新疆和西藏棕鳟群体的遗传多样性分析
2022-08-09吴昊天李亚琳陈晓雯王成辉
吴昊天,李亚琳,王 军,陈晓雯,陆 颖*,王成辉*
(1水产种质资源发掘与利用教育部重点实验室(上海海洋大学),上海 201306;2农业农村部淡水水产种质资源重点实验室(上海海洋大学),上海 201306)
0 引言
【研究意义】棕鳟(Salmo trutta)属于鲑形目(Salmoniformes)鲑科(Salmonidae)鲑亚科(Salmononae),原产于欧洲、非洲北部和中亚一些地区(胡江伟和阿地力,2018)。棕鳟是适应高海拔的冷水鱼类,其肉质鲜美,具有重要的经济价值。新疆乌鲁木齐达阪城东沟冷水鱼实验场和西藏亚东县亚东鲑鱼繁育基地是我国目前仅有的2个棕鳟人工养殖基地(分别简称为“新疆棕鳟”和“西藏棕鳟”),棕鳟人工养殖业的发展对当地居民实现脱贫致富具有重要作用。然而,新疆棕鳟与西藏棕鳟的养殖方式不同,气候环境也存在较大差异,因此,对两地棕鳟进行群体遗传学研究,有助于从基因组水平深入了解这些群体的遗传结构,进而指导物种保护和育种研究。【前人研究进展】新疆棕鳟和西藏棕鳟的来源可追溯到19世纪末20世纪初。1866年,英国人将棕鳟引入青藏高原的亚东河流域养殖,在我国被称为亚东鲑鱼(胡江伟和阿地力,2018;高擘为等,2020)。与青藏高原的其他鱼类相比(陈毅峰,2002),亚东鲑鱼的生长速度和性成熟更快,使得亚东鲑鱼在当地迅速建立了种群(豪富华等,2007)。1900年左右,来自英国的棕鳟被引入克什米尔(Sahoo et al.,2016),随后从克什米尔传播到巴基斯坦。自2011年起,新疆地区开始从巴基斯坦引入棕鳟养殖,后逐步推广到新疆多地(胡江伟和阿地力,2018)。棕鳟在国内的发源历史及其养殖群体的遗传多样性现状目前还极少有人关注。豪富华(2006)、豪富华等(2006)曾对亚东鲑鱼及引种自丹麦的棕鳟进行生物学和遗传多样性分析,发现亚东鲑鱼相较于引种自丹麦的棕鳟有着包括生长速度、胚胎发育和繁殖特点上的差异,但其对线粒体基因组分子标记的研究中发现亚东鲑鱼与丹麦棕鳟的序列无任何差异,但通过11个多态性微卫星标记的研究却发现亚东鲑鱼的遗传多样性高于丹麦棕鳟。孟玮等(2017)基于ITS1基因序列对西藏棕鳟和引自巴基斯坦的新疆棕鳟进行群体遗传分析,认为新疆棕鳟的遗传多样性高于西藏棕鳟。目前,国际上主要使用微卫星分子标记(Eszterbauer et al.,2015;Prodöhl et al.,2019)和利用限制性酶切位点相关DNA测序(Restriction-site associated DNA,RAD-seq)得到的单核苷酸多态性(SNP)标记(Leit‐wein et al.,2016)来研究棕鳟群体的遗传多样性。Lemopoulos等(2019)比较使用RADseq得到的SNP标记与使用微卫星标记估计棕鳟遗传多样性和杂合水平的效果,发现使用SNP标记在对多位点的估计上更具优势。尽管棕鳟染色体水平的基因组序列已发表(Hansen et al.,2021),但通过基因组重测序对棕鳟群体进行遗传多样性研究的报道还较少。截至2022年1月,仅Saha等(2022)参考棕鳟染色体水平基因组,基于重测序数据,在瑞典Bunnersjöarna湖区的2个共存棕鳟群体中找到2个群体间存在生殖隔离的证据。【本研究切入点】棕鳟具有40条染色体,曾发生过染色体加倍,高度的遗传变异性以致使用少数的基因或分子标记难以正确反映棕鳟遗传多样性的实际水平。2019年,英国Sanger研究所完成了25个物种的基因组计划,其中包括棕鳟的高质量基因组序列(Hansen et al.,2021),这使得在基因组水平解析棕鳟的遗传多态性成为可能。【拟解决的关键问题】采用第二代高通量测序技术对来自新疆达坂城和西藏亚东的2个棕鳟群体进行基因组重测序,从基因组水平研究2个棕鳟群体的遗传多样性和群体结构,并通过群体间选择消除分析探究2个群体基因组中选择信号的差异,为棕鳟的分子辅助育种打下数据和方法基础。
