金属离子对中间锦鸡儿2C 型丝/苏氨酸蛋白磷酸酶酶活的体外调节*
2022-08-06刘昱成韩晓东雷艳红高仙灵李国婧贾永红
刘昱成 韩晓东 雷艳红 高仙灵 戴 玉 李国婧 贾永红**
(1.内蒙古农业大学植物逆境生理与分子生物学自治区重点实验室,内蒙古 呼和浩特 010018;2.内蒙古赤峰市翁牛特旗农牧局,内蒙古 赤峰 024500;3.呼和浩特职业学院,内蒙古 呼和浩特 010051)
植物中存在着多种复杂的信号通路,共同调节植物的生命活动,蛋白质的磷酸化/去磷酸化是生物调节细胞过程的一种基本机制[1],是细胞信号通路的重要组成部分[2]。蛋白质的磷酸化和去磷酸化分别与蛋白激酶和蛋白磷酸酶有关,丝/苏氨酸蛋白磷酸酶家族根据序列的相似性可以分为PPP和PPM 2 个亚家族,PPP 亚家族包括PP1、PP2A、PP2B 和PP4-7,而PPM 亚家族则是由依赖锰离子和镁离子的蛋白磷酸酶组成。2C 型蛋白磷酸酶(PP2Cs)属于PPM 亚家族[3],是蛋白磷酸酶中最大的一个亚家族,被认为在信号途径中作为负调控因子起作用,通过与蛋白激酶相互拮抗进而调控蛋白质的磷酸化程度。
PP2Cs 参与多种激素的信号通路,包括脱落酸(ABA),乙烯等[2,4],目前研究比较深入的是PP2Cs在ABA 信号通路中的作用[5,6]。ABA 是植物中抵抗生物胁迫和环境胁迫的重要激素,可以调节多种基因的表达,是一种与渗透胁迫相关的植物激素[7]。研究发现,PP2Cs 在ABA 信号通路中有着重要的作用,如调控植物抗病[8]、ABA 依赖的植物发育(包括种子的萌发、根的伸长和气孔的开闭等[9-12])、非生物胁迫(主要为干旱和盐胁迫[6,13])和不依赖ABA的抗逆境胁迫[14]等过程。Xue T 等在拟南芥中鉴定出80 个PP2Cs 基因,水稻中有78 个。生物信息学分析发现PP2C 家族是目前发现的植物中最大的基因家族之一。系统发育分析将拟南芥和水稻中的PP2Cs 分别划分为11(包括A-E、F1、F2、G-H家族)和13 个亚家族(包括A-E、F1、F2、G-L 家族),不同亚家族的植物PP2Cs 参与不同的信号转导途径[1]。
中间锦鸡儿(Caragana intermedia)属于豆科锦鸡儿属灌木,生命力顽强,广泛分布于内蒙古干旱以及半干旱地区[15],是防治水土流失和土地荒漠化的优良造林树种之一,对于保护环境及维持生态平衡有着重要意义[16,17]。本研究利用原核表达系统对中间锦鸡儿的CiPP2C37-like 蛋白进行了表达,研究了不同因素对CiPP2C37-like 的酶活性影响。
1 研究材料、实验试剂及仪器
1.1 研究材料
野生型中间锦鸡儿成株叶片采自内蒙古乌兰察布市四子王旗。
1.2 主要试剂
Imidazole、NaCl(分析纯)、Tris-HCl、E·coliDH5α、E·coliBL21、琼脂糖凝胶DNA 回收试剂盒(Tiangen)、植物总RNA 提取试剂盒(Tiangen)、质粒小提试剂盒(上海生工)、反转录试剂盒(全式金)、EGTA、MgCl2(分析纯)、BSA(北京酷来博科技有限公司)、β-巯基乙醇、Ser/Thr Phosphatase Assay System(普洛麦格)、酵母粉(Thermo)、蛋白胨(Thermo)、IPTG (北京酷来博科技有限公司)。
1.3 主要仪器
PCR 扩增仪(德国SENSO)、台式冷冻离心机(美国Backman Coulter)、灭菌锅(日本三洋)、凝胶成像仪(德国SYNGENE)、定量紫外检测仪(美国Quawel)、水平槽电泳仪(中国六一有限公司)、分析天平(德国Sartorius)、恒温培养箱(上海福玛有限公司)、pH计(德国Sartorius)、酶标仪(美国Bio Tek)、NTA-亲和层析柱(上海生物工程有限公司)、超滤管(美国MiLipore)。
2 实验方法
2.1 构建CiPP2C37-like 系统发育树
