香蕉果实MAPs测定方法的建立及其含量变化规律研究
2022-08-09孙陈康马兆成窦同心高慧君易干军
董 涛,孙陈康,马兆成,窦同心,高慧君,易干军*
(1广东省农业科学院果树研究所/农业农村部南亚热带果树生物学与遗传资源利用重点实验室/广东省热带亚热带果树研究重点实验室,广东广州 510640;2华中农业大学园艺林学学院/园艺植物生物学教育部重点实验室,湖北武汉 430070)
0 引言
【研究意义】香蕉是一种常见的水果,同时也是重要的粮食作物,其气味芬芳、甘甜爽口、肉软滑腻,深受人们喜爱(Pino and Febles,2013)。前人研究表明,香蕉果实中富含血清素、色氨酸、左旋多巴、儿茶酚胺等多种单胺类及其前体物质(Monoamines and their precursors,MAPs),具有抗氧化、抗抑郁等多种药用和保健功能,有极高的研究与应用价值(Volkow et al.,2012;洪佳敏等,2016;Singh et al.,2016;Szeitz and Bandiera,2018;Zhao et al.,2019)。因此,建立一种特异、准确、快速检测香蕉果实MAPs的方法,明确香蕉果实发育过程中MAPs各组分的含量变化规律,对深入研究其在果实不同发育阶段的积累特点与形成代谢机制具有重要意义。【前人研究进展】MAPs包括单胺类物质血清素(Serotonin,SERO)、多巴胺(Dopamine,DA)、去甲肾上腺素(Norepineph‐rine,NE)、肾上腺素(Adrenaline),以及其前体物质色氨酸(Tryptophan,Trp)、色胺(Tryptamine,TRM)、5-羟色氨酸(5-hydroxy-tryptophan,5-HTP)、酪氨酸(Tyrosine,Tyr)、酪胺(Tyramine,TYRA)、左旋多巴(Levodopa,L-dopa)等(Hornykiewicz,2010;Mauro et al.,2013;Zhang et al.,2019;Hu et al.,2020)。以往文献中关于香蕉果实中MAPs含量检测的研究不多。1958年,香蕉果肉和果皮中被检测到DA、SERO、NE和5-HTP,这是首次在香蕉中检测到MAPs的存在(Anderson et al.,1958;Waalkes et al.,1958)。Kanazawa和Sakakibara(2000)对催熟期间Cavendish香蕉果实的DA、L-dopa、NE和抗坏血酸等多种具有抗氧化活性的化合物进行分离,同时对抗氧化物质进行定量,发现了一种抗氧化活性很强的水溶性物质DA,果肉中DA含量达2.5~10 mg/100 g。