1 材料与方法
1.1 样品收集和基因组DNA提取
于西藏亚东县亚东鲑鱼繁育基地和新疆乌鲁木齐市达坂城东沟冷水鱼实验场的2个棕鳟养殖群体中各采集12条棕鳟,共24个样本。剪取样本尾鳍1 cm,液氮速冻后提取基因组DNA,构建双端测序文库,使用Illumina Hiseq-x-ten测序平台进行全基因组随机测序,平均测序深度10X。
1.2 基因组重测序和SNP鉴定
依据已公布的棕鳟基因组序列作为参考序列(Hansen et al.,2021),使用Burrows-Wheeler Aligner将测序读段比对至参考基因组上(Li and Durbin,2009)。使用Genome Analysis Toolkit(McKenna etal.,2010)和SAMtools软件(Li et al.,2009)进行SNP鉴定,在默认参数下鉴定出SNP。使用VCFtools(Danecek et al.,2011),在-max-missing=0.2和mac=2的条件下进行过滤。
1.3 群体遗传多态性分析
计算4种与遗传多态性相关的统计量,包括核苷酸多态性(Pi)(Ness et al.,2010)、观测杂合度(Ho)、期望杂合度(He)和多态信息含量(PIC)(Chesnokov and Artemyeva,2015),计算Pi、Ho、He和PIC在基因组上的平均值,以衡量新疆棕鳟与西藏棕鳟的遗传多态性差异。
1.4 群体结构和亲缘关系分析
使用Plink将vcf文件转换为等位基因信息文件,并进行主成分分析(Principal components analysis,PCA)。使用IQ-Tree绘制系统发育进化树(Nguyen et al.,2015)。计算过程中选择使用经验碱基替换率的TVM模型(Transversion model),Bootstrap次数为1000。使用Admixture对2个棕鳟的群体结构进行分析(Alexander et al.,2009),并通过计算CV error进行验证。
1.5 选择印迹分析
使用PopLDdecay分析2个棕鳟群体的连锁不平衡衰减(Linkage disequilibrium decay)速度(Zhang et al.,2019),以确定群体中可能存在的选择信号。使用Vcftools在长度为50 kb、步长为10 kb的基因组长度滑窗中计算群体分化指数(FST)和Pi值,并进行基于FST和Pi的选择消除分析,筛选同时满足Pi值相差最大和FST值最大(top1%)的区域作为具体明显选择印迹的染色体候选区域,位于这些区域的基因则为可能受到选择的候选基因。
1.6 候选基因筛选
将在选择印迹分析中得到的染色体区段进行SNPeff注释(Cingolani et al.,2012),将其中发生错义突变、选择性剪接突变和基因调控原件突变的基因序列在KEGG代谢途径数据库中运行Blast‐KOALA进行通路分析,注释基因功能及参与的代谢途径。
2 结果与分析
2.1 群体遗传多样性分析结果
