三峡水库支流浮游植物演替内外驱动力辨析
2022-08-10陈佳俊张佳磊
陈佳俊,张佳磊
(湖北工业大学土木建筑与环境学院/河湖生态修复与藻类利用湖北省重点实验室,武汉 430068)
三峡蓄水成库后,因改变三峡水库的水动力条件,从而影响三峡水库支流的水温结构、光照条件和营养盐迁移转化规律[1-4],导致水体中浮游植物群落特征发生一系列的改变,由河流型硅甲藻向湖泊型蓝绿藻水华演替[5],而多数蓝藻在代谢过程中分泌的藻毒素破坏了生态环境,对饮用水源安全产生严重影响,引起国内外的广泛关注[6-8]。
浮游植物是水生态系统中的初级生产者,其群落结构能反映水质状况的好坏[9]。浮游植物群落的变化受到水温、光照、营养盐以及水动力条件的影响[10],水温对藻类生长速率有一定影响,不同水温下浮游植物的种类存在显著差异[11]。不同水文期的水体光学特性存在显著差异,真光层深度以及光合有效辐射的变化影响浮游植物的生物量[12]。水体变缓,颗粒物携带营养盐沉积效应加强,营养盐在支流库湾中的滞留,浮游植物生物量升高[13]。河流、水库等流动性水体,其浮游植物生境的主要参量是水动力条件,已有研究表明水华暴发与水动力条件呈显著相关[14-17]。综上所述,三峡水库的浮游植物演替是外部环境条件改变所引起的。
但是从生态学角度来看,浮游植物的演替过程是内部竞争与外部环境条件共同驱动下发生的。目前多关注外部因素,而对内部因素效果关注较少,根本原因是内部因素不好分辨。趋同性的核心是指浮游植物群落演替由环境条件所决定;在相同的环境条件下,浮游植物群落将向同一方向进行演替。不同环境条件下,浮游植物群落将向不同的方向进行演替。趋异性是指浮游植物群落的演替由群落的初始结构所决定;在相同的环境条件下,由于初期群落结构的不同,也将向不同的方向演替[18]。为明确三峡库区浮游植物演替的内外驱动力因素,本研究基于浮游植物群落构建的趋同性和趋异性理论,开展了温度、光照对浮游植物群落影响的室内控制试验,以期探究影响河道型水库浮游植物群落生物量与多样性的关键因子。
1 材料与方法
1.1 藻种来源与试验装置
藻种来源为香溪河河水。光照系统采用日光灯组连续提供,确保光照强度分布均匀且稳定在(2 000±150)lx;水温用可调式绝缘电热棒控制,精确度为±1 ℃。试验装置如图1所示,将9个长50 cm、宽38 cm、高30 cm 的白色水框平行布置,放置于自然条件下。在每个水框中均匀放入2 个微型水泵,用以扰动水体,避免水体中的藻类自然沉降。微型水泵通过硅胶软管与空气压缩机连接;进气前端设有孔径为0.2 μm 的空气过滤器用以除尘除菌,另设置空气开关阀用以控制进气量的大小。