González-Montelongo等(2010)研究表明,用各种溶剂纯溶液或其水溶液浸提香蕉皮,提取物中能鉴定出DA和Ldopa,且不同的溶剂浸提能获得不同的DA和L-dopa提取率。SERO及其前体物质Trp和TRM存在于多种果蔬中,日本研究人员对38种常见果蔬中SERO、Trp和TRM含量进行定量分析,结果显示,香蕉、猕猴桃、凤梨、鳄梨、番茄和土豆中SERO和Trp含量较其他果蔬高(Islam et al.,2016)。核桃也含有较丰富的Trp和SERO(Schröder et al.,1999)。Borges等(2019)挑选了20个香牙蕉和大蕉品种进行催熟处理,选择3个催熟时期(绿熟期、黄熟期和完熟期)进行DA、L-dopa、TYRA和SERO等8种生物胺含量的检测,结果表明DA、L-dopa、TYRA和SERO含量在果实催熟过程中呈下降趋势,同时研究不同处理工序对果实中生物活性胺含量的影响,发现带皮水煮后煮食蕉Pelipita果肉中DA、NE、精胺和亚精胺含量均有所提升。【本研究切入点】目前,国内外有关香蕉果实发育过程中MAPs含量变化规律的研究未见报道。【拟解决的关键问题】以目前国内主栽的巴西香蕉(AAAcv.Baxi)、粉杂1号粉蕉(ABB group,Fenza 1)和东莞大蕉(ABB group,Dongguan Dajiao)为研究对象,建立一种特异、准确、快速测定香蕉果实MAPs的方法,利用该方法系统地探讨香蕉果实发育过程中MAPs的积累和变化规律,以期为进一步研究其合成与代谢机制提供参考依据。
1 材料与方法
1.1 试验材料
供试材料为巴西香蕉、粉杂1号粉蕉和东莞大蕉3个栽培品种发育过程中的香蕉果实,种植于广东省农业科学院果树研究所增城实验基地,果园生长环境条件和水肥管理均一致。选取生长健壮且长势一致、无病虫害的3个香蕉品种植株各30株。使用的所有试剂和药品均为色谱级,NE、DA、L-dopa、TYRA、Tyr、SERO、TRM和Trp标准品购自广州康龙生物科技有限公司,超纯水、三氟乙酸和甲醇用于配制流动相。主要仪器设备包括高效液相色谱仪(Agilent 1290,美国安捷伦科技有限公司)和光电二极管列阵检测器(DAD-3000,上海伍丰科学仪器有限公司)。
1.2 试验方法
1.2.1 取样方法 试验材料于2020年4月15日—7月3日采集。香蕉蕉果断蕾后,每隔15 d进行一次取样,共采集5个时期的样品,分别为断蕾后15、30、45、60和75 d。其中,断蕾后75 d取样时间根据不同品种香蕉果实发育情况进行调整,以果实成熟度达八成熟为标准。巴西香蕉和粉杂1号粉蕉断蕾后75 d采收时间分别为蕉果断蕾后的75和78 d,果指饱满度约为八成(巴西香蕉果实直径为3.37 cm或围长100 mm、粉杂1号粉蕉直径为3.93 cm或围长为130 mm)(黎源,2014)。