北部湾沿岸三种常见赤潮藻类对浮游微食物网主要类群的影响
2022-08-15陆家昌陆敏桢赖俊翔庄军莲
陆家昌,陆敏桢,李 杰,赖俊翔,庄军莲
(广西科学院、广西北部湾海洋研究中心、广西近海海洋环境科学重点实验室,广西 南宁 530007)
浮游植物是海洋初级生产者,通过光合作用将水体中的无机营养转化成颗粒有机物。在浮游植物的生产中,约有一半的初级生产以溶解有机物的形式释放到水体中[1]。浮游细菌利用水体中的溶解有机物进行生产,并通过微食物网中微型鞭毛虫(nanoflagellate, NF)和纤毛虫等原生动物的摄食,将细菌的生产向更高营养级传递,从而使得这部分初级生产重新回到经典食物网中[2-3]。异养细菌、微型鞭毛虫和纤毛虫是微食物网中的重要组成类群[4],自然水体中,浮游植物与微食物网各类群之间关系复杂。一方面,浮游细菌和大部分含色素体微型鞭毛虫(pigmented nanoflagellate, PNF)可以与藻类竞争营养盐[5];另一方面,异养细菌的分解和纤毛虫等原生动物的排泄又是营养盐再生的重要途径,特别是在贫营养海区,这一途径又是浮游植物生长所需营养盐的重要来源[6-8]。此外,异养微型鞭毛虫(heterotrophic nanoflagellate, HNF)和纤毛虫又可以作为捕食者,直接以合适粒径的浮游植物为食,对浮游植物形成下行控制[9-10]。
适宜条件下,浮游植物暴发性增殖形成藻华,积累大量颗粒有机物。藻华衰亡后,沉降到底部,其中的有机物通过异养细菌的分解进入微食物网,在藻华的衰亡、分解过程中,微食物网发挥着重要作用[11]。此外,微食物网的结构和功能也是近岸浮游植物群落变化的重要调节因素,其在营养盐循环中的作用对藻华的形成具有重要影响[12]。因此,了解不同赤潮种与微食物网结构和功能之间的关系,有助于认识藻华形成和衰亡过程中微食物网的功能和作用。
不同类群的浮游植物的生理生态特性不同,其对光照、温度、营养盐等环境因子的适应性也有差异,因此不同种类的藻华的暴发也具有一定时序性,比如,甲藻赤潮一般出现在硅藻藻华之后,而球形棕囊藻(Phaeocystisglobosa)赤潮多出现在较寒冷的季节[13-15]。然而,这种时序性并不是一成不变的。野外调查发现,红色哈卡藻(Akashiwosanguinea)、球形棕囊藻和中肋骨条藻(Skeletonemacostatum)都曾在水温20 ℃左右形成藻华[16-18]。此外,不同原因种藻华对海洋环境的影响也不相同。一般认为,除可能分解耗氧,在海洋底部形成缺氧区外,大部分的硅藻藻华对海洋环境是无害的,甚至有些种类硅藻的藻华对渔业生产还可能是有益的[19]。而红色哈卡藻和球形棕囊藻会分泌毒素、在海水表面形成泡沫,对海洋环境和渔业生产造成危害,形成的藻华被认为是有害藻华[20-22]。本研究以自然水体中的微食物网为研究对象,以中肋骨条藻这一北部湾近岸常见的硅藻为对照,通过向自然水体中引入浮游植物的方法,分别比较有毒有害的红色哈卡藻、球形棕囊藻与中肋骨条藻对微食物网主要类群影响的差异。本研究有助于认识微食物网在北部湾近岸有毒有害藻华形成和衰亡过程中的作用。
1 材料与方法
1.1 实验设计
实验所需微食物网主要生物类群通过采集自然海水的方式获得。海水采样体积为100 L,于2019年3月19日采自北部湾近岸海域(21.528 144°N,108.168 819°E),采样站位水深为5.3 m,采水深度为0.5 m,采样时水温为19.2 ℃,盐度为30.5。自然海水中的叶绿素a浓度为3.62 μg /L,浮游植物以硅藻为主。
