基于RNA-seq 筛选白藜芦醇致猪卵巢颗粒细胞凋亡的可变剪接基因
2022-08-13章会斌刘阳光陈一歌许黎明徐启隆殷宗俊张晓东
章会斌,韩 政,刘阳光,周 忍,陈一歌,许黎明,徐启隆,殷宗俊*,张晓东*
(1.安徽农业大学动物科技学院,安徽合肥 230036;2.地方畜禽遗传资源保护与生物育种安徽省重点实验室,安徽合肥 230036)
白藜芦醇(Resveratrol,RES)作为一种非黄酮类多酚化合物,广泛参与细胞抗氧化作用,其甲基化类似物在猪卵巢颗粒细胞(Porcine Ovarian Granulosa Cells,pGCs)中表现出抑制作用,能影响颗粒细胞雌二醇及黄体酮的产生。此外已有研究证实RES 能促进大鼠卵巢膜间质细胞凋亡,损伤颗粒细胞形态并抑制细胞增殖。可变剪接(Alternative Splicing,AS)普遍存在于高等生物体内,是一种常见的调节基因表达的机制。通过剪接基因产生多个mRNA,从而指导蛋白质发生结构及功能的多样性变化。近年来,有学者针对畜禽基因可变剪接事件的鉴定和生信分析,揭示了可变剪接在动物体内的重要生理机制。研究发现,可变剪接可以通过调控信号通路中关键基因的转录,影响机体发育,并参与疾病的发生。在猪不同发育时期睾丸组织中鉴定到差异可变剪接基因与各时期的生理状态有关,而北京鸭的皮脂中特有的可变剪接基因参与油脂合成,胸肌中发生可变剪接的基因与骨骼肌发育相关。
课题组前期已有研究指出,RES 可抑制pGCs 增殖,并对pGCs 凋亡产生影响。基于此,本研究构建了RES 诱导pGCs 凋亡模型,首次通过转录组测序(RNA-seq)及生物信息学技术鉴定分析了RES 诱导pGCs 凋亡的可变剪接事件及差异可变剪接基因,有助于在分子水平进一步认识pGCs 凋亡的具体机制。
1 材料与方法
1.1 测序样本的采集与制备 本实验所用猪卵巢采集自某商业屠宰场180 日龄的三元杂交猪。猪只屠宰后立即采集、分离卵巢,并保存在37℃含500 IU/mL 青/链霉素的无菌生理盐水中。2 h 内用保温瓶运回实验室备用。采用抽吸法收集卵泡液,并分离纯化pGCs。将RES 溶解在二甲基亚砜(DMSO)中,制备0、25、50、75、100 μmol/L RES(RES 购自上海麦克林生化科技公司,纯度≥99%)的完全培养基。将纯化后的pGCs 接种至细胞培养板,待细胞在培养板内增殖到70%~80%的汇合度,更换含有已知浓度RES 的培养基继续培养48 h。