由于东莞大蕉采收标准未见报道,因此参照巴西香蕉和粉杂1号粉蕉采收时间与方法自行确立,经5次采样统计分析得出(自上而下取第二把蕉果中间位置,左右排开连续取7根果指进行测量统计),果实饱满度为八成熟时,直径为3.96 cm或围长为138 mm。选择树体健壮、生长正常的果树取果,选取每条蕉的第二把作为试验样品,同时采摘3株植株上面的果实,作为3个生物学重复。
1.2.2 样品制备 用自来水冲洗果实,以清除碎屑或其他杂质。去除果尖和果柄,切取果指中间部分,沸水中静置10 s,使多酚氧化酶失活(Jiménez and García-Carmona,1999)。剥皮,皮肉分离,切片后立即液氮速冻,存放于-80 ℃冰箱中备用。将待测果皮和果肉放入冷冻干燥机去除水分,得到的干燥样品用研磨机研磨成粉;分别取1 g香蕉果肉或果皮冻干粉,加入20%甲醇5 mL和0.1%三氟乙酸水溶液5 mL,用涡旋仪振荡15 s,重复3次,放入4 ℃冰箱冷藏过夜;3000 r/min离心30 min,取3 mL上清液氮吹浓缩至1 mL,12000 r/min离心5 min,0.22 μm滤膜过滤,置于上样瓶,同一品种重复提取3次,放入4 ℃冰箱待上样检测。
1.2.3 标准品溶液制备 精密称取NE、DA、L-do‐pa、TYRA、Tyr、SERO、TRM和Trp标准品各5 mg,加入20%甲醇水溶液5 mL,振荡摇匀至完全溶解,制成1 mg/mL的标准溶液。0.22 μm滤膜过滤,置于棕色瓶,放入4 ℃冰箱待上样检测。
1.2.4 反向高效液相色谱—紫外检测(RP-HPLCUV)条件 所用色谱柱为C色谱柱(Eclipse Plus 95Å Ccolumn,4.6×250 mm,5 µm,Agilent)。甲醇为流动相A,0.1%三氟乙酸水溶液为流动相B,pH 3.1。使用梯度剖面对待分离化合物进行洗脱分离:0~14 min,线性梯度5%~30% A,95%~70% B,柱温28 ℃、流速1.0 mL/min。每个样品的总运行时间为30 min,进样量5 μL,洗脱条件如表1所示。所有样品检测均进行3个重复。
1.2.5 MAPs定量分析统计 香蕉样品色谱图中目标物质峰峰面积,根据每个目标化合物的线性回归方程计算样品溶液浓度。
