表达基因,筛选鸡胚胎第7~9天发育过程中影响鸡羽毛生长发育的关键基因、生物过程和信号通路,探究皮肤上皮和间充质协同作用对毛囊生长发育的影响。
本研究共获得E7ep和E9ep差异基因626个,这些差异基因存在189条互作关系,基于Degree获得PPI网络排名前十的核心基因分别是WNT3A、FN1、WNT7A、FZD8、WNT3、WIF1、SHH、WNT9B、WNT4、WNT6,主要集中在Wnt信号通路上,通过GO功能注释和KEGG通路富集分析发现,E7ep和E9ep差异基因主要富集在血管系统发育相关的生物过程和Wnt、TGF-β信号通路上。在个体皮肤附属物中,WNT帮助形成局部生长区域,并形成发育中的羽毛,如WNT6过表达诱导不同羽毛区域的局部生长,WNT3A扩大了羽毛道的大小、促进了个体羽毛的形成[15-16]。TGF-β信号通路中最重要的成员是TGF-β超家族,该家族包含大量不同的多肽形态发生因子,包括TGF-β本身及BMP和GDF,能够诱导毛囊进入退行期,如在小鼠中研究发现,TGF-β2通过激活IFN-γ和BDNF诱导毛囊进入退行期[17]。在毛囊生长发育初期,毛囊干细胞的激活需要抑制TGF-β信号通路中BMP的表达和激活Wnt信号通路中β-catenin的稳定表达共同完成。Wnt、TGF-β信号通路对血管的生成具有重要的调控作用[15,18]。在鸡胚胎第7~9天的发育过程中,胚胎皮肤上皮开始有血管和神经的生成,毛囊周围血管可提供毛囊生长所需的营养物质及各类细胞调控因子,进而调控毛囊细胞的生长分化[19]。
本研究共获得E7me和E9me差异基因690个,基于Degree获得PPI网络排名前十的核心基因分别是FGF2、SDC1、ITGB3、BMP4、EGFR、ITGA9、FLT1、MYL10、RUNX2、PTPN11。活化的内皮细胞在FGF2的作用下开始增殖,并形成新的芽[18],使细胞间的接触松动,导致毛细血管融合形成动脉和静脉。FGF2可以通过诱导鸡皮肤形成致密的真皮组织,致密真皮组织进一步发育成羽芽[20]。在表皮发育过程中,BMP信号传导是皮肤上皮和间充质相互作用过程中的关键途径,具有抑制毛囊生长发育的作用[13,21]。研究发现,EGFR抑制剂能够缩短羽芽之间的距离并增加羽芽的数量[22];小鼠缺乏EGFR将不能长出毛发,小鼠皮肤中缺失EGFR将导致基质细胞有丝分裂活性增加、毛囊凋亡和形成毛发的上皮细胞系分化受损[23]。但EGFR在禽类皮肤毛囊生长发育过程中是否有相似的作用仍需进一步研究。E7me和E9me差异基因主要涉及肌肉发育、循环系统的发育和钙信号通路,钙信号通路能够影响皮肤间充质细胞的运动方向,在加速伤口愈合、减少疤痕形成、伤口收缩和诱导毛发再生等过程中有广泛的应用[24]。
本研究共获得胚胎期第7和9天鸡背部皮肤上皮和间充质中共同上调基因26个,共同下调基因48个,共同上下调基因PPI网络中的主要基因包括MYC、FGF18、JAG1等。MYC在人和小鼠毛囊周期中已有报道,参与小鼠毛囊形态发生的不同阶段及毛囊角化细胞的增殖或分化[25-26],c-Myc蛋白是β-catenin下游的转录因子,在鸡胚胎期第8~12天表达量显著增加,在调节毛囊干细胞分化中是必不可少的[27]。FGF18在小鼠毛囊中高表达,可诱导休止期毛囊再生,在鸡羽毛生长中的作用目前仍不清楚[28]。JAG1是Notch的配体,通过Notch信号通路参与毛囊的生长调控[29-31]。Notch信号通路被认为是调节包括毛囊在内的多细胞生物干细胞稳态和发育的主要信号转导通路。在毛囊发育过程中,Notch1的活性是决定细胞命运的关键。Notch1在早期基板的形成中并不发挥作用,但与真皮内陷毛乳头的形成有关。Notch1敲除以后将会导致退行期提前,毛发逐渐变短[32-33]。对背部皮肤上皮和间充质共同上下调基因进行KEGG通路富集分析,结果发现共同上下调基因主要富集在Wnt信号通路上。Wnt/β-catenin信号通路在胚胎期第8~12天能够促进羽毛的生长和发育[34-35]。Wnt信号通路在背部皮肤上皮和间充质互作中具有重要作用,在皮肤羽毛形成的诱导期,毛囊的形成依赖于单层上皮和真皮细胞浓缩物之间的一系列相互作用,上皮细胞接收来自底层间充质的指示信号,而间充质细胞需要表皮信号来形成毛乳头。在毛囊基板形成之前,真皮间充质细胞均匀地接受广泛的上皮发出的Wnt信号,真皮间充质细胞随后产生第一个真皮Wnt信号,诱导表皮基底细胞定期聚集,导致毛发基板的形成,初始诱导后,发展中的基板产生Wnt配体,以诱导底层间充质细胞形成真皮凝聚物[36],最终形成皮肤毛囊的毛乳头,而毛乳头是毛囊生长发育的调控中心。在胚胎期第7~9天发育过程中,E7me和E9me差异基因在Wnt信号通路上并没有富集,但是上皮和间充质共同上下调基因富集在Wnt信号通路上,这可能与分析过程中数据样本的选取有关,并不代表E7me和E9me差异基因在Wnt信号通路上没有富集。
4 结 论
本研究通过对GSE62882的数据集中部分数据进行生物信息学分析,结果表明在胚胎期第7~9天上皮和间充质共同上下调基因主要富集在Wnt信号通路上,表明上皮和间充质通过Wnt信号诱导皮肤毛囊的生长发育,为进一步探究皮肤上皮和间充质在毛囊生长发育中的作用提供理论参考。