1.2 细胞凋亡的检测 在细胞加入RES 后的24 h 和48 h对pGCs 进行凋亡检测。参考Annexin V-FITC/PI 双染细胞凋亡检测试剂盒说明书对pGCs 凋亡进行检测,贴壁pGCs 用胰酶消化收集,用预冷PBS 洗涤2 次,弃上清。随后加入5 μL Annexin V-FITC 染液,混匀后避光孵育15 min。再加入10 μL PI 染液混匀,再次孵育5 min 后,立即用流式细胞仪检测pGCs 的凋亡。实验重复3 次。
1.3 总RNA 的提取及转录组测序 采用TRIzol 法提取pGCs 的总RNA,利用Nanodrop 检测RNA 浓度及纯度,进一步利用Agilent 2100 bioanalyzer 检测RNA 完整性。经质检合格后委托北京诺禾致源生物科技有限公司进行RNA-seq 文库构建和测序。基于Illumina Hiseq 2500 测序平台构建链特异性文库。经FastQC 软件质控,去除带接头的序列(Reads)、低质量序列及无效碱基后,筛选得到纯净序列(Clean Reads)用于后续分析。使用HISAT2 软件(v2.1.0)将Clean Reads 与猪参考基因组(v11.1)比对,以FPKM(Fragments Per Kilobase of transcript per Million mapped reads)作 为标准归一化各组基因表达量。本文基于RNA-seq 数据鉴定发生在pGCs 中的可变剪接事件,研究参与pGCs凋亡的可变剪接基因。