本研究中所涉及的测序数据共得到4694474928条Illumina序列(reads),与参考基因组的mapping率超过97%。经SNP Calling,共鉴定出8559684个SNP位点(表1)。在鉴定出的SNP中,碱基转换率略高于碱基颠换率,说明在棕鳟基因组中同样存在碱基转换偏差(赵辉等,2006)。

表1 样本及测序信息Table 1 Information of the samples and sequencing
对4种表征遗传多态性的统计量Pi、He、Ho、PIC进行计算和分析,发现2个群体的群体遗传多样性非常相近(表2),其中,西藏群体的Pi值约为0.348,仅略高于新疆的0.336;其他3个统计量也有类似结果。此外,无论是新疆群体还是西藏群体,群体内部的He和Ho值均较相近,符合哈迪—温伯格平衡,说明2 个群体内部的交配方式可能以随机交配为主(Chesnokov and Artemyeva,2015)。对比全基因组中2个群体所鉴定到的SNP密度,发现2个群体的SNP密度分布高度相似(图1),但在1号染色体的11~39 Mb处、4号染色体的2和18 Mb处、13号染色体的60 Mb处、18号染色体的10、27和29.7 Mb等处、20号染色体的6 Mb处、21号染色体的16~42.5 Mb处、25号染色体的25.1 Mb处、28号染色体的8、14和17 Mb处,以及31号染色体的7 Mb处的SNP密度差异明显。
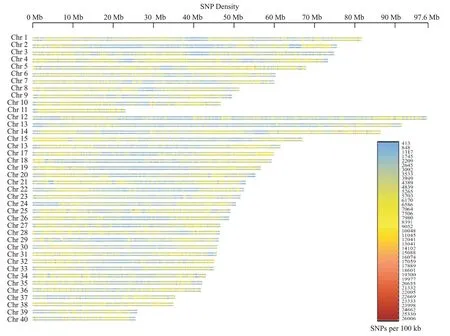
图1 新疆群体(上)和西藏群体(下)的SNP密度图Fig.1 SNP density diagram of Xinjiang(upper)and Tibetan(lower)populations

表2 群体遗传多样性比较Table 2 Genetic diversity comparison between the two popu‐lations
2.2 群体结构分析结果
在构建的系统发育进化树中,有新疆棕鳟个体与西藏棕鳟聚为一簇,说明这2个群体的亲缘关系较近(图2)。但在用列前3位的主成分数据绘制的三维PCA分析图中,发现主成分分析可将2个群体较好地分开,说明新疆棕鳟群体与西藏棕鳟群体在整体上存在一定的遗传距离(图3)。此外,在2个群体内部,各个体间的点分布不紧密,说明群体的遗传单一性不高,个体间存在一定的遗传差异。
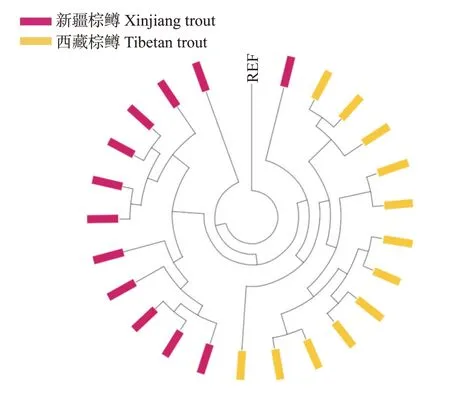
图2 新疆和西藏棕鳟个体系统发育进化树Fig.2 Phylogenetic tree of Xinjiang and Tibetan brown trout individuals
使用Admixture进行群体结构分析,当预测出的祖先群体数为2时(K=2)已不能将这2个群体完全分开(图4)。类似的,在CV error验证中,K=1时的CV error最低,说明新疆群体和西藏群体可能来自同一种源(图5)。2种分析方法均验证2个群体具有共同的祖先,结合其他学者对亚东鲑鱼种质来源的研究,基本上可确定新疆群体和西藏群体均来自于大西洋棕鳟居群(户国等,2016)。

图5 新疆和西藏棕鳟群体CV error验证Fig.5 CV error verification in Xinjiang and Tibetan brown trout populations
考虑到群体内连锁不平衡(LD)的发生,存在自然选择、遗传漂变、个体的迁入或迁出、近亲繁殖等因素影响,比较2个群体的连锁不平衡衰减速度(LDdecay)时,以LD降低到最大值一半时的平均LD长度来衡量连锁不平衡衰减速度,结果(图6)发现,2个群体连锁不平衡衰减速度几乎相等,新疆群体略高。在2个棕鳟群体中未观察到Liu等(2019)在7个黄瓜群体间发现的LD衰减曲线形状和数值显著差异,说明新疆群体和西藏群体经历的上述群体事件可能相似。