在重力作用下以200 μm孔径筛绢反复过滤自然海水水样3次去除捕食者后,分装到容积为15 L的透明塑料瓶中,每个塑料瓶装10 L。实验所用中肋骨条藻、红色哈卡藻和球形棕囊藻游离单细胞均由广西北部湾海洋研究中心藻种室提供,培养温度为20.0 ℃,光照强度4 500 lx,光照和黑暗时间为12 h∶12 h。将中肋骨条藻、红色哈卡藻和球形棕囊藻游离单细胞按8 μg/L的叶绿素a终浓度引入实验水体,3种藻类接种后,水体中最终的细胞密度分别为(1.83±0.37)×104、(4.75±0.45)×102、(5.23±1.23)×105cells/mL。每个处理设3个平行。实验以添加中肋骨条藻这一北部湾近岸海域常见的硅藻为对照组,分别比较有毒有害的红色哈卡藻、球形棕囊藻与中肋骨条藻对微食物网主要类群影响的差异。由于过滤去除捕食者后,培养过程中自然海水中会自发形成优势种,因此实验不设置空白海水组,而以第0 天的结果作为代表。为去除原f/2培养液[23]的干扰,微藻在进行实验前先在3 500 r/min下离心5 min,再以不含营养盐的人工海水再悬浮。离心前,先通过预实验确定转速和离心时间,以确保离心不会对藻细胞造成损伤。实验在控温培养箱中进行,实验温度为20 ℃,光照强度(4 290±730) lx,设置光照和黑暗时间为12 h∶12 h。培养2 d后,添加藻类均成为绝对优势种,显微镜下很难再找到原来的藻类。实验用球形棕囊藻为游离单细胞藻,实验过程中未见形成囊体,流式细胞分析表明其为球形棕囊藻的单细胞体。实验于每天上午9:00左右采样,采样前先轻轻摇晃,混合均匀。分别取2 mL和10 mL水样用于细菌和微型鞭毛虫计数,取50 mL水样用于营养盐浓度测定,取100 mL水样进行藻细胞计数,每个样品取3个平行样。由于培养体积有限,实验仅在开始和结束时对纤毛虫样品进行取样,取样体积为1.5 L。
1.2 细菌和微型鞭毛虫的计数
用于细菌和微型鞭毛虫分析的水样先以终浓度为0.5%(体积分数)的戊二醛进行固定,并以终浓度为10 μg/mL的4’,6-二脒基-2-苯基吲哚避光条件下染色10 min,而后低压过滤(<13.33 kPa)到黑膜(聚碳酸酯滤膜,孔径为0.22 μm)上制成玻片,于-20 ℃保存。玻片在荧光显微镜下进行镜检,紫外激发光下计数细菌数量,计数数量不少于300个。紫外激发光下找到微型鞭毛虫细胞,再切换到蓝光下确认该鞭毛虫是否有自发红色荧光,有自发红色荧光的是含色素体微型鞭毛虫,没有的是异养微型鞭毛虫。通过显微镜(Nikon 80i)自带的软件系统(NIS-elements V3)对微型鞭毛虫的细胞粒径进行测量,根据粒径的大小,分别将异养微型鞭毛虫和含色素体微型鞭毛虫划分为2~5、>5~10、>10~20 μm 3个类群。为确保镜检结果的可靠性,需保证计数的微型鞭毛虫的个体总数不少于100个[24]。
1.3 纤毛虫和藻细胞的计数
实验前后,不同培养瓶中各取1.5 L水样装入塑料样品瓶中,以Lugol’s液固定,终浓度为1%(体积分数)。样品于阴凉处自然沉淀48 h后,以虹吸管将上清液小心吸出,浓缩到剩余约120 mL。分两次吸取30 mL浓缩液于倒置显微镜下观察计数纤毛虫的个体密度。本研究不对纤毛虫种类进行鉴定,而只对其个体密度进行计数。另外,每天从培养瓶中取5 mL水样,加Lugol’s液固定后直接于倒置显微镜下观察计数藻细胞密度。