1.4 可变剪接事件鉴定 通过rMATS 软件(v4.0.2)对9 个样本的可变剪接位点进行分析。rMATS 主要识别外显子跳跃(Skipping Exon,SE)、内含子滞留(Retention Intro,RI)、外显子互斥(Mutually Excl usive Exons,MXE)、5'端可变剪接(Alternative 5'Splici ng Site,A5SS)、3'端可变剪接(Alternative 3'Splicing Site,A3SS)等5 类可变剪接事件。此外对值进行错误发现率(False Discovery Rate,FDR)校正,FDR<0.01 作为判定可变剪接差异显著性的阈值。
1.5 差异基因的GO 注释和KEGG 通路分析 使用GOseq R 软件包对差异基因进行基因本体(Gene Ontology,GO)富集分析,利用KOBAS(v2.0)软件进行京都基因与基因组百科全书(Kyoto Encyclopedia of Gens and Genomes,KEGG)通路富集分析,并基于Edge R 软件绘制差异可变剪接基因的火山图。
1.6 实时荧光定量(qRT-PCR)检测 随机挑选6 个基因(),利用qRT-PCR 检测其相对表达量。使用Primer Premier软件(v5.0)设计引物,以作为内参基因,委托上海生物工程股份有限公司合成所需引物(引物序列见表1)。PCR 反应体系总体积为20 μL:iTaqUniversal SYBRGreen Super Mix 10 μL,上下游引物各0.4 μL(0.1 μg/μL),cDNA 模板1 μL,ddHO 8.2 μL。PCR 反应程序为:95 ℃预变性30 s;95 ℃5 s,61℃35 s(40 个循环);97℃30 s,65℃90 s,97℃10 s。实验在Bio-rad CFX96 Touch实时荧光定量PCR 仪中进行。

表1 实时荧光定量引物序列
1.7 统计分析 使用Excel 2010 整理数据,采用2法计算基因相对表达量,使用SPSS 软件(v20.0)对数据进行统计学分析,方差分析和显著性检验使用-test和one-way ANOVA 来进行分析,并在GraphPad 软件上绘图。
2 结果与分析