1.2 试验方法
设有3 个试验组,分别为空白对照组、温度组和光照组。每组设3 个平行,共9 个白色水框。空白对照组为自然条件下的光照及温度;温度组为自然条件下的光照,温度设定为26 ℃;光照组为自然条件下的温度,光照设定为2 000 lx。
监测时段为2016 年5 月4—26 日,采样频率为前14 d 每天采样1 次,之后每3 d 采样1 次,共17 次。为了解试验前各水框的初始状况,试验初始测定每个水框中藻类的初始群落结构,以及每个水框中TN、TP 浓度,之后每3 d 测定1 次,满足藻类生长所需的氮、磷营养盐阀值(TN>0.2 mg/L,TP>0.02 mg/L)。24 h 不间断曝气,促进营养物质循环,尽量避免藻类沉降。
1.3 测定指标及方法
试验开始后,每天早上09:00 左右取样。光照强度和水体温度分别用TES-1339 照度计(台湾泰仕)和玻棒煤油温度计(河北光华)测定。TN、TP 浓度利用UV-1700 紫外分光光度计(上海美析)进行测定[19],藻类群落结构利用AlgacountS300 藻类智能鉴定计数仪(杭州迅数)进行鉴定和计数。藻类种类根据文献[20,21]进行鉴定,藻细胞计数和生物量测算根据文献[22]。藻种统计至属即可完全反映藻类群落特征及演替过程,因此本研究藻类鉴定数据统计到属[23]。
1.4 数据处理
1.4.1 生物量

式中,N为1 L 水样中浮游生物的数量,ind/L;Cs为框面积,mm2;Fs为视野面积,mm2;Fn为每片计数过的视野数;V为1 L 水样经浓缩后的体积,mL;V0为计数框容积,mL;Pn为计数的浮游植物个数。
按上述方法进行采样、稀释、计数。Cs为400 mm2,Fs为0.156 549,Fn为100,V为1 000 mL,V0为0.10 mL。因此只要计数方法确定,就可以得到N。
1.4.2 演替速率 通常在生境稳定条件下,群落演替的结果是达到顶极群落(Climax)。演替速率是群落结构改变潜势的客观测度。Lewis(1978)修正了Jassby and Goldman(1974)建立的群落演替速率计算方法,并在藻类群落生态研究中广为采用,其计算公式如下。
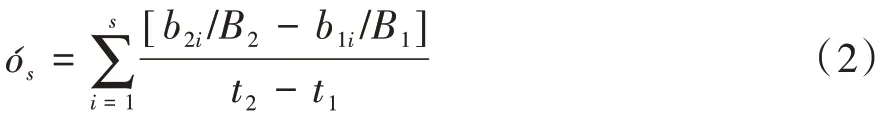
式中,B2、B1分别为群落演替过程中2 个时间状态(t2、t1)的藻类生物量,而b2i、b1i则为上述相应状态下群落中第i个藻种的生物量。
