毛囊发生过程及调控方式的研究进展
2022-09-14封洋
封 洋
(甘肃农业大学动物科学技术学院,甘肃兰州 730070)
毛囊(Hair Follicle,HF)为皮肤的附属器官,由初级毛囊(Primary Follicle,PF)和次级毛囊(Secondary Follicle,SF)构成,初级毛囊着生有髓的针毛,次级毛囊着生无髓的绒毛。毛皮动物的针毛粗且硬,突出于绒毛,绒毛细而软,提高毛囊发育水平可增加毛密度,对毛用动物生产有重大意义。毛囊是唯一的终生呈周期性生长的器官,其发育具有可调控性,目前对于绒山羊、水貂、獭兔等毛皮动物毛囊调控的研究主要集中在出生后动物的毛囊发育调控,主要是针对毛囊周期进行研究,提高效果有限,因此需要从其他方向探寻提高毛囊密度的方法。哺乳动物的毛囊结构类似,发育过程也相似,可分为胚胎期发育(发生)和出生后发育。本文综述了胚胎时期毛囊发生过程、发育周期、调控通路等,提出胚胎时期是毛囊发育的重要时期,此期间的调控能够影响毛囊命运,以期为提高经济动物毛密度提供新的思路和观点。
1 毛囊的发生过程及调控的信号通路
1.1 毛囊发生过程 动物毛囊发生在胚胎时期,毛囊发育与胚胎表皮分层有着密切联系。经典学说认为毛囊发生分为3 个阶段,即毛基板的形成(Hair Placode Formation)、毛囊器官的形成(Hair Follicle Organogenesis)和细胞的分化(Cytodifferentiation)(图1)。2019 年Saxena 等提出,按照毛囊内不同类型细胞的分化时期作为毛囊发生阶段的划分方法,共分为10 个阶段,更详细地阐述了毛囊发生过程中毛囊内细胞以及毛囊形态的变化。哺乳动物的毛囊发生规律相似,目前研究主要集中在人类和小鼠上,下文以小鼠毛囊发生过程为例来阐述。

图1 胚胎阶段毛囊的发生[6]
小鼠妊娠期为21 d,无论用以上哪种方法划分阶段,学者们都认为毛囊发生始于胚胎龄12 d 左右,结束于17 d 左右。由图2 可知,在第0 阶段(胎龄12.5 d之前),表皮细胞为一层均匀的细胞,没有任何标志的特殊形态(图2-a);在第1 阶段(第0 阶段结束到胎龄13.5 d),来自真皮的信号诱导表皮细胞开始形成毛囊基板(Placode,Pc)结构,Pc 结构形成之前,还有一个Pc 前体细胞(pre-Pc)阶段,在这个阶段形态学上没有变化,但是许多信号和基因已经开始发挥作用,这标志着毛囊发生已经开始(图2-b);在第2 阶段(第1 阶段结束到胎龄14 d),pre-Pc 建立后激发了附近成纤维细胞聚集,这些细胞处于向真皮聚集(Dermal Condensate,DC)的过渡阶段(图2-c),即pre-DC 细胞阶段;在第3 阶段(第2 阶段结束到胎龄14.5 d),Pc 结构细胞已经可以明显与其他表皮细胞区分开来,它更厚密、更大,并垂直于角化细胞,内陷于真皮细胞,前缘有基膜包裹,DC 在这个时期可以被认为是一个早期的聚集细胞群,在这个阶段DC 比pre-DC 包含更多的细胞,它开始沿着前-后(A-P)轴极化(图2-d);在第4 阶段(第3 阶段结束到胎龄15 d),Pc 结构慢慢变长并凹进入真皮层,形成毛基质(Hair Germ)(图2-e),而DC 细胞变得更致密,极化变得更加突出,为即将成为真皮毛乳头(Dermal Papilla)做准备。