1.4 营养盐浓度的测定
所有与营养盐浓度测定相关的实验器具均以1 mol /L的HCl进行浸泡,并用去离子水洗净。水样经0.45 μm孔径的醋酸纤维滤膜过滤后于-20 ℃冰箱中储存。实验结束后应用荷兰Skalar公司生产的营养盐自动分析仪对溶解无机氮(dissolved inorganic nitrogen, DIN)和溶解活性磷酸盐浓度进行测定,其中溶解无机氮是硝酸盐、亚硝酸盐和氨氮3项之和。
1.5 数据处理
数据以平均值±标准差的形式表示,使用SPSS 16.0软件对数据进行统计学处理,置信区间设为95%,组间参数比较在符合正态分布或近似正态分布及方差整齐前提下进行。时间序列的培养实验中,营养盐浓度、微型鞭毛虫个体密度和细菌的细胞密度的变化都不是独立的,它们是前一个时间点的结果的延续,前一个时间点的结果对其后的结果存在影响。对于具有时间序列的延续性数据,采用重复测量方差分析(repeated measured ANOVA)法进行分析。实验前后各组间纤毛虫个体密度的差异采用Turkey HSD进行比较。
2 结果与讨论
2.1 浮游植物的密度变化
3种藻细胞的密度均在接种后迅速增加,中肋骨条藻的细胞密度在接种第4天达到峰值(7.02±0.08)×105cells/mL,红色哈卡藻在第5天达到峰值(11.65±0.08)×102cells/mL,而球形棕囊藻游离单细胞的迅速增殖期只有2 d,峰值为(17.25±1.16)×105cells/mL。中肋骨条藻和红色哈卡藻细胞在经历4~5 d的增殖后迅速衰亡,但球形棕囊藻游离单细胞在2 d的迅速增殖后并未衰亡,其细胞密度一直维持相对稳定[(18.92±1.42)×105cells/mL],直至实验结束(图1)。

图1 不同添加组浮游植物细胞密度的变化Fig. 1 Variations of the phytoplankton density among different treatments
2.2 营养盐浓度的变化
实验起始时,水体中的磷酸盐浓度为(0.21±0.00) μmol/L,溶解无机氮的浓度为(51.52±0.68)μmol/L。实验水体是一个氮过剩,而磷缺乏的系统。浮游植物的生长受磷酸盐的限制。不同浮游植物对营养盐的利用不同,球形棕囊藻添加组在第二天即将磷酸盐消耗殆尽,第二天后系统中的磷酸盐浓度一直维持在极低水平(约为0.06 μmol/L)。红色哈卡藻虽然在第5天后开始衰败,但水体中的磷酸盐浓度从实验开始就一直降低,直至实验结束时,水体中磷酸盐浓度降至0.09 μmol/L。实验开始后中肋骨条藻快速增殖,迅速消耗水中磷酸盐,在第5天时,磷酸盐浓度降至最低,与其他两种藻不同,中肋骨条藻在衰亡后迅速向水体中释放磷酸盐。第5天后水体中磷酸盐浓度开始升高,实验结束时,水体中磷酸盐浓度与实验起始时比,恢复了79%(0.17 μmol /L,图2)。
球形棕囊藻添加组中溶解无机氮浓度显著低于其他两组(P<0.05),而中肋骨条藻和红色哈卡藻添加组中溶解无机氮浓度无显著差异(P>0.05)。与实验初始溶解无机氮浓度相比,实验结束时,红色哈卡藻添加组中溶解无机氮浓度降低了19.80%,中肋骨条藻添加组降低了7.37%。中肋骨条藻和红色哈卡藻添加组中溶解无机氮浓度在前7天类似。除了第4天大幅变动外,球形棕囊藻添加组中溶解无机氮浓度的变化与磷酸盐浓度变化类似,在迅速降低后,该组溶解无机氮在低浓度状态下保持相对平衡,实验结束时,球形棕囊藻添加组溶解无机氮浓度降低了90.21%(图2)。