1.4.3 多样性指数 常用的多样性指数[23-25]有Mar⁃galef 指 数、Shannon-Winener 指 数 和Jaccard 指 数。Margalef 指数主要反映群落物种的丰富度,浮游植物种类愈多,S越大。Shannon-Winener 指数反映群落结构的复杂程度,是衡量种类数和均匀度的综合指标。Jaccard 指数反映样点间浮游植物群落结构的相似性,J愈大,表明样点间的浮游群落结构愈相似。
1)Margalef丰富度指数(S)。

2)Shannon-Winener 指数(H)。

3)Jaccard 种类相似性指数(J)。

式中,S为群落中物种数,Pi=Ni/N;Ni为第i种藻的个数,N为观察到的总个数。J为相似性指数,a为样点A 的种类数;b为样点B 的种类数;c为样点A 和B 共同的种类数。当0<J<0.25 时,样点A 和B 的浮游植物群落结构达到极不相似水平;当0.25≤J<0.50 时,样点A 和B 的浮游植物群落结构达到中等不相似水平;当0.50≤J<0.75 时,样点A 和B 的浮游植物群落结构达到中等相似水平;当0.75≤J<1.00时,样点A 和B 的浮游植物群落结构达到极相似水平[26]。
1.4.4 数据分析 数据分析利用SPSS 17.0 统计软件,采取单因素ANOVA 均值分析方法。P<0.05 为显著,P<0.01 为极显著。
2 结果与分析
2.1 不同环境条件下藻类群落演变特征
2.1.1 藻类细胞生物量 在试验期间,对照组、温度组和光照组的藻类细胞密度存在显著差异(ANO⁃VA,P<0.05)。对照组、温度组和光照组时间变化趋势一致,均呈先上升后下降趋势,但是其达到峰值的时间略有差别,分别为5 月13 日、5 月10 日和5 月9日。对照组、温度组和光照组在试验期间的生物量平 均 值 为(105.8±82.5)×105、(109.8±90.1)×105、(402.5±289.6)×105ind/L,光照组藻类细胞生物量显著大于对照组和温度组(图2)。
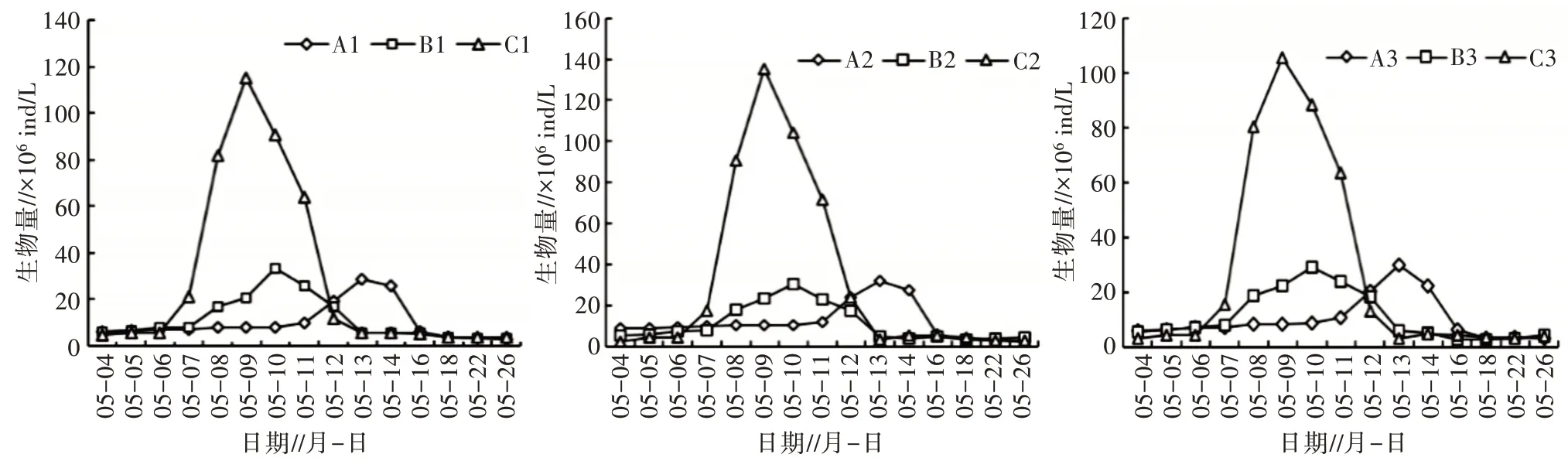
图2 不同环境条件藻类细胞生物量变化
2.1.2 藻类的群落结构特征
1)不同环境条件藻类群落特征。根据形态鉴定,试验期间共鉴定出浮游植物27 种,隶属于4门[20]。其中种类最多的是绿藻门(Chlorophyta)(12种),其次是硅藻门(Bacillariophyta)(9 种)、蓝藻门(Cyanophyta)(4 种)和裸藻门(Euglenophyta)(2 种)。