图2 毛囊发生的第0 到第4 阶段[7]
在第5 阶段(第4 阶段结束到胎龄15.5 d),表皮细胞伸长,毛囊继续向真皮层生长,将来的毛囊轴基本已经定位好,此阶段已经从毛基质期到了毛钉期(Hair Peg)。第0 阶段到第5 阶段都是属于早期的毛囊发生阶段,从胎龄15.5 d 开始进入到第6 阶段,这段时间属于毛囊发生晚期,在这期间毛囊正式形成。在第 6阶段(第5 阶段结束到胎龄16 d),毛钉形态上更接近成熟的毛囊,在毛钉离DC 最近的基底部开始逐渐转化为球形,有大概一半以上的DC 细胞被周围的Matrix细胞包裹,这个时期DC 形态已经越来越接近真皮毛乳头,并且在此阶段内根鞘开始形成。在第7 阶段,内根鞘通过正在发育的毛囊向上生长,毛管开始发育,这阶段的DC 细胞几乎全部被周围的 matrix 细胞包裹住。在第8 阶段,毛囊不断生长,其根部已经延伸到真皮下部的边界,此时期毛管清晰可见。在第9 阶段,毛干顶端从内根鞘进入到毛管中,真皮毛乳头逐渐变窄。在第10 阶段,毛囊达到最大长度,它延伸到表皮组织中,此时毛干从表皮中延伸出来。第6-7 阶段发生的时间是在第5 阶段结束到胎龄16 d,8-10 阶段发生时间是从第7 阶段结束到胎龄17.5 d。10 个阶段完成意味着在胚胎阶段毛囊发生的结束,这个毛囊发育过程以及阶段的划分适用于哺乳动物,只是由于妊娠期的不同,每个阶段开始和结束的时间略有差异。
毛囊发生过程在胚胎时期已经全部完成,初级毛囊和次级毛囊已经固定在真皮层中,在此时期动物的毛囊密度基本已经确定。
1.2 胚胎时期调控毛囊发生的信号通路 在毛囊发生过程中,能够调控毛囊发生和生长的信号通路有很多,在小鼠中发现Wnt、Eda/Edar、Fgf、Bmp、Shh 和TGF信号通路等对毛囊具有重要作用。
1.2.1 Wnt/-catenin 信号通路对毛囊的调控作用 Wnt/-catenin 信号通路在毛囊形成过程中起着非常重要的作用。Wnt10b 表达和Wnt 信号定位于胎儿时期毛囊新生阶段1 和2 Pc 前体中,这是对于Pc 诱导必不可少的通路,表皮中-catenin 蛋白可以阻止Pc 形成。同样,Wnt 抑制剂Dkk1(通常在早期HFs 周围的真皮成纤维细胞中发现)和Dkk2 的表达也阻止了Pc 形成。除了驱动Pc 启动,局部表皮Wnt 活性也是Pc 形成所必需的,在Pc 前体(第1 和第2 阶段)到Pc(第3 阶段)的过渡过程中,其可以协调细胞迁移,早期Pc 形态发生期间的活体成像和跟踪细胞分裂证明Wnt 和Eda信号通过定向迁移介导Pc 形成重新排布。
1.2.2 Eda 信号通路对毛囊发生的调控 当TNF 家族成员膜配体Eda 与其受体Edar 结合后,下游NF-B 被激活进而触发Pc 结构形成。在毛囊发生过程中,Eda在整个表皮中均表达,而Edar 的表达位置仅限于第1 和第2 阶段Pc 前体细胞和第3 阶段后的Pc 结构。Edar 是Pc 前体出现的标志之一,Eda/Edar 信号对于Pc 前体诱导HF 有一定作用。在缺失Eda 的情况下,Pc 前体一直停留在第1 阶段到第2 阶段,没有进一步发展到第3 阶段形成Pc。此时可以形成部分 HFs 结构,但下一步分化的HFs 根部受损,无法弯曲,这表明Eda 信号在发干弯曲的分子机制中发挥着重要作用。在小鼠上试验表明,如果EdaA1 信号缺失,小鼠只会长出细毛。