1.2.6 方法的适应性验证 验证参数包括特异性、准确性、精确性、检测限和定量限等。特异性:配制浓度为50.0 μg/mL的各标准化合物溶液,分别加入待测样品中,和相同份数但未加标的样品一起进行检测分析,以确认每种峰的同一性,以此确定本检测方法在香蕉样品检测中的特异性。准确性:用加标回收率验证检测方法的准确性,通过分析5个重复的加标样品确定方法的准确度。精确性:日内重复性和日间重复性检验方法的精确性,在1 d 3个时间段和在3 d内对5个重复样品进行检测,分别计算相对标准偏差(RSD)。检测限(LOD):信噪比为3时的标准化合物样品浓度,即峰高是基线噪声3倍高度时的标准化合物样品浓度为LOD。定量限(LOQ)=LOD×10/3。
1.3 统计分析
利用SISVAR(V1.2)对试验数据进行方差分析和Tukey Test(<0.05)均值比较检验(Ferreira,2011),以Excel 2007制图。
2 结果与分析
2.1 香蕉果实中MAPs检测方法的建立
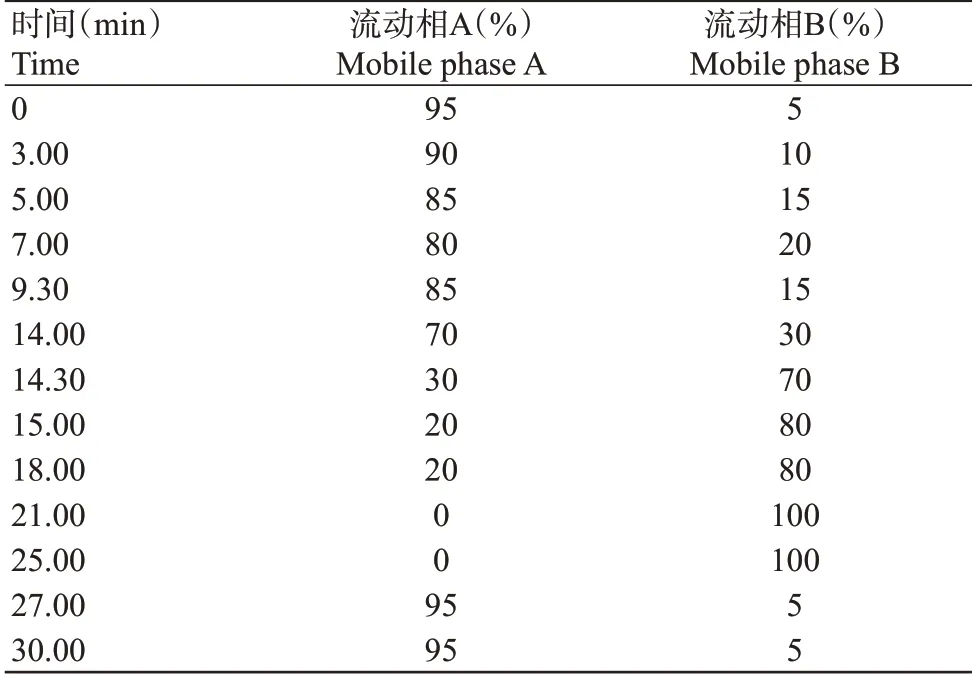
利用本研究建立的RP-HPLC-UV法,可以准确、快速地测定香蕉果肉和果皮中NE、DA、L-dopa、TYRA、Tyr、SERO、TRM 和Trp 等8 种MAPs 含 量。MAPs紫外吸收波长光谱(图1)显示,在210~350 nm波长范围内实现物质峰校准,通过比较保留时间和最大吸收波长鉴定每个化合物物质峰。TYRA和Tyr最大吸收波长为276 nm,SERO最大吸收波长为278 nm,TRM和Trp最大吸收波长为280 nm,NE、DA和L-dopa最大吸收波长为282 nm。从样品色谱图(图2)可看出,8种MAPs均成功分离,基线平稳无漂移,峰型对称且良好,无重叠峰、驼峰、前沿和拖尾现象,保留时间稳定。
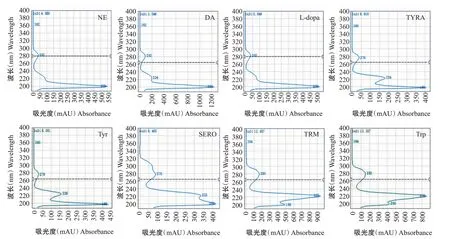
香蕉果实8种MAPs定量检测方法的适用性验证参数如表2所示。校准曲线的线性判定系数均高于0.999000,表明浓度与信号面积之间存在高度相关性。因此,校准曲线的线性方程适用于香蕉中单胺类及其前体物质的定量分析,线性范围为1.0~16.0 μg/mL。方法的准确度通过加标样品中分析物的回收率进行评估,范围为95.3%~106.1%,说明该方法具有较高的准确性。日内精密度范围为0.37%~0.82%,日间精密度范围为0.35%~0.83%,表明该方法同时具有较高的精准性。检测限为0.0012~0.0016 mg/100 g,定量限为0.0040~0.0053 mg/100 g,显示该方法具有很好的灵敏度。8种定量检测的MAPs在色谱柱中被完全分离开,且峰型良好,无重叠峰,色谱图基线平稳无起伏,待检测物质在14 min内全部完成分离。

2.2 NE和DA含量的变化