不同的光照和温度条件对浮游植物群落的演替有明显的影响,试验组的优势种存在一定的差异性。初始培养期,光照组、温度组、对照组的浮游植物群落结构均由硅藻、蓝藻、绿藻和裸藻组成。从初始培养到培养稳定后优势藻类的种类明显减少,其中,小环藻属(Cyclotella)是光照组最主要的优势种,温度组主要优势种是衣藻属(Chlamydomonas)和小球藻属(Chlorella),对照组的优势藻类为微囊藻(Micro⁃cystis)和色球藻(Chroococcus)(表1)。

表1 各试验组的藻类演替规律
2)不同环境条件藻类演替速率比较。演替速率是指在某一时间段内藻类群落的演替速率。根据表2 可以看出,光照组的藻类演替速率最快,温度组次之,对照组生长最为缓慢。所以光照和温度条件对藻类群落结构的改变有一定影响。
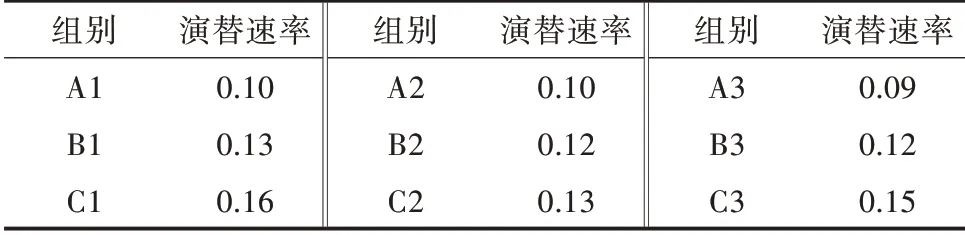
表2 不同环境条件下的演替速率
3)藻类群落结构相似性比较。由图3 可以看出,试验期间不同环境条件下试验组的浮游植物相似性指数在0.27~0.67。试验初期,各试验组的浮游植物相似性指数在0.55~0.60,其中对照组与光照组的相似性指数在0.30~0.43,平均值为0.48±0.03,为中等不相似水平;对照组与温度组、光照组与温度组群落结构平均值分别为0.54±0.04、0.55±0.05,达到中等相似水平。5 月26 日浮游植物到达演替后期,对照组与温度组、光照组,温度组与光照组的浮游植物相似性指数介于0.36~0.45,其种类相似性水平平均值分别为0.39±0.03、0.39±0.01、0.40±0.04,为中等不相似水平。
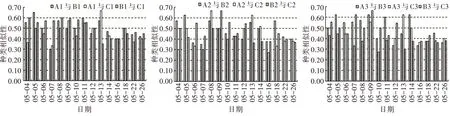
图3 不同环境条件Jaccard 种类相似性指数变化
4)藻类的物种丰富度及多样性指数。光照组与温度组具有极显著差异(P<0.01),光照组与对照组呈显著差异(P<0.05),温度组与对照组差异不显著(P>0.05)。从藻类的Shannon-Winener 多样性指数分析结果(图4)可以看出,各试验组的多样性指数呈波动下降后上升趋势;对照组、温度组、光照组的平均值分别为0.97±0.50、0.91±0.54、1.27±0.36,藻类群落多样性大小为C>A>B。

图4 不同环境条件Shannon-Winener 多样性指数变化