1.2.3 Shh 信号通路对毛囊发生的调控 在发育过程中,Shh 对毛发生长和形态发生至关重要。Shh 信号通路在 Pc 前体和Pc 细胞的所有阶段均表达,Shh 信号通过Patched(Ptc)和Multipass Transmembrane Protein Smoothened(Smo)的联合受体复合物传递,其受体在表皮和真皮中均表达。Shh 缺失小鼠在第4 阶段Pc 和DC 均正常诱导,但HF 发育停滞。皮肤特异性切除Shh 信号通路受体中的Smo 会导致皮肤Shh 信号传导受到抑制,进而影响DC 中Noggin 的表达。Noggin 对DC 的维持非常重要,其缺失后皮肤中的Pc 不能表达Shh mRNA 和蛋白,Noggin 是介导对毛囊发生同样重要的BMP 信号通路的因子,这一系列说明Shh 信号和BMP 信号在HFs 的发生中建立了正反馈回路。此外,Shh 信号对早期HF 形态发生中(第3阶段到第4 阶段过渡期间)形成极化也起着重要作用。
1.2.4 Fgf20 信号通路对毛囊发生的调控 Fgf20 信号是目前唯一确定的DC 形成的必须表皮信号,DC 结构的形成是通过依赖Fgf20 信号引导的细胞迁移和聚集而不是增殖来实现的,当被敲除后,无法形成DC结构,次级毛囊也大量减少。最近的研究表明在发生毛囊细胞聚集之前,成纤维细胞需要Fgf20 来进一步转化为DC 结构。另外,试验发现,Fgf20 发生突变会导致Pc 结构的形态由圆形转变为条纹状,这说明Fgf20 对Pc 的形态也起着至关重要的作用。虽然Fgf20信号是Wnt 和Eda 信号的下游靶点,但敲除后Pc 中的Edar 表达水平也显著降低。综上,Fgf20 是Pc 前体(第1 阶段)向真皮发出的第1 个上皮信号,负责促进成纤维细胞向DC 的转化,接着Fgf20 指示前Pc 细胞迁移和聚集,形成聚集的DC。
1.2.5 TGF信号对毛囊发生的调控 TGF信号被认为是毛囊向下生长的关键,TGF2 由真皮层分泌,并对上皮的受体表达起重要作用。TGF2 缺失的小鼠的毛囊向下生长过程中受到抑制,这说明此过程需要TGF信号,若把TGF受体敲除,则毛囊较少且生长迟缓。另外,也有研究表明Eda 信号通路的传导是通过激活TGF信号传导来完成的。
1.2.6 BMP 信号通路对毛囊发生的调控 Wnt、Eda 和Fgf20 信号可以促进毛囊发生,BMP 信号起到的则是抑制作用。在毛囊形态发生过程中,配体Bmp2 和Bmp4 在Pc 和DC 中表达,在毛囊周围的上皮细胞中也检测到了BMP 受体的表达,其作用也是抑制毛囊发生。而BMP 抑制剂Noggin 在DC 中富集,其可以抑制BMP 信号激活和促进Pc 的形成。
综上,在第一次表皮信号启动毛囊发生后,Wnt 和Eda 信号被激活促进Pc 前体的形成和Pc 结构的建立。在Fgf20 等信号通路的诱导下,DC 结构逐渐成型并开始聚集成熟的DC 细胞。同时,来自Pc 和DC 的抑制信号例如BMP 信号通路等抑制了毛囊的形成。来自DC 的真皮信号随后促进Pc 细胞的增殖以使毛囊向下生长,通过Shh 和TGF信号通路持续的相互作用,进一步促进初始毛囊结构向成熟HF 的形成发展。在胎儿期结束前,在各种通路和基因的共同调控下,毛囊发生完成,在胚胎时期毛囊的密度直接决定着个体的毛密度,也决定着毛囊的命运。