香蕉果实发育过程中,巴西香蕉中NE含量先升后降,果肉和果皮中含量均在断蕾后30 d达峰值,而东莞大蕉和粉杂1号果实中NE含量随果实发育变化不明显(图3-A和图3-C)。3个品种果皮中,巴西香蕉的NE含量最高(35.52~62.18 mg/100 g),东莞大蕉次之(5.82~9.29 mg/100 g),粉杂1号最少(1.33~2.05 mg/100 g);3个品种果肉中也呈现类似规律,巴西香蕉的NE含量为3.17~22.14 mg/100 g,东莞大蕉为1.27~2.07 mg/100 g,粉杂1号为0.91~1.33 mg/100 g。3个品种果皮中NE含量均高于果肉。
DA与NE的含量变化趋势基本一致。巴西香蕉中DA含量呈先升后降的变化趋势,果肉和果皮中均在断蕾后30 d达峰值,含量分别为239.81和1906.54 mg/100 g,断蕾后75 d达最低值(7.08和793.44 mg/100 g),东莞大蕉果肉中DA含量随果实发育逐渐降低(4.80~23.67 mg/100 g),而粉杂1号果肉中DA含量随果实发育变化不明显(4.47~6.07 mg/100 g)(图3-B和图3-D)。3个品种果皮中DA含量均高于果肉,且与NE变化规律类似,巴西香蕉果皮中DA含量最高(793.44~1906.54 mg/100 g),东莞大蕉次之(378.92~631.63 mg/100 g),粉杂1号最低(103.52~167.81 mg/100 g)。

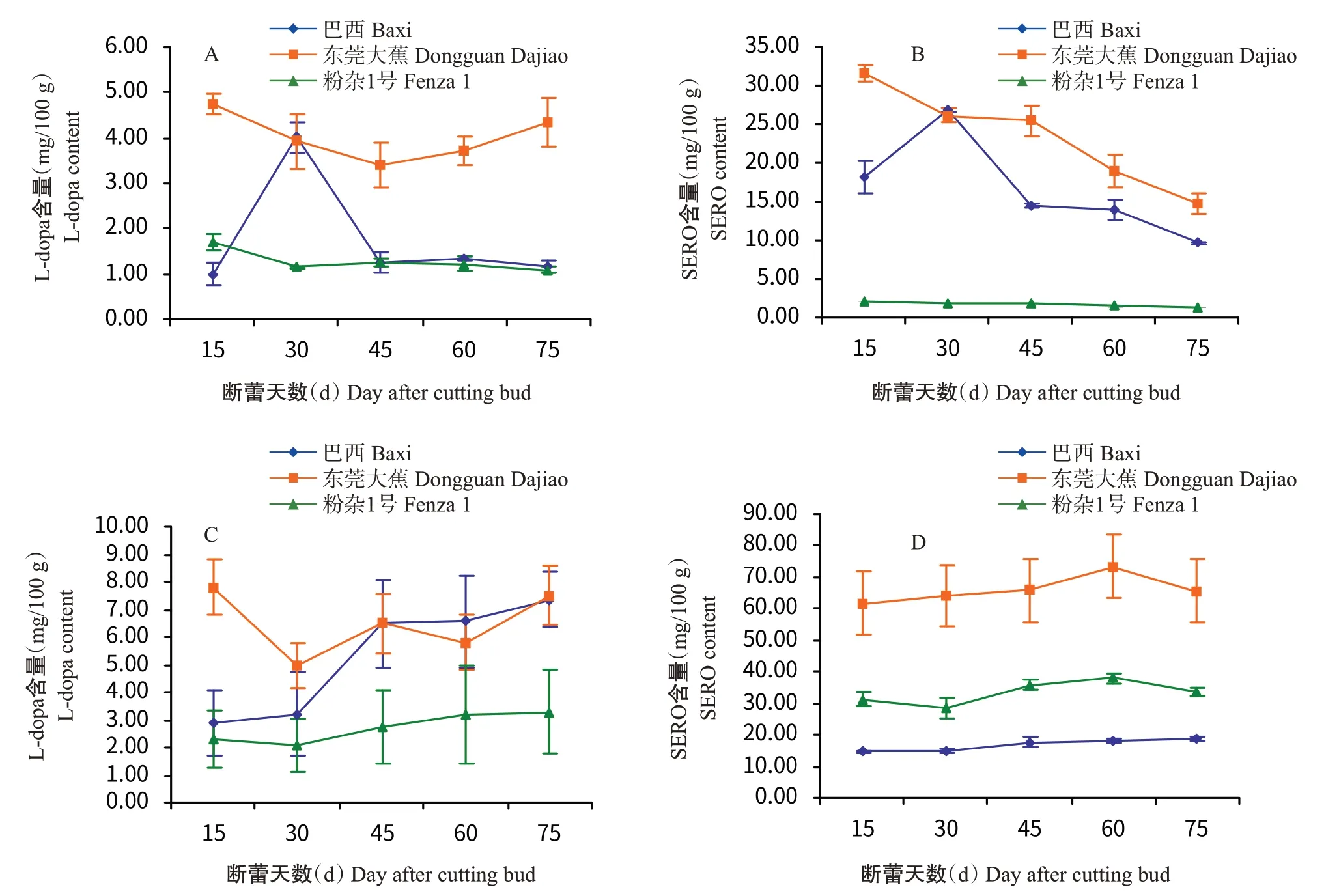
2.3 L-dopa和SERO含量的变化
在果实发育过程中,巴西香蕉果肉中L-dopa含量先升后降,在断蕾后30 d达峰值(4.02 mg/100 g),而果皮中L-dopa含量整体呈上升趋势(图4-A和图4-C)。东莞大蕉果肉和果皮中L-dopa含量变化规律基本一致,呈先降后升的变化趋势。粉杂1号果肉中L-dopa含量呈下降趋势,果皮中含量呈逐渐上升趋势。3个品种果皮中L-dopa含量均高于果肉。粉杂1号果皮中L-dopa含量(2.08~3.29 mg/100 g)则低于巴西香蕉(2.91~7.35 mg/100 g)和东莞大蕉(4.95~7.81 mg/100 g)。
在果实发育过程中,巴西香蕉果肉中SERO含量先升后降,在断蕾后30 d达峰值(26.95 mg/100 g),果皮中含量则呈缓慢上升趋势。东莞大蕉果肉中SERO含量随着果实的发育不断降低,果皮中含量呈先升后降的变化趋势,在断蕾后60 d达峰值(73.31 mg/100 g)。粉杂1号果肉中SERO含量呈下降趋势,果皮中的含量与巴西香蕉果皮中变化趋势基本一致。东莞大蕉和粉杂1号果皮中SERO含量明显高于果肉(图4-B和图4-D)。
2.4 Tyr和TYRA含量的变化