从浮游植物的Margalef 丰富度指数分析结果(图5)可以看出,光照组、温度组和对照组的Mar⁃galef 丰富度指数分别为0.13~0.72、0.13~0.51、0.12~0.62,其平均值分别为0.39±0.12、0.28±0.09、0.29±0.09,光照组物种丰富度均在5 月12 日达到最大,在5 月8—26 日维持稳定,不同试验组间的物种丰富度差异显著(P<0.05)。

图5 不同环境条件Margalef丰富度指数变化
2.2 相同环境条件下藻类群落演变特征
2.2.1 藻类细胞生物量 3 个平行组之间的藻类细胞密度不存在显著性差异(P>0.05),由图6 可以看出,相同环境条件下的平行组整体变化趋势一致,随时间的推进均呈先上升后下降的趋势。
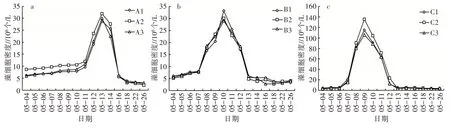
图6 相同环境条件藻类细胞生物量变化
2.2.2 藻类的群落结构特征
1)藻类群落鉴定结果及演替速率。分别对相同条件各平行组藻类群落形态鉴定(表3),结果表明,稳定期光照组的3 个平行组优势种(优势藻以超过总生物量10%为评定标准)均为小环藻属(56.35%,58.24%,55.69%)。温度组的3 个平行组优势种均为衣 藻 属(58.22%,56.25%,60.36%)和 小 球 藻 属(36.17%,40.05%,35.14%)。由于对照组3 个平行处于自然条件下,环境因子干扰,其对浮游植物演替不显著,导致浮游植物的内部竞争更为明显[18]。因此,对照组的3 个平行组优势种不一致,A1与A3均为微囊藻属(40.67%,38.50%),色球藻属(20.67%,19.16%);A2为 微 囊 藻 属(42.58%)、小 球 藻 属(30.61%)。
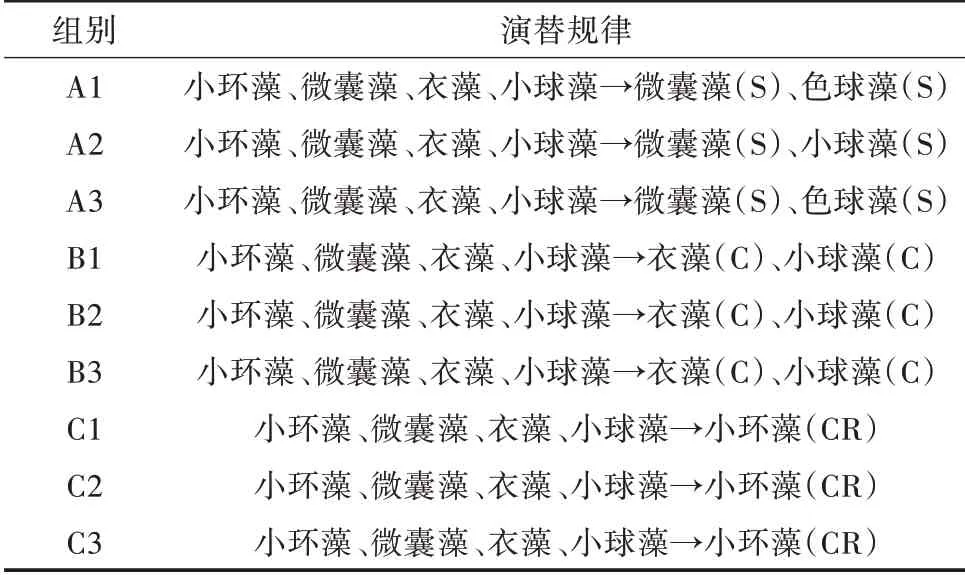
表3 相同条件各平行试验组藻类演替规律比较
2)相同环境条件下藻类群落相似性比较。由相同环境条件Jaccard 种类相似性指数变化(图7)可以看出,对照组、温度组和光照组的初始浮游植物相似性指数分别在0.56~0.71、0.36~0.50、0.43~0.57,其平均值分别为0.62±0.08、0.68±0.16、0.57±0.12。在浮游植物达到峰值时,浮游植物相似性指数介于0.50~0.83,浮游植物群落达到中等相似水平。5 月26 日浮游植物演替后期,各试验组的浮游植物相似性指数在0.67~1.00,对照组、温度组和光照组的种类相似性平均值分别为0.78±0.05、0.91±0.08、0.88±0.06,达到中等以上相似水平。
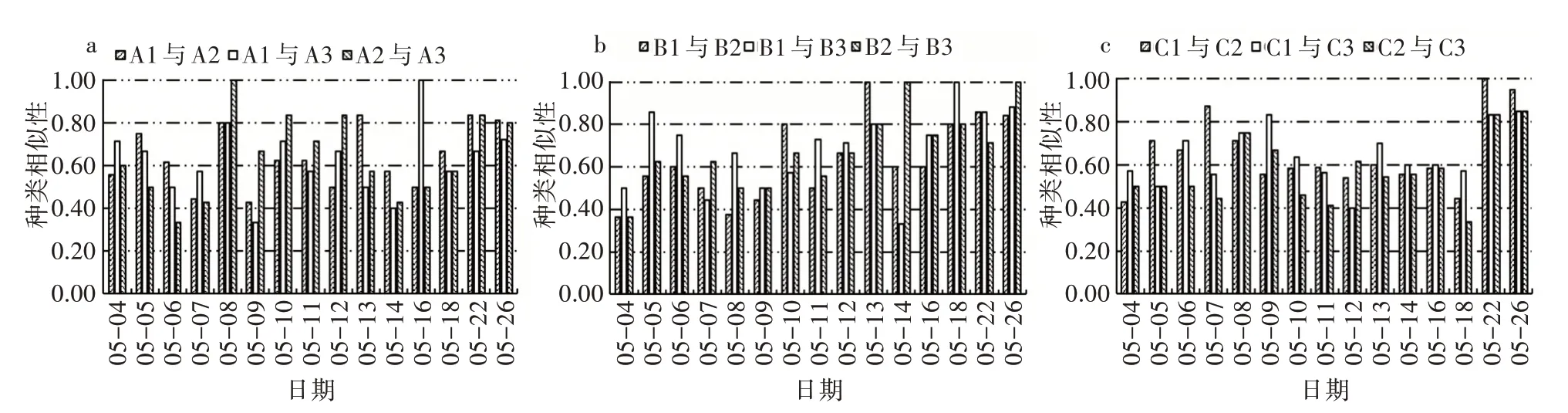
图7 相同环境条件Jaccard 种类相似性指数变化