2.1 白藜芦醇对猪卵巢颗粒细胞的影响 如图1 所示,RES明显诱导了pGCs 凋亡,并且当RES 浓度达到50 μmol/L时,与对照组相比差异极显著。因此选取50 μmol/L 作为低浓度诱导凋亡处理组(标记为L),100 μmol/L 为高浓度诱导凋亡处理组(标记为H),分别与空白对照组(标记为C)做比较,探讨RES 诱导pGCs 凋亡的差异基因表达。

图1 不同浓度RES 对pGCs 的影响
2.2 数据产出与质控 pGCs 的9 个样本经Illumina PE150系统测序后,平均共产生90 510 724 个Raw Reads,去除带adapter 的Reads、含未知碱基的Reads 以及成对的低质量Reads 后,平均每个样本筛选到89 216 903 个Clean Reads(表2)。与猪的参考基因组(v11.1)平均比对率达96.04%上,测序总体错误率均在0.03%以内,其中高质量Clean Reads 的Q30 占比在94%以上,说明测序数据真实可靠。

表2 转录组测序数据及比对结果
2.3 基因表达趋势分析 对3 组pGCs 中基因的表达模式进行聚类(<0.001),发现3 组pGCs 呈现8 种表达模式(图2),分为8 个模块(分别是Profile 0、1、2、3、4、5、6、7)。有676 个基因表达与RES 浓度呈正相关(Profile 7),329 个基因表达量与浓度呈负相关(Profile 0),391 个基因的表达量在低浓度处理组中存在拐点(Profile 6)。对Profile 6 中的基因进行GO 功能富集和KEGG 通路分析(<0.05),发现Profile 6 中的基因主要富集在细胞内,与DNA 复制、谷胱甘肽运输、氧化还原酶活性等条目相关,参与卵巢类固醇生成、甾醇激素合成、谷胱甘肽代谢等信号通路(图3)。
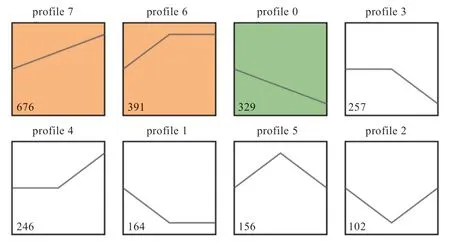
图2 基因表达趋势聚类
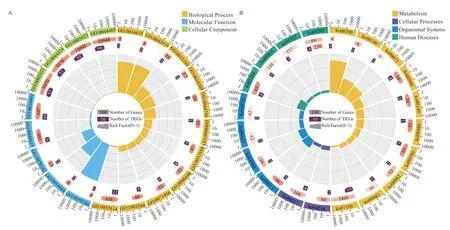
图3 模块6 中基因的GO 和KEGG 富集