图6 新疆棕鳟和西藏棕鳟的连锁不平衡衰减Fig.6 Linkage disequilibrium decay of Xinjiang and Tibetan brown trout populations
2.3 群体选择消除分析结果
为寻找受到较强选择的染色体区域,通过鉴定得到的每个个体SNP位点,在2个群体间进行群体选择消除分析。结果(图7)显示,在群体Pi比值[log2(Pi疆/Pi藏)]最低的1%区域与FST最大的1%区域交集(图7左上角)所对应的染色体区域中,新疆群体的Pi低于西藏群体,且群体分化指数较高。推测在新疆群体的第5和第19号染色体上有大片段区域可能受到强烈的选择;与之相似,Pi比值最高的1%区域与FST最大的1%区域的交集(图7右上角)所对应的染色体区域中,西藏群体的Pi低于新疆群体,且群体分化指数较高,推测在西藏群体的第17和第18号染色体上有大片段区域可能受到强烈的选择。根据棕鳟的基因组注释信息(Hansen et al.,2021),在这些染色体区域上,共分布有94个编码蛋白基因,其中33来自新疆群体,61个来自西藏群体。

图7 新疆棕鳟和西藏棕鳟群体间基于FST和Pi的选择消除分析Fig.7 FST-and Pi-based selective sweep analysis between Xinjiang and Tibetan brown trout populations
使用KEGG代谢通路数据库对受到选择的基因可能参与的代谢途径进行分析,结果(表3)显示,这些基因分别参与多个不同的信号通路,且这些信号通路主要与信号分子传导、免疫和疾病等生理过程相关。群体选择消除分析中鉴定出位于受到强烈选择区域的94个基因中有21个基因与免疫或疾病相关,其中4个来自于新疆群体,17个来自于西藏群体(图8)。西藏群体中受到选择的基因所参与的通路主要包括PI3K-Akt信号传导途径、中性粒细胞胞外捕集器的形成、JAK-STAT信号传导途径、Hippo信号传导途径、表皮生长因子受体酪氨酸激酶抑制剂抗性、cAMP信号传导途径、细胞因子—细胞因子受体相互作用、FoxO信号传导途径、磷脂酰肌醇信号系统、神经活性配体—受体相互作用、mTOR信号传导途径、轴突引导、ECM-受体相互作用、细胞粘附分子和NOD样受体信号传导途径,主要位于第1、4、14、17、18、19、21、23、26、30和31号染色体上;而新疆群体的这些基因参与的代谢途径主要集中于MAPK信号传导途径、Ras信号传导途径、钙信号传导途径、TNF信号传导途径、PI3K-Akt信号传导途径和Hippo信号传导途径,主要位于第5、29和39号染色体上。

表3 在不同KEGG通路上受到选择的基因数量Table 3 Number of the involved genes under selection in different KEGG pathways

图8 新疆和西藏群体FST曼哈顿图Fig.8 Manhattan graph of FST between Xinjiang and Tibetan populations
3 讨论