从NCBI 数据库中选择已知功能的PP2C 基因,利用MEGA 软件基于1 000 次重复进行系统进化树分析。
2.2 表达目的蛋白
2.2.1 RT-PCR 获得中间锦鸡儿cDNA
取一定量的新鲜野生型中间锦鸡儿叶片于液氮中保存,将样品放在研钵中低温研磨,采用总RNA提取试剂盒(Tiangen)提取中间锦鸡儿RNA,提取步骤均按说明书进行。1%琼脂糖凝胶电泳检验后用反转录试剂盒(全式金)对柠条总RNA 进行RTPCR,具体操作均按说明书进行。得到中间锦鸡儿的cDNA 存于-20 ℃冰箱,用于后续实验。
2.2.2 构建CiPP2C37-like 基因原核表达载体
根据cDNA 末端快速扩增技术得到的CiPP2C37-like 基因序列[16],利用Primer Premier 5.0 设计引物,以柠条cDNA 为模板进行PCR 扩增,扩增条件为94℃预变性2 min;94 ℃,30 s;55 ℃,30 s;72 ℃,2 min,30 个循环;72 ℃,5 min;1%琼脂糖凝胶电泳验证。采用琼脂糖凝胶DNA 回收试剂盒(Tiangen)回收目的基因,具体操作步骤按说明书进行。利用T4DNA 聚合酶的外切活性产生的粘性末端将目的基因连接到pET-30a 载体上,连接产物42 ℃,热激45 s 后转入克隆菌株DH5α 中。利用卡那霉素抗性平板筛选阳性克隆后,挑取单菌落于试管中扩增菌体。用质粒小提试剂盒提取质粒,使用Xho I、Xba I进行双酶切验证后送华大基因公司测序。
2.2.3 诱导表达CiPPC37-like 蛋白
将重组质粒42 ℃,热激45 s 后转入表达菌株E·coli BL21 中。利用卡那霉素抗性平板筛选阳性克隆,将筛选出的阳性克隆接入终浓度为0.05 mg/mL卡那霉素的100 mL LB 培养基中,37 ℃震荡培养6 h后,将100 mL 菌液接入含有终浓度为0.05 mg/mL 卡那霉素的1L LB 液体培养基中继续震荡培养2 h,终浓度0.1 mM IPTG、16 ℃过夜诱导。
2.2.4 纯化CiPPC37-like 蛋白
诱导12 h 后,5 000 r,离心15 min 收集菌体,用30 mL NTA-0(25 mM Tris-HCl,0.5 M NaCl,pH=8.0)重悬菌体。加入终浓度为1 mM 的DTT,在超声波信号发生器中超声2 s,间隔4 s,破碎20 min。18 000 r,离心40 min 去除溶液中破碎的菌体。采用Ni-NTA柱纯化蛋白上清,分别用25 mL 含有0、10、20、50、100、200、500 mM 咪唑的NTA-0 洗脱目的蛋白,收集含有不同浓度咪唑的NTA-0 洗脱的穿透液,进行SDS-PAGE 电泳分析。采用超滤浓缩去除穿透液中含有的磷离子,得到的粗酶液用于后续酶活的测定[18]。
2.3 CiPPC37-like 蛋白酶活的测定及计算
CiPP2C37-like 磷酸酶活性的测定采用丝苏氨酸磷酸酶测定系统(普洛麦格),每个反应体系为50 μL,使用NTA-0 调整蛋白浓度使每个体系中蛋白含量在5 μg 以下。将10 μL 反应缓冲液(50 mM咪唑,pH7.2,5 mM MgCl2,0.2 mM EGTA 和0.1 mg/mL牛血清白蛋白)与5 μL 多肽底物(随丝苏氨酸磷酸酶测定系统试剂盒提供)预混匀后在对应反应温度下孵育3 min,加入粗酶样品反应15 min。室温静置15 min,待颜色稳定后测定600 nm 处的吸光值。
酶活力的定义为:在一定条件下,每分钟释放出1 μmoL 的磷酸盐的酶量为一个酶活力单位(U)。计算公式如下:
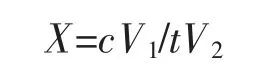
式中,X 为磷酸酶的酶活力,U/mL;c 为磷酸盐的浓度,μmol/L;V1为反应的总体积,mL;V2为粗酶液的用量,mL;t 为作用时间,min[19]。
2.3.1 磷酸盐标准曲线的绘制