2.4 可变剪接事件鉴定及差异基因筛选 将9 个样本的转录组数据与猪参考基因组比对后,通过rMATS 软件对基因组比对结果进行可变剪接分析,在低浓度组与对照组(L vs.C)的比较中,共鉴定出34 186 个可变剪接事件。在高浓度组与对照组(H vs.C)的比较中,共鉴定出34 536 个可变剪接事件(表3)。在不同的可变剪接事件类型中,以SE 类型的可变剪接为主。以<0.05、FDR<0.01 作为差异显著的判断标准,在L vs.C 组中筛选到198 个基因参与了236 个可变剪接事件,在H vs.C 组中,有603 个基因发生了779 个可变剪接事件。

表3 可变剪接事件统计
不同浓度RES 诱导pGCs 凋亡的比较中,SE 类型的可变剪接数量均最多。在L vs.C、H vs.C 组中,pGCs 发生SE、RI、MXE 等类型可变剪接事件的比例基本无明显变化(图4),分别为75%、7%、9%,以及78%、8%、7%。在L vs.C 组中,SE 事件有178 个,在H vs.C 组中,SE 事件则增加到605 个。A3SS、A5SS类型分别由8%下降到4%,由1%上升到3%。由此可见,参与不同浓度RES 诱导pGCs 凋亡的可变剪接基因具有特异性,可变剪接事件的数量在不同细胞状态下存在较大差异。

图4 各类型可变剪接事件的比例
利用DESeq 软件对3 组pGCs 的可变剪接基因进行差异分析,按照差异表达倍数log2|FC|>1、显著性<0.05 为标准筛选差异可变剪接基因,结果如图5 所示,L vs.C 组有6 个差异可变剪接基因(3 个基因显著上调,3 个基因显著下调)。H vs.C 组有35 个差异可变剪接基因(15 个基因显著上调,20 个基因显著下调)。
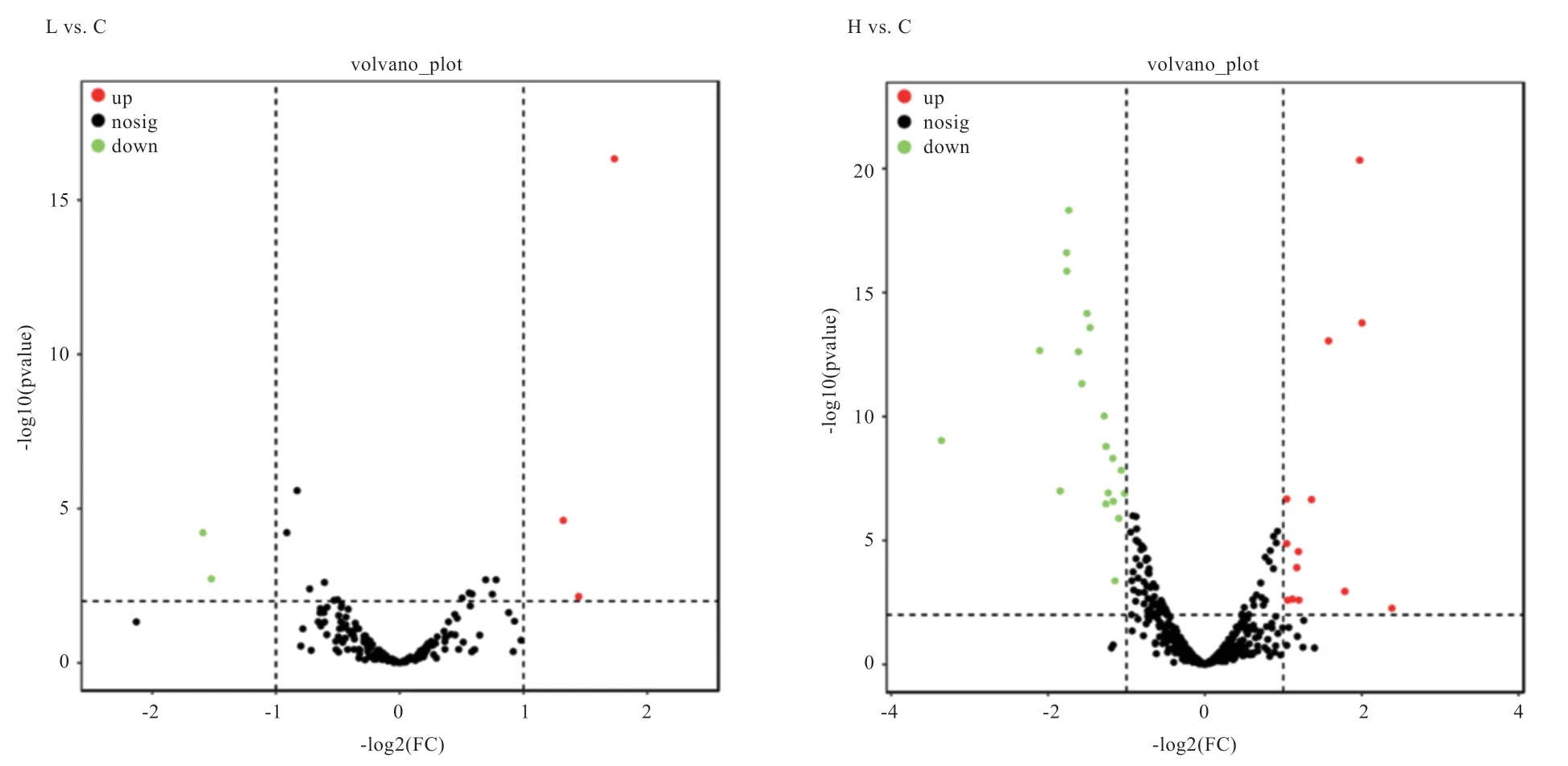
图5 差异可变剪接基因火山图
2.5 差异可变剪接基因GO 富集分析 为了解发生可变剪接基因的功能,对发生差异可变剪接的基因进行了GO 注释和功能分析。L vs.C 组中6 个发生差异可变剪接的基因被富集到208 个生物学过程(Biological Process,BP)条目、27 个分子功能(Molecular Function,MF)条目和23 个细胞组分(Cellular Components,CC)条目,推测发生差异可变剪接的基因可能广泛参与生物学过程。针对富集到的结果,选取具有统计学意义(<0.05)的10 个GO 条目,发现差异可变剪接基因主要富集在谷氨酰胺合成与代谢、氨基酸合成、细胞凋亡、氧化还原等生物学过程(表4)。