2.3 浮游细菌的密度变化
实验起始时,水体中浮游细菌的细胞密度为(7.97±0.99)×106cells/mL,实验结束时,各实验组的浮游细菌的细胞密度均有所提高。此时,中肋骨条藻、红色哈卡藻和球形棕囊藻添加组浮游细菌细胞密度分别为(21.93±2.20)×106、(10.23±2.35)×106、(13.28±0.96)×106cells/mL。实验过程中,中肋骨条藻添加组的浮游细菌的细胞密度显著高于其他两组(P<0.05),而球形棕囊藻和红色哈卡藻添加组浮游细菌的细胞密度无显著差异(P>0.05,图3)。
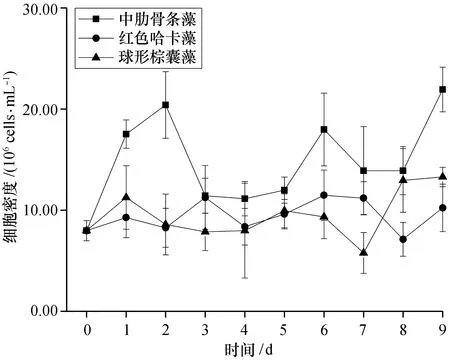
图3 不同添加组异养细菌细胞密度的变化Fig. 3 Variations of the density of heterotrophic bacteriaamong different treatments
2.4 微型鞭毛虫的密度变化
红色哈卡藻添加组的含色素体微型鞭毛虫的密度高于中肋骨条藻和球形棕囊藻添加组。红色哈卡藻添加组的异养微型鞭毛虫和含色素体微型鞭毛虫的密度均显著高于中肋骨条藻和球形棕囊藻添加组(P<0.05)。虽然球形棕囊藻添加组异养微型鞭毛虫的密度与中肋骨条藻无显著差异(P>0.05),但是球形棕囊藻添加组含色素体微型鞭毛虫的密度显著低于中肋骨条藻(P<0.05,图4)。较之中肋骨条藻,红色哈卡藻添加组2~5 μm 异养微型鞭毛虫、>5~10 μm异养微型鞭毛虫、2~5 μm 含色素体微型鞭毛虫和>5~10 μm 含色素体微型鞭毛虫的密度显著提高(P<0.05,图5)。虽然球形棕囊藻添加组2~5 μm 异养微型鞭毛虫密度与中肋骨条藻添加组无显著差异(P>0.05),甚至除个别时间点外,球形棕囊藻添加组2~5 μm 异养微型鞭毛虫密度明显高于中肋骨条藻添加组,但是,球形棕囊藻添加组>5~10 μm 异养微型鞭毛虫、2~5 μm 含色素体微型鞭毛虫和>5~10 μm 含色素体微型鞭毛虫密度显著低于中肋骨条藻添加组(P<0.05,图5)。

图4 不同添加组异养微型鞭毛虫和含色素体微型鞭毛虫的个体密度变化Fig. 4 Variations of HNF and PNF densities among different treatments

图5 不同添加组不同粒径微型鞭毛虫的个体密度变化Fig. 5 Size-dependent variations of nanoflagellate density among different treatments
除第2天外,无论是生长还是衰退,所有组的HNF/PNF的比值均大于1。实验开始时,实验水体处于磷限制状态。此时,HNF/PNF的比值为12.7±0.2,表明营养盐限制条件下,微型鞭毛虫群落偏向于异养。虽然统计结果显示无显著差异(P>0.05),但在藻类生长阶段,红色哈卡藻添加组比中肋骨条藻添加组更偏向异养,而在衰亡阶段,红色哈卡藻添加组和中肋骨条藻添加组无明显差异。较之红色哈卡藻和中肋骨条藻,球形棕囊藻添加组的微型鞭毛虫群落显著偏向于异养(P<0.05,图6)。
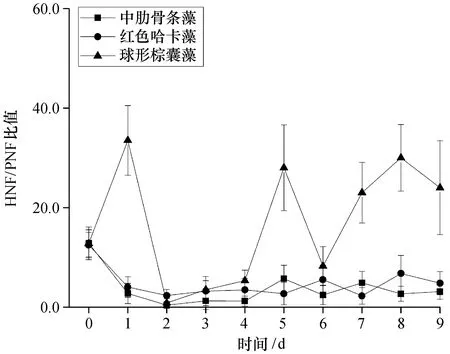
图6 不同添加组HNF和PNF的比值Fig. 6 Ratio of HNF to PNF among different treatments