2 出生后毛囊的发育
毛囊发生过程在胚胎时期已经完成,出生后毛囊也是可以再次变化的,这时期的变化呈一定的周期性。这个周期性主要是与毛囊干细胞(Hair Follicle Stem Cell,HFSC)的周期性生长、分化、增殖和DP 细胞之间的信号互相关联有关。传统研究通常认为毛囊是有周期的,分为生长期(Anagen)、退行期(Catagen)和休止期(Telogen)。生长期的毛囊处于最活跃的阶段,DP变大并发出增殖信号,HFSC 迅速增殖,分化出新的毛干,同时DP 向真皮层方向生长,毛囊形态结构处于动态的平衡,这期间也是毛发状态最好的时期;退行期的真皮毛乳头会回缩到外根鞘隆起位置,伴随着外根鞘上皮细胞大量调亡,毛干的生长也停止,并分化形成杵状毛发;退行期过后在休止期毛囊细胞活动减弱,毛干脱落,休止期的后期毛球再生,HFSC 重新开始增殖,毛囊相关调控因子开始活跃,为下一个生长期做准备。
毛囊周而复始的周期过程伴随着哺乳动物的一生,虽然由于物种不同,各个时期略有差异,甚至在同一个物种的不同部位毛囊的周期性也有差异,但是毛囊的周期规律在哺乳动物中普遍适用。表1 对常见几种哺乳动物的毛囊周期生长情况进行了总结。
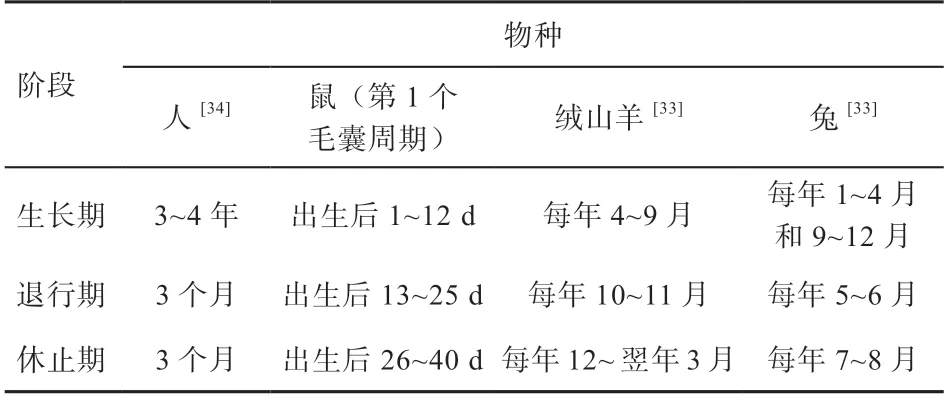
表1 哺乳动物毛囊生长周期
毛囊状态直接决定着毛品质,在不同周期内毛品质不同。通常在生长期的毛密度较高,品质较好,而退行期绒毛会脱落,此时期毛密度较低。有研究表明,初级毛囊几乎不会随着毛囊发育周期呈现出变化,次级毛囊才会有完整的周期性变化。也就是说出生后毛囊的周期发育主要是指次级毛囊,对出生后的动物毛囊干预和调控主要是与毛囊发育周期相关,通过对毛囊生长期的延长或者休止期的缩短调控进而影响次级毛囊密度。
3 关于哺乳出生前和出生后毛囊调控的研究
3.1 出生后毛囊的调控 对于哺乳动物来说,调控毛囊发育一般从营养、激素等方面来着手,目前也有研究表明压力减少也可以促进毛发生长。在营养方面,蛋氨酸、胱氨酸、半胱氨酸对毛发的发育起着重要作用,发现蛋氨酸可以通过刺激Wnt/-catenin 信号通路显著增加獭兔背部的毛囊密度,并延长毛干的生长期。傅祥超等研究发现蛋氨酸和半胱氨酸可显著提高獭兔毛密度,但其认为从商品分级评分的角度来考虑,含硫氨基酸不能显著提高被毛评分,所以仅从增加含硫氨基酸添加水平的角度来大幅度提高被毛品质不太可能。另外,B 族维生素对毛囊发育也有促进作用,核黄素缺乏会导致毛发脱落,在獭兔饲养中,也有研究发现适量添加核黄素会一定程度促进毛囊发育。除此之外,一些激素的应用也可以调节毛囊发育,以人为例,雄激素对头皮区域的毛囊生长起到抑制作用,导致雄激素脱发,这也是人类脱发的最主要的原因,因此用一些雄激素阻断剂可以减少脱发发生。此外,催乳素对哺乳动物毛发生长的影响已被深入研究,它对毛干伸长有抑制作用并能诱导毛囊退行期提前。