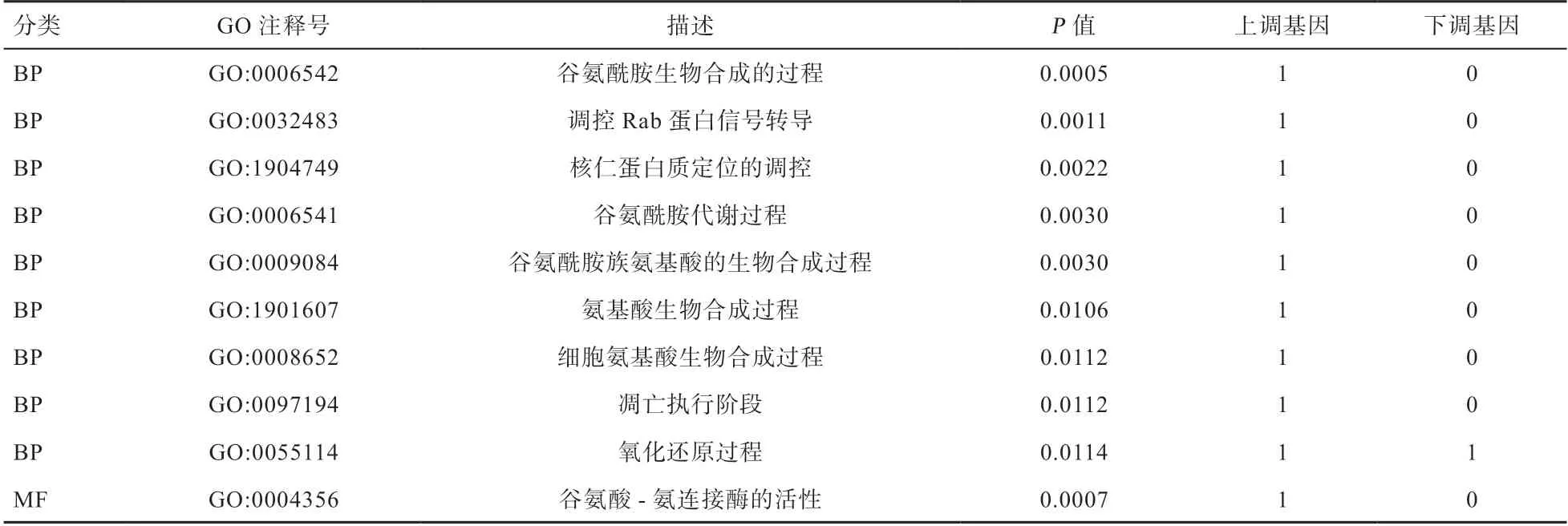
表4 L vs.C 组合差异可变剪接基因GO 分析
H vs.C 组中发生差异可变剪接的基因有35 种,被富集到831 个生物学过程条目、167 个分子功能条目和134 个细胞组分条目。其中显著富集的10 个GO 条目(表5),主要与脂多糖结合、细胞对缺氧反应的正向调节、吞噬小泡中的蛋白质加工、低氧诱导因子-1 信号通路正调控、吞噬囊泡中蛋白质加工的调控、谷氨酰胺生物合成、磷脂酰胆碱分解代谢等生物过程相关,在分子功能上发挥脂多糖运输功能。

表5 H vs.C 组合差异可变剪接基因GO 分析
2.6 差异可变剪接基因KEGG 通路富集分析 如图6 所示,L vs.C 组中差异可变剪接基因主要富集于代谢途径、卵巢类固醇生成、氨基酸生成及代谢等通路。H vs.C 组发生差异可变剪接的基因被富集到卵巢类固醇生成、-氨基丁酸能突触、谷氨酸能突触、精氨酸及叶酸生物合成、维生素消化吸收等通路。
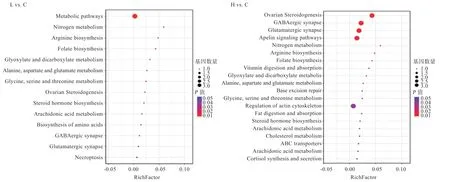
图6 差异可变剪接基因的KEGG 富集分析
2.7 白藜芦醇诱导猪卵巢颗粒细胞凋亡关键可变剪接基因的筛选 基于所筛选2 个组合的差异可变剪接基因,对其差异表达进行了分析(图7),发现有4 个基因在L vs.C 及H vs.C 中存在共同表达,分别是醛酮还原酶家族1 成员C3 基因(Aldo-keto Reductase Family 1 Member C1,)、谷氨酸-氨连接酶基因(Glutamate-Ammonia Ligase,)、肌氨酸脱氢酶基因(Sarcosine Dehydrogenase,)以及MAP 激酶激活死亡结构域基因(MAP Kinase Activating Death Domain,)。