基因组重测序在许多物种中能获取百万以上的SNP分子标记,是鱼类功能基因关联定位及群体遗传研究的有力工具。以大西洋鲑鱼性成熟年龄相关基因的研究为例,前人基于6.5K SNP阵列数据进行过全基因组关联分析,虽然得到了与性状关联的基因座,但未能获得具体的基因或基因位置(Gutier‐rez et al.,2015);Ayllon等(2015)基于野生和驯化的大西洋鲑鱼群体的基因组重测序数据,获得4326591个SNP标记,远高于Gutierrez 等(2015)采用SNP阵列获得的标记数量,同样通过全基因组关联分析,鉴定出vgll3基因是与大西洋鲑鱼性成熟年龄相关的基因。在基因组庞大,特别是经历过额外的全基因组加倍事件的鲑科和鲟科鱼类中(Berthelot et al.,2014;Lien et al.,2016;Du et al.,2020),以基因组测序数据鉴定SNP分子标记进行分析较其他分子标记更准确(豪富华,2006)。新疆达坂城和西藏亚东县的养殖棕鳟群体在SNP密度分布、连锁不平衡衰减速度和核苷酸多态性等方面存在高度的相似性,暗示这2个群体可能引自同一种源。尽管新疆棕鳟引入我国仅比西藏棕鳟晚150多年,但强烈的自然或人工选择压力能对棕鳟的进化产生影响,导致生物学特性在短期内即发生改变(Pigeon et al.,2016),对虹鳟鱼的多性状人工选育可达每代7%的遗传增益(Kause et al.,2005)。因此,在西藏和新疆的棕鳟养殖群体中可能存在适应性进化差异。与引自巴基斯坦的养殖棕鳟不同,西藏棕鳟有着更为悠久的野生历史,从1866年棕鳟被引入到1998年棕鳟的人工养殖研究取得成功,西藏棕鳟维持了130多年的野生状态。棕鳟相比西藏高原水体中的土著鱼类,其性成熟较早且生长速度快,与亚东河当地鱼类相比具有明显优势(豪富华,2006),使得棕鳟养殖在西藏获得成功,也可能是导致西藏棕鳟遗传多样性更高的原因。
选择消除是指在强烈选择压力的作用下,有利变异的等位基因频率会在较短时间内快速增加,其增加速度远高于正常水平,以至于有利变异邻近的稀有变异与之连锁地被保留下来,并导致有利变异邻近区域核苷酸多态性的减少。这是一种特殊的遗传搭车现象(Stephan,2019)。对来自不同群体的基因组重测序数据进行选择消除分析,能发现不同群体中受到强烈选择的候选基因。该方法已应用于罗非鱼和大西洋鲑鱼的驯化相关候选基因筛选(Na‐val-Sanchez et al.,2020;Nayfa et al.,2020),并在3个不同罗非鱼养殖群体间定位到多个与生长、早期发育和免疫特性相关的基因(Cádiz et al.,2020)。本研究中,选择消除分析得到的候选基因主要与免疫相关,或许由于新疆棕鳟和西藏棕鳟在引种前已适应高海拔水体,均经历过人为驯化,导致2个群体的适应差异主要集中在不同水体的病原体抗性上。
西藏棕鳟相比新疆棕鳟有着更高的群体遗传多样性,但西藏棕鳟受到选择的功能基因更多,功能分布更广,暗示着两地棕鳟经受的选择压力存在差异。单一的选择压力会导致群体的遗传多态性降低,并产生集中在少数染色体上的选择印迹,与新疆棕鳟群体中发现的选择印迹分布相似;而在西藏棕鳟中,发现的选择印迹分散在多条染色体的多个区段上,说明更可能是多种选择因素共同导致的结果。与引入我国前后均养殖在中低海拔地区(2300 m以下)的新疆棕鳟不同,生活在海拔2700~3700 m亚东河流域的西藏棕鳟在人工养殖成功前长期处于野生状态,且在1992年被列为西藏二级重点水生野生保护动物前经历了严重的过度捕捞,这些因素可能共同影响了西藏棕鳟中选择印迹的分布。
4 结论
新疆棕鳟和西藏棕鳟相近的亲缘关系和较小的遗传差异揭示两个棕鳟群体可能来自同一种源,2个群体受到选择压力较大的基因集中在免疫相关通路中,未发现存在明显选择信号的生长相关基因,说明新疆棕鳟和西藏棕鳟在引种后的人工选育程度不足,受到的选择压力仍以自然选择为主。