2.5 纤毛虫的密度比较
实验开始时,中肋骨条藻、红色哈卡藻和球形棕囊藻添加组纤毛虫的个体密度分别为(12.12±3.56)×103、(13.43±1.01)×103、(13.79±6.52)×103ind/L,各组之间无显著差异(P>0.05)。由于已经过滤去除捕食者,较之实验开始时,实验结束时纤毛虫的密度均显著提高(P<0.05),实验结束时中肋骨条藻、红色哈卡藻和球形棕囊藻添加组纤毛虫的个体密度分别为(1 192.92±714.96)×103、(1 480.83±214.95)×103、(142.88 ±34.78)×103ind/L。各组间纤毛虫密度差异显著(P<0.05),纤毛虫密度红色哈卡藻添加组>中肋骨条藻添加组>球形棕囊藻添加组(图7)。

图7 不同添加组纤毛虫个体密度的比较Fig. 7 Comparisons on the density of ciliates among different treatments“*”表示同种藻类添加组在实验开始和结束时纤毛虫密度差异显著,P<0.05;小写字母表示实验开始时,各藻类添加组间纤毛虫密度的差异,字母相同,表示差异不显著,P>0.05;大写字母表示实验结束时,各藻类添加组间纤毛虫密度的差异,字母不同表示差异显著,P<0.05。
2.6 讨论
2.6.1 球形棕囊藻对微食物网的影响 实验用球形棕囊藻游离单细胞为单倍体。棕囊藻属的种类可以形成单倍体以适应贫营养条件[20,25],实验结果也表明在磷酸盐几乎耗尽的情况下,球形棕囊藻游离单细胞数量仍维持稳定,并未衰亡(图1、2),这也与我们在藻种室的日常培养中观察到的球形棕囊藻游离细胞在指数生长期后仍然保持长时间稳定的现象相符。较之大粒径的硅藻和甲藻,小粒径浮游植物在对营养盐的竞争中更具优势[26]。实验中,球形棕囊藻添加组的磷酸盐在2 d后降到最低值,而中肋骨条藻添加组需要5 d,红色哈卡藻添加组需要9 d(图2)。实验所用球形棕囊藻细胞粒径微小,只有4 μm左右,可与含色素体微型鞭毛虫竞争营养盐,从而降低微食物网中含色素体微型鞭毛虫的密度(图4)。由于含色素体微型鞭毛虫密度的降低远大于异养微型鞭毛虫密度的降低(图4、6),从而导致球形棕囊藻添加组微型鞭毛虫的营养结构(HNF/PNF)偏向异养(图6)。实验中,各组HNF/PNF的比值均大于1,通常情况下,富营养水体中该值小于1[27-28];就绝对值而言,实验水体是一个氮丰富的系统,但又同时是一个磷限制的系统(图2),磷的限制可能是导致这一现象的一个重要原因。球形棕囊藻不仅缺乏后生动物生长所必须的脂肪酸[比如二十碳五烯酸 (EPA)、二十二碳六烯酸 (DHA) 等],还可产生二甲基硫、丙烯酸等有毒有害物质,导致浮游动物中毒死亡[29-31]。虽然球形棕囊藻对原生动物的影响尚有争议[32-33],但不能排除球形棕囊藻也可能对原生动物存在毒害作用,从而导致球形棕囊藻添加组纤毛虫的密度在实验结束时显著低于红色哈卡藻和中肋骨条藻添加组(P<0.05,图7)。虽然统计结果差异不显著(P>0.05),但实验后期,球形棕囊藻添加组>5 ~10 μm 异养微型鞭毛虫和>10~20 μm 异养微型鞭毛虫的密度明显低于中肋骨条藻和红色哈卡藻添加组(图5),这可能也与该粒径范围的异养微型鞭毛虫摄食球形棕囊藻游离单细胞,从而遭受毒害作用有关。