结合上文的GO功能富集和KEGG 通路分析,发现及基因被显著富集到80 个GO 条目和20 条KEGG 信号通路(图8),广泛参与了与细胞生长发育、信号转导、细胞凋亡及代谢过程。其中,基因被64 个GO 条目及13 个信号通路富集,表明基因对细胞生命活动影响程度更高。以上分析结果表明这4 个发生差异可变剪接的基因,可能在RES 诱导pGCs 凋亡中发挥作用。

图7 差异可变剪接基因的维恩图
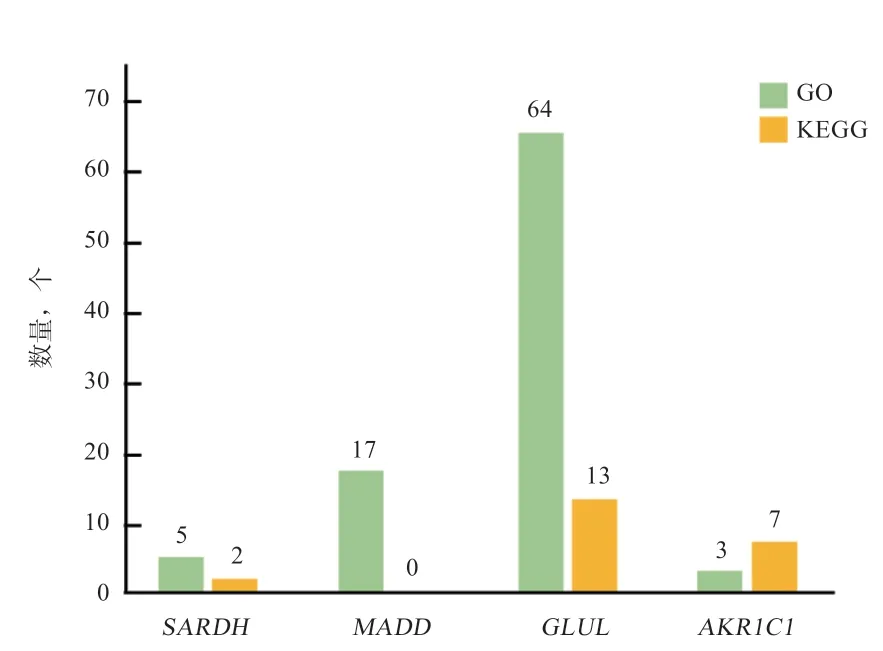
图8 共表达差异基因富集到GO/KEGG 的数目
2.8 qRT-PCR 验证测序可靠性 为了验证测序结果,随机选择了6 个基因进行qRT-PCR 验证(图9)。结果显示,差异表达基因在各组中的表达趋势与转录组测序集中的结果一致,证实了测序数据的可靠性。
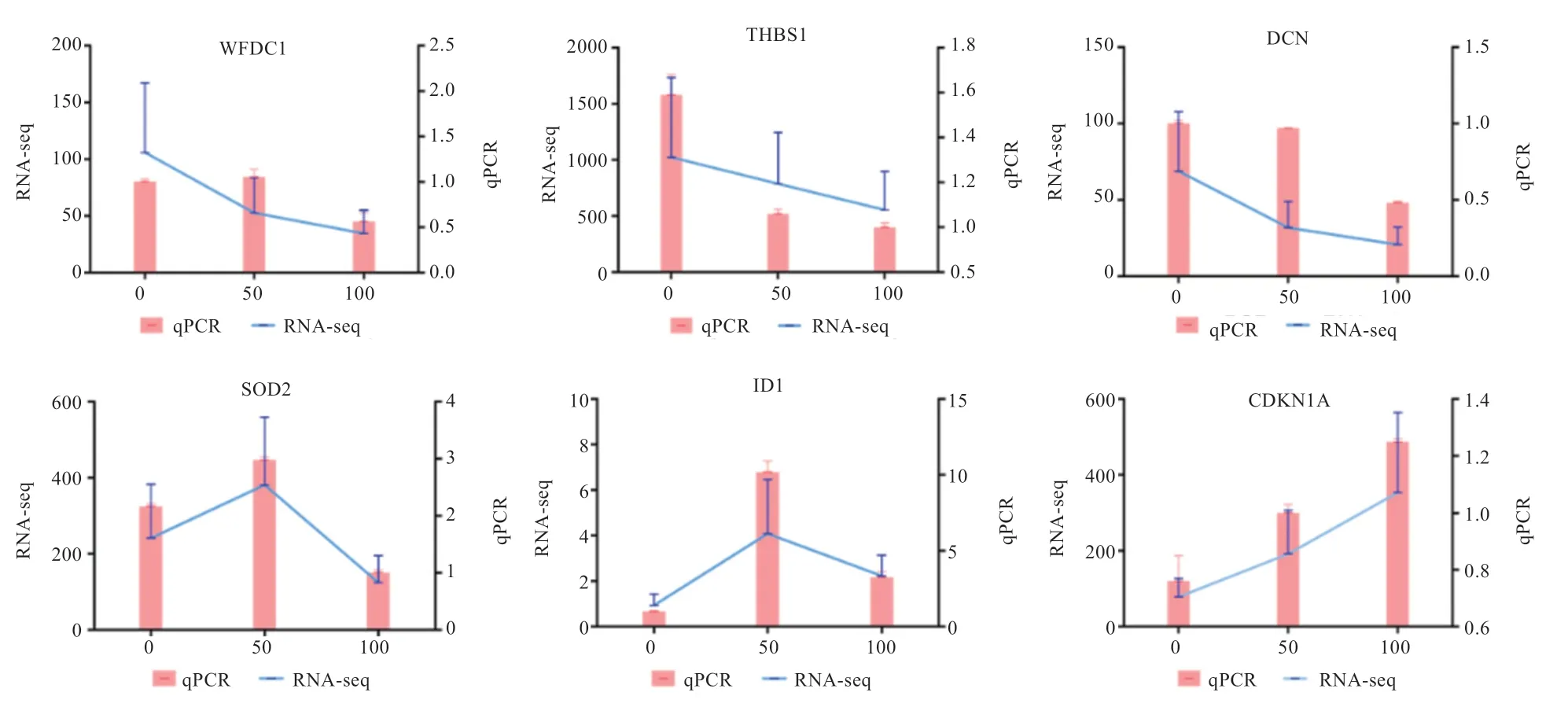
图9 差异可变剪接基因的qRT-PCR 和RNA-seq 结果比对图
3 讨 论
一个基因可以在mRNA 前体中以不同的剪接方式形成多种mRNA 剪接异构体,经过翻译,最终在蛋白水平层面表现出不同的功能和结构特异性,被认为是形成基因组和蛋白组多样性的一种有效机制。目前研究表明,基因可通过可变剪接等调控机制,对细胞增殖分化、坏死及凋亡等生命过程产生重要影响。作为生命科学领域研究的重要技术,RNA-seq 能高效产生基因组序列、准确定量出基因表达量,结合生信分析手段可以实现在全基因组水平分析各种细胞不同生理状态下的可变剪接事件。
本研究证实了50 μmol/L 和100 μmol/L RES 极显著的促进了pGCs 凋亡,并通过RNA-seq 和生信分析,对RES 诱导pGCs 凋亡相关基因的差异表达和可变剪接进行研究,在50 μmol/L RES 处理组发现了6 个差异可变剪接基因,100 μmol/L RES 处理组有35 个差异基因发生了可变剪接。此外大多数发生可变剪接的基因主要发生了外显子跳跃类型的剪接,3'端可变剪接和5'端可变剪接的类型较少,表明可变剪接对于调控这些基因的表达具有重要作用。