由图5可知,Tyr和TYRA含量在3个品种的果肉和果皮中均呈现出类似的变化规律。果肉中,巴西香蕉的Tyr和TYRA含量随着果实发育急剧升高,在断蕾后30 d达峰值(43.74和118.29 mg/100 g),之后迅速降低,Tyr含量在断蕾后45 d达最低值(1.14 mg/100 g),TYRA含量在断蕾后75 d达最低值(12.58 mg/100 g);东莞大蕉和粉杂1号中的含量则变化不明显(图5-A和图5-B)。果皮中,3个品种Tyr和TYRA含量整体呈下降趋势,东莞大蕉果皮中Tyr含量高于巴西香蕉和粉杂1号;TYRA含量则在巴西香蕉中最高(128.41~374.94 mg/100 g),东莞大蕉次之(19.28~110.33 mg/100 g),粉杂1号最低(4.82~9.59 mg/100 g)(图5-C和图5-D)。

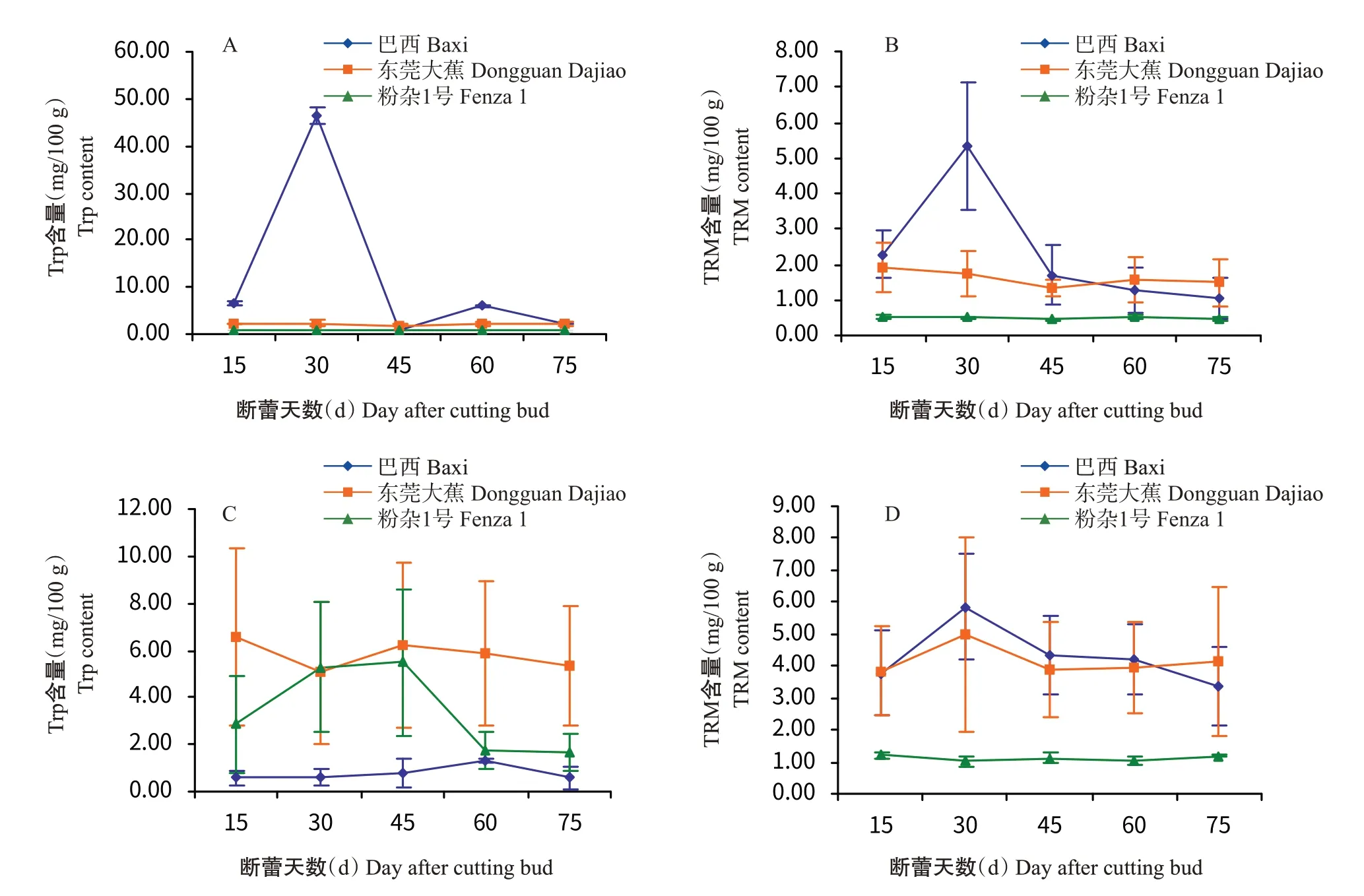
2.5 Trp和TRM含量的变化
由图6-A和图6-B可知,Trp和TRM含量在3个香蕉品种的果肉中表现出类似的变化趋势,巴西香蕉果肉中Trp和TRM含量均在断蕾后30 d达最高值(46.41和5.33 mg/100 g),而后急剧降低,Trp含量在断蕾后45 d达最低值(0.90 mg/100 g),TRM含量在断蕾后75 d达最低值(1.04 mg/100 g);东莞大蕉和粉杂1号果肉中的Trp和TRM含量随果实发育变化不明显。巴西香蕉和东莞大蕉果皮中Trp含量在果实发育过程中无明显变化,粉杂1号果皮中则有先升后降的变化过程;3个品种中,东莞大蕉果皮Trp含量最高,其次是粉杂1号,巴西香蕉最少(图6-C)。3个品种果皮中,TRM含量随果实发育整体变化不明显,粉杂1号的含量明显低于巴西香蕉和东莞大蕉(图6-D)。
3 讨论
MAPs的检测方法主要包括薄层色谱法(TLC)、放射免疫分析法(RIA)、酶联免疫吸附法(Enzymelinked immunosorbent assay)和高效液相色谱法(HPLC)。其中,HPLC检测准确度高、灵敏度高、分析成本相对较低、操作简便,配合不同类型检测器,能进行多种类MAPs的检测,包括DA、NE和L-dopa等化合物和其他多胺类物质(Islam et al.,2016)。近年来,有研究利用微芯片电泳紫外荧光技术检测香蕉中的DA、SERO、Trp和Tyr等天然化合物含量(Ohla et al.,2011),但其检测成本高,不利于大量检测。液相或气相色谱连接不同检测器进行定性定量分析仍是目前测定植物中MAPs的最常用方法。HPLC-UV相较高效液相色谱—质谱联用(HPLCMS)一次检测物质种类较少,但价格便宜,产业应用价值更大。本研究利用RP-HPLC-UV建立了香蕉中NE、DA、L-dopa、TYRA、Tyr、SERO、TRM和Trp等代谢物的检测体系,可以准确、快速地测定不同香蕉组织中8种MAPs含量,且操作方便,成本较低。