另外,促黄体素还通过减少Ki-67 阳性细胞的数量和增加TUNEL+细胞的数量,在HFs 角质形成细胞的增殖和凋亡中发挥重要作用。褪黑激素(Melatonin,MT)是一种重要的促进毛囊发育的激素,可以通过刺激生长期的毛囊影响而调节毛发生长。在绒山羊研究中发现,褪黑激素在体外和体内试验中都可促进次级毛囊生长,使羊绒产量提高,活跃的次级毛囊数量增加。在獭兔研究中也发现褪黑激素可以促进次级毛囊发育。关于毛囊调控的研究都证实,出生后的动物毛囊调控只对次级毛囊有显著作用,而对初级毛囊的密度影响要小于次级毛囊。原因是出生后动物对于毛囊的调控主要在影响毛囊周期,而毛囊的发育在胚胎期间已经完成,所以对毛囊数量的影响程度有限。
3.2 胚胎时期毛囊发生的调控 胎儿在子宫内,表皮上皮和间质真皮的相互作用形成了毛囊,后期的各种营养和外界条件等干预和处理对次级毛囊的周期可以起到改善作用,但对于初级毛囊效果甚微。胚胎发生过程中毛囊密度在出生后将永久固定,由于毛囊是固定在皮下组织中的器官,在胎儿时期,毛囊的位置也是固定并伴随终身。因此,从胎儿时期着手去提高初级毛囊密度可以从根本上提高毛皮动物毛囊密度。
胎盘是母体妊娠过程中的重要组成部分,它连接着母体和胎儿,起着输送营养物质,排出代谢产物,并分泌相关激素维持妊娠的重要作用,子宫的微环境与胎儿发育水平息息相关。在胎儿时期,作者在对獭兔在孕期母体添加褪黑激素,发现仔兔出生后的初级毛囊和次级毛囊都显著增加,并发现褪黑激素受体基因表达的改变,以及Wnt 和MAPK 等与毛囊发育相关信号通路的改变(数据未发表);也有报道称在妊娠期绒山羊的饲料中添加纳米硒可以改善胎儿的毛囊发育并促进其生长发育;妊娠期妇女的饮食营养水平可直接影响出生婴儿毛囊中等位基因的DNA 的甲基化程度,这说明孕期的影响与毛囊发育直接相关;在禽类上,一项研究蛋氨酸(Met)在雏鸡胚胎期对羽毛的毛囊和羽毛发育的调节作用中发现,-Met 可以通过激活Wnt/-catenin 信号传导提高羽毛的发育水平,从而促进羽毛的生长。
关于母体调控毛囊的研究报道很少,但通过有限的报道可以确定,在胚胎时期的干预和调控可以对毛囊发育起到显著的改善作用。图3 为胚胎时期对毛囊调控的假设机制图,在胚胎时期通过营养、激素或者其他条件进行调控,调控元素通过母体垂直传递给胎儿,在各种通路的共同作用下,对Pc 和DC 的形成起到促进作用,进而促进胎儿毛囊生长,此正向作用在动物体内的表现是长期甚至一生的。在毛囊发生时期的调控可以影响毛囊的命运,进而提高毛皮质量,这对于毛皮动物意义重大。

图3 调控哺乳动物胚胎时期毛囊发育假设机制图
4 总结与展望
被毛作为哺乳动物重要的经济性状,具有较大的潜在研究价值,毛囊作为产生毛发的主要器官,是皮肤的重要附属物,提高毛囊发育水平是畜牧工作者研究的问题。目前的研究几乎都是在出生后对动物的毛囊发育进行调控,通过毛囊的发生过程,发生的调控机制的研究可以发现,毛囊主要发生的时间点在妊娠中后期,而出生后所谓的毛囊发育只是次级毛囊的周期变化,从本文中可知,胚胎时期哺乳动物的毛囊发生影响着毛囊细胞命运,因此,从胚胎时期对毛囊的调控尤为重要,也是目前研究中缺失的部分。随着各种研究技术的发展,提高毛囊发育水平和研究机制已经成为可能,本文观点的提出可为毛皮动物的育种工作和提高动物毛品质提供新的研究方向和思路。