3)藻类的物种丰富度及多样性指数。相同环境下,平行组的藻类多样性指数无显著差异(ANOVA,P>0.05)。相同环境下各水框中浮游植物群落的Shannon-Winener 指数与Marglef 丰富度指数(图8、图9)随时间的变化规律相似。各平行试验组藻类多样性指数呈相同的变化趋势,总体上表现为先减小后增大的变化规律。光照组、温度组和对照组分别为0.50~2.07、0.09~1.83、0.23~2.22,其中光照组在5 月8 日多样性指数最小,在5 月18—26 日趋于稳定;温度组在5 月10 日多样性指数最小,在5 月16—26 日浮游植物多样性指数趋于稳定;对照组A1与A2的多样性指数呈先增大后减小再增大的变化规律,A3表现为先减小后增大的变化趋势。
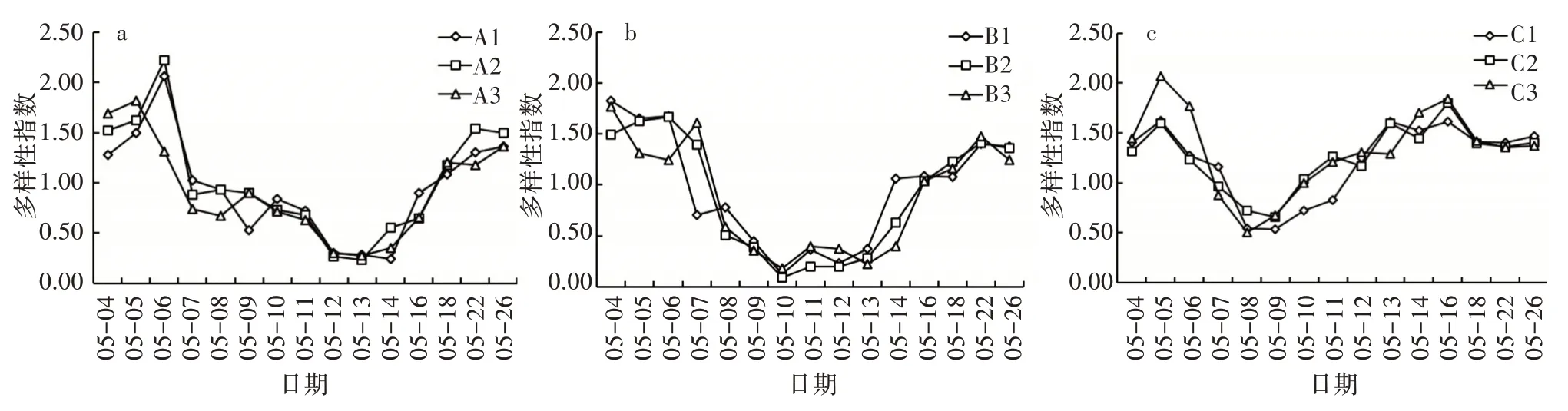
图8 相同条件下Shannon-Winener 多样性指数变化

图9 相同条件下Margalef丰富度变化
3 讨论
3.1 环境因子对浮游植物群落结构的影响
试验期间,各试验组水体处于富营养化水平(TN>0.2 mg/L,TP>0.02 mg/L)。鉴于浮游植物自身含有营养盐,氮、磷、营养盐含量与浮游植物生物量的变化有很强的自相关性,并且各试验组水体为富营养化水体,因此营养盐对各试验组浮游植物群落结构变化的影响不是主要因素,驱动浮游植物群落改变的主要外界因素是温度和光照[27]。
浮游植物群落演替表征为生境变化下,不同浮游植物随时间的变化呈相互取代,是自然界发生且可辨识的[28]。在富营养化水体中,浮游植物生物量的增加将导致其群落多样性的下降[29,30]。浮游植物在适宜的生境下往往具有较高的生长速率,从而生物量能在充足的时间内得到积累而形成优势;不适浮游植物的生境将因其生长速率受到抑制,在此生境下不具备优势,而使其被其他浮游植物所取代[10]。在不同环境下3 个试验组的浮游植物生物量在不同时间节点达到峰值,并且其藻类生物量大小相差较大。光照组最先达到峰值,温度组次之,对照组最后达到峰值。在浮游植物生物量达到最大时,光照组的优势种演替为小环藻,温度组的优势种演替为衣藻,对照组的优势种演替为微囊藻。在试验后期,光照组的优势种均为小环藻;温度组的优势种均为衣藻;对照组光热条件较差,浮游植物的生长受到光热条件限制较为明显,具有低光强的微囊藻、小球藻以及色球藻在该生境条件下具备生存优势,在试验期间演替为优势种。但是当外界环境因子对浮游植物演替不显著时,浮游植物的内部竞争更为明显[18]。导致其3 个平行的优势种不一致,A1与A3组为微囊藻、色球藻;A2组为微囊藻、小球藻。不同环境条件下Jaccard 种类多样性指数达到中等不相似水平,Margelef 丰富度指数与Shannon-Winener 多样性指数随时间的变化呈显著差异(ANOVA,P<0.05)。由此可见,不同环境条件下浮游植物群落演替表现为趋异性。