González-Montelongo等(2010)研究香蕉果皮抗氧化成分的最佳提取条件时发现,甲醇水溶液更有利于提取香蕉果实中的DA,而100%异丙醇更有利于提取L-dopa。本研究采用20%甲醇和0.1%三氟乙酸水溶液对香蕉果实MAPs物质进行提取,结果表明,8种化合物可以被提取出来。其中,果皮样品中检测到DA含量最高达1906.54 mg/100 g,远高于Kanazawa和Sakakibara(2000)检测到的最高值1290 mg/100 g,表明本研究的提取条件非常适用于香蕉果实中DA的定量检测。但本研究中检测到的TYRA、TRM和L-dopa等含量较低,说明上述物质可能更适合利用HPLC-MS等精密度更高的检测技术进行测定分析。因此,本研究所用提取条件更有利于香蕉果实中DA的提取,而其他种类MAPs的最佳提取条件需进一步摸索。
目前,国内外对香蕉等水果中MAPs含量的检测分析有一些研究,但尚未见香蕉果实发育过程中MAPs含量检测的相关报道。Kanazawa和Ashida(1998a,1998b)研究发现,香蕉皮中含有强抗氧化活性化合物DA,其抗氧化活性与维生素C相当,认为DA能帮助增强人体消化道清除脂质过氧化产物的能力。本研究结果表明,3个香蕉品种果肉中均含有DA,但其含量明显低于果皮,与前人研究结果(Borges et al.,2019)基本一致。Borges等(2019)推荐消费者食用带皮水煮后的香蕉果肉,因水煮后的香蕉皮中DA会转移到果肉中,使果肉中DA和SERO含量均有所上升。除了DA外,本研究的3个香蕉品种果皮中NE、L-dopa、SERO和TYRA含量也明显高于果肉;DA、NE、L-dopa和TYRA均位于香蕉果实多巴胺代谢通路上,L-dopa和TYRA是合成DA和NE的前体物质,上述结果表明在香蕉果皮中可能存在着更为活跃的多巴胺合成代谢过程。Kanazawa和Sakakibara(2000)研究发现卡文迪许香蕉果皮中DA含量为865~1290 mg/100 g,本研究结果与其基本一致,但果肉中DA含量为0.72~10 mg/100 g,远低于本研究结果(7.08~239.81 mg/100 g)。Borges等(2019)检测了20个不同香蕉品种果实中DA含量的变化情况,其中鲜食蕉DA含量最高的品种为Yangambi Km5,其果肉中DA含量为26.5~38.1 mg/100 g,果皮中DA含量最高的鲜食蕉为Ouroda Mata,含量为76.4~305.5 mg/100 g,均低于本研究结果。