2.6.2 红色哈卡藻对微食物网的影响 磷限制水体中,实验赤潮藻类的生长主要受磷酸盐浓度的影响(图1、2)。与其他两种藻类类似,红色哈卡藻增殖过程中(前5天),水体中的磷酸盐浓度也迅速减少,但减少的幅度小于球形棕囊藻和中肋骨条藻添加组(图2),这可能与红色哈卡藻的营养方式有关。红色哈卡藻不仅可以进行光合自养,也可进行吞噬异养,特别是在贫营养条件下,红色哈卡藻通过捕食补充生长所需营养物质[34-35]。实验还发现红色哈卡藻衰亡后,水体中的磷酸盐并没有像中肋骨条藻添加组一样重新释放到水体中,而是持续降低(图2)。就粒径大小而言,微型鞭毛虫远小于红色哈卡藻,红色哈卡藻可能捕食微型鞭毛虫,而异养微型鞭毛虫不可能捕食红色哈卡藻。但我们却在实验过程中观察到成群的异养微型鞭毛虫在攻击啃食正在游动的红色哈卡藻细胞。前人研究也发现确实有些微型鞭毛虫可以“围捕”红色哈卡藻,而且异养微型鞭毛虫对红色哈卡藻的摄食远大于对异养细菌和聚球藻的摄食[36]。这或者可以解释为何红色哈卡藻添加组异养微型鞭毛虫的密度显著高于中肋骨条藻和球形棕囊藻添加组(图4、5),但“围捕”红色哈卡藻的异养微型鞭毛虫的种类及其生理生态学特征还有待进一步研究。异养微型鞭毛虫对红色哈卡藻的啃食加快了红色哈卡藻的分解,释放的营养盐(主要是磷酸盐)促进了含色素体微型鞭毛虫的生长(图4、5)。异养微型鞭毛虫和含色素体微型鞭毛虫密度的显著增加意味着纤毛虫可利用的食物增加,从而使得红色哈卡藻添加组纤毛虫的密度显著高于中肋骨条藻和球形棕囊藻添加组(图7)。
2.6.3 中肋骨条藻对微食物网的影响 中肋骨条藻添加组营养盐的变化与红色哈卡藻和球形棕囊藻添加组不同,伴随着中肋骨条藻的快速增殖,水体中营养盐,特别是磷酸盐浓度迅速降低,在磷酸盐消耗殆尽后,中肋骨条藻迅速衰亡,营养盐也随之被重新释放到水体中(图1、2)。这表明微食物网,特别是微食物网中的异养细菌在中肋骨条藻衰亡藻体的分解和营养盐的再生过程中发挥重要作用。结果也确实表明,中肋骨条藻添加组的细菌密度要高于红色哈卡藻和球形棕囊藻添加组(图3)。然而,中肋骨条藻添加组中细菌的主要捕食者2~5 μm 异养微型鞭毛虫的密度却显著低于红色哈卡藻添加组,而与球形棕囊藻添加组无显著差异(图5)。而且虽然中肋骨条藻衰亡后,营养盐的释放使得该组磷酸盐的浓度在实验后期高于其他两组(图2),但除纤毛虫、2~5μm 异养微型鞭毛虫和>10~20 μm 异养微型鞭毛虫外,其他微型鞭毛虫类群,特别是含色素体微型鞭毛虫的密度也处于球形棕囊藻和红色哈卡藻添加组之间(图5),其中一个可能的解释是由于纤毛虫和>10~20 μm 异养微型鞭毛虫对<10 μm微型鞭毛虫的摄食一方面释放了异养微型鞭毛虫对细菌的摄食压力,从而促进了细菌的生长并进一步促进了细菌对中肋骨条藻的分解;另外一方面,不只是浮游细菌分解硅藻细胞释放营养盐,捕食者的摄食与代谢在营养盐再生中也发挥着非常重要的作用,而且这些捕食者对含色素体微型鞭毛虫的摄食也可能减少了含色素体微型鞭毛虫类群对营养盐的利用,从而导致水体中营养盐,特别是磷酸盐浓度的迅速恢复(图2)。
3 结论
由于不同藻类的生理生态特征不同,其对微食物网各主要类群的影响也不尽相同。球形棕囊藻游离单细胞由于个体微小,又适应低营养环境,其对微食物网的影响主要是与微食物网中的含色素体微型鞭毛虫竞争营养盐,其次,球形棕囊藻还能分泌有毒有害物质,抵御纤毛虫等原生动物的捕食。红色哈卡藻比较特殊,可被某些异养微型鞭毛虫类群摄食,从而提高微型鞭毛虫及纤毛虫的密度。与其他两种藻类相比,中肋骨条藻对微食物网主要类群的影响更容易受到水体中营养盐的调节。