通过对发生差异可变剪接的基因进行GO 功能富集分析发现,大多数基因富集在氨基酸合成及代谢、细胞凋亡、氧化还原及细胞缺氧反应调节、蛋白质加工调控等与细胞生命活动相关的GO 条目中,这表明可变剪接通过调控差异基因的蛋白质产物而在RES 诱导pGCs凋亡的过程中发挥重要作用。此外,不同浓度RES 处理pGCs,存在着不同的特异性可变剪接基因,证实了可变剪接在不同细胞状态下对蛋白质产物结构及功能起调控作用。在KEGG 通路富集分析中,差异可变剪接基因主要富集到的信号通路(卵巢类固醇生成信号通路、代谢途径信号通路的信号通路、氨基酸代谢相关信号通路等)均与激素分泌、细胞凋亡、有机物的吸收相关,这与前人研究一致。研究表明,卵巢类固醇生成信号通路受损会抑制哺乳动物卵泡排卵、导致颗粒细胞功能异常,代谢途径信号通路广泛发生在机体,参与生命活动的调节。
为进一步筛选可能参与RES 诱导pGCs 凋亡的可变剪接基因,结合差异可变剪接基因的GO 和KEGG 富集分析结果,发现及基因广泛参与了细胞生命活动相关的GO 条目和信号通路,可能是参与RES 诱导pGCs 凋亡的关键基因。其中,基因家族成员被证实参与了多囊卵巢综合征和卵巢肿瘤的发生,与妊娠晚期卵泡黄体退化相关。基因与谷氨酰胺合成相关,已有研究指出该基因可以参与细胞坏死性凋亡进程和有丝分裂。基因可以与基因互作,在肿瘤细胞侵袭、迁移及细胞凋亡过程中发挥重要作用。而基因可以通过抑制细胞凋亡促进肺腺癌细胞生长,与卵巢癌细胞死亡相关。前人研究结果提示,本文所筛选的4 个差异可变剪接基因均与细胞增殖及凋亡相关,这些可变剪接基因可能在RES 诱导pGCs凋亡过程中发挥特定功能。
4 结 论
本研究基于RNA-seq 和生物信息学分析,鉴别并筛选了白藜芦醇诱导猪卵巢颗粒细胞凋亡过程中发生的差异可变剪接事件,发现大多数基因发生了外显子跳跃形式的可变剪接。通过GO 和KEGG 分析,进一步筛选得到了及等可能参与白藜芦醇诱导猪卵巢颗粒细胞凋亡的差异可变剪接基因。本研究为猪卵巢颗粒细胞凋亡机制的研究提供了一定的数据支持,也为进一步研究白藜芦醇对卵巢发育的影响提供了科学的理论基础。