造成研究结果差异的原因,一方面是因为品种、检测方法和种植区域不同,另外一个主要原因可能是研究对象所处的发育时期不同。本研究结果表明,巴西香蕉的DA含量随着果实发育有快速升高的过程,峰值出现在断蕾后30 d前后(果肉239.81 mg/100 g 、果皮1906.54 mg/100 g),之后果实中DA含量持续下降,果实成熟时达到最低值(果肉7.08 mg/100 g、果皮793.44 mg/100 g)。Borges等(2019)、Kanazawa和Sakakibara(2000)的研究材料均在香蕉果实成熟时采集,可能是造成其研究结果明显低于本研究结果的主要原因。
不同香蕉品种MAPs的含量有明显差异。断蕾后15~30 d,巴西香蕉果肉中NE、DA、TYRA、Tyr、TRM和Trp含量均快速上升,L-dopa和SERO含量也有缓慢上升的趋势,之后,上述物质含量呈下降趋势。在巴西香蕉果实整个发育过程中,MAPs含量变化极大,含量相差2倍(TRM)~50倍(Trp)不等。东莞大蕉果肉中仅有SERO含量明显降低,DA含量缓慢下降,NE、L-dopa、Tyr、TYRA和TRM含量的变化不明显。粉杂1号MAPs含量相对较少,且在整个果实发育过程中大部分MAPs含量变化不明显。查询KEGG数据库(https://www.kegg.jp/)表明,上述8种MAPs均为Tyr和Trp代谢通路上的化合物,其含量变化与苯丙氨酸、酪氨酸和色氨酸的合成及代谢通路(Mus00400)紧密有关。因此,推测在巴西香蕉果实发育成熟过程中,可能在断蕾后30 d之前的某个时间段,某些或某个调控果实MAPs合成代谢的基因表达水平发生显著改变,从而导致Mus00400代谢通路上包括MAPs在内的化合物含量发生变化,而东莞大蕉和粉杂1号中不存在上述基因表达水平显著改变的过程;品种间产生上述差异的原因可能与品种的基因型有直接关系,巴西香蕉基因型为AAA,而粉杂1号粉蕉和东莞大蕉基因型均为ABB。因此,后续可根据上述特点,研究香蕉果实断蕾30 d前后与MAPs合成和代谢相关基因的表达情况,为下一步解析香蕉果实MAPs代谢调控机制打下基础。
4 结论
利用RP-HPLC-UV建立了一种测定香蕉果实中MAPs含量的方法,可以准确、灵敏、快速地测定香蕉果肉和果皮中NE、DA、L-dopa、TYRA、Tyr、SERO、TRM和Trp等8种MAPs物质的含量。通过对3个香蕉品种不同发育阶段的果实MAPs含量进行测定,发现巴西香蕉果实发育过程中MAPs含量变化明显区别于东莞大蕉和粉杂1号粉蕉。本研究结果可为下一步利用基因组数据筛选不同基因型的差异表达基因,并进一步阐明MAPs的合成代谢机理提供参考。