在相同环境下,组间浮游植物演替具有相似的变化趋势,浮游植物群落的构建表现为趋同性。3个试验组的浮游植物生物量均在同一天达到峰值,其生物量较接近。Jaccard 种类相似性达到中等以上相似,Margelef 丰富度指数与Shannon-Winener 多样性指数(ANOVA,P>0.05)随时间的变化无显著差异。根据以上比较可以看出,外界的环境因子对浮游植物群落演替的驱动力较大。
3.2 基于生态学理论辨析三峡水库浮游植物群落结构演替驱动力因素
水生环境浮游植物生物量和群落结构的改变取决于物理(水文,水动力条件)、化学(营养盐)以及生物因素(浮游动物捕食作用,浮游植物内部竞争)的相互作用[10,31-33]。不同湖库的浮游植物群落结构演替因素有所区别,有研究表明,水温、光辐射强度以及营养盐浓度是浅水湖泊产生群落结构演替的主导因子[34,35]。对于深水型水库的调查结果显示,水体下光热结构(光照和水体层化模式)、水体滞留时间、外源营养盐输入等环境因子的改变导致浮游植物群落结构的演替[36,37]。三峡水库的特殊水动力学造成了支流水生生境的改变,有研究表明,水位波动、水温、光热结构和水体稳定系数是改变三峡水库支流浮游植物群落结构的关键因素[38-41]。三峡水库支流的浮游植物群落结构演替的驱动力主要是外界的环境因子。
根据“生态学理论”,Reynolds[10]认为浮游植物生境特征与物质、能量的供给相关,物质供给主要为浮游植物对水体营养盐的利用程度,能量供给则为温度和光照2 个方面。在稳定的环境中,水体中的浮游植物群落会相互竞争,导致群落多样性下降,生物量达到最大,演替速率达到最低。对于外界环境条件影响较大的光照组和温度组生物量相继达到峰值,此时的群落多样性指数达到最低值。
4 结论
不同外界环境条件下,光照组浮游植物群落结构演替速率最快,相比于温度组和对照组,其生物量具有明显的升高趋势,光照条件对浮游植物群落演替的驱动更为显著。 不同试验组的优势种呈现出一定的差异性。浮游植物在生物量达到峰值时,光照组的群落结构以小环藻占优,温度组的优势种为衣藻和小球藻,对照组的优势种为微囊藻和色球藻,在外界环境因子影响较为显著时,浮游植物群落结构向同一方向演替。