用无磷水(随丝苏氨酸磷酸酶测定系统试剂盒提供)以1∶20 的比例稀释磷酸盐标准(随丝苏氨酸磷酸酶测定系统试剂盒提供)。向96 孔板中分别加入0、2、4、10、20 μL和40μL稀释后的磷酸盐标准液,用1×PP2C 缓冲液将体系补足50 μL。向每个梯度中加入等体积的钼酸盐染料显色,待颜色稳定后测定600 nm 处的吸光度[20]。
2.3.2 测定不同温度、pH 值、金属离子对CiPP2C37-like 磷酸酶活性的响应
在20~60 ℃的温度范围内,以5 ℃为间隔设定温度梯度,测定不同温度下的酶活变化。加入等体积的钼酸盐染料显色,待颜色稳定后测定600 nm处吸光值。分别配置pH 为5.5、6、6.5、7、7.5、8、8.5、9 的5×PP2C 缓冲液,测定不同pH 值下的酶促反应体系的酶活。金属离子能够与酶活性中心结合,参与电子的转移反应,稳定酶的构象,对酶活力有着较大的影响。配置终浓度为50 mM 的金属离子(Cu2+、Li+、K+、Na+、Ba2+)5×PP2C 缓冲液,调节pH=8.0,以不添加其他金属离子的PP2C 缓冲液配置酶活体系为对照,测定酶活性。
2.3.3 CiPP2C37-like 磷酸酶对磷酸肽的亲和性
将1 mM 的磷酸肽(随普洛麦格试剂盒提供)按比例分别稀释至0.25、0.5、1 mM。每50 μL反应体系加入5μL底物,底物终浓度分别为0.25、0.1、0.05mM。测定600 nm 处的吸光值,采用双倒数作图法计算Km 值。
3 结果与分析
3.1 原核表达系统表达CiPP2C37-like 蛋白
3.1.1 中间锦鸡儿cDNA
为了获得中间锦鸡儿cDNA,以中间锦鸡儿叶片为材料,采用总RNA 提取试剂盒(Tiangen)提取中间锦鸡儿总RNA,经琼脂糖凝胶电泳验证(图1),提取的总RNA 略有降解,经定量紫外检测仪检测,A260/A280=2.153,可用于RT-PCR。

图1 中间锦鸡儿总RNA 琼脂糖凝胶电泳
3.1.2 CiPP2C37-like 全长cDNA 的克隆及构建pET-30a::CiPP2C37-like 表达载体
以中间锦鸡儿cDNA 为模板进行PCR 反应,获得CiPP2C37-like 基因,大小为1227 bp。由图2可见,条带处于2 000 bp 与1 000 bp 之间,特异性较强,基本符合预期结果,可用于后续实验。将2.5 μL T4 聚合酶处理的PCR 产物和1 μL同样经T4 聚合酶处理的线性化的pET-30a 载体混合,22 ℃孵育20 min 后加入1 μL 25 mM 的EDTA 终止反应,22℃继续孵育20 min。热激转化DH5α,对重组质粒进行双酶切验证(图3),双酶切验证成功的质粒送华大基因进行测序,测序结果正确。
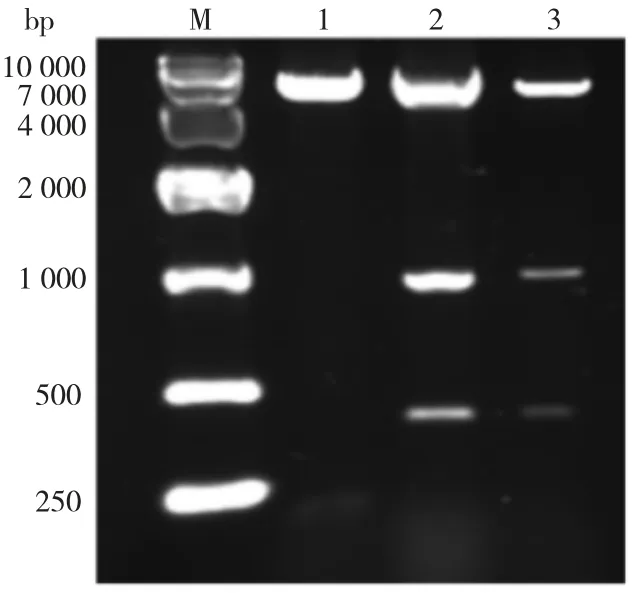
图3 pET-30a-CiPP2C37-like 重组质粒的双酶切鉴定
3.1.3 表达及纯化可溶性CiPP2C37-like 蛋白
为了得到纯度较高的CiPP2C37-like 蛋白,对破碎后的菌体经低温高速离心,取上清进行Ni-NTA柱纯化。CiPP2C37-like 蛋白大小约为44.88 ku,大部分以包涵体的形式存在于菌体沉淀中,但上清中仍存在部分可溶性蛋白能够用于酶活性的测定(图4)。CiPP2C37-like 蛋白在50 mM 和100 mM 咪唑浓度下被洗脱。将50 mM 和100 mM 咪唑洗脱的穿透液进行超滤浓缩,得到粗酶液。
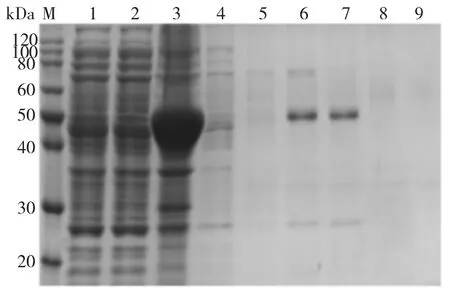
图4 Ni 柱纯化的CiPP2C37-like蛋白SDS-PAGE 电泳
3.2 磷酸盐标准曲线的绘制
为获得用于后续进行酶活力的计算对应OD值下的磷酸盐含量,我们绘制了磷酸盐标准曲线,绘制方法均按Ser/Thr Phosphatase Assay System 试剂盒说明书进行。R2=0.997 4>0.995(图5),符合标准曲线的标准,可以用于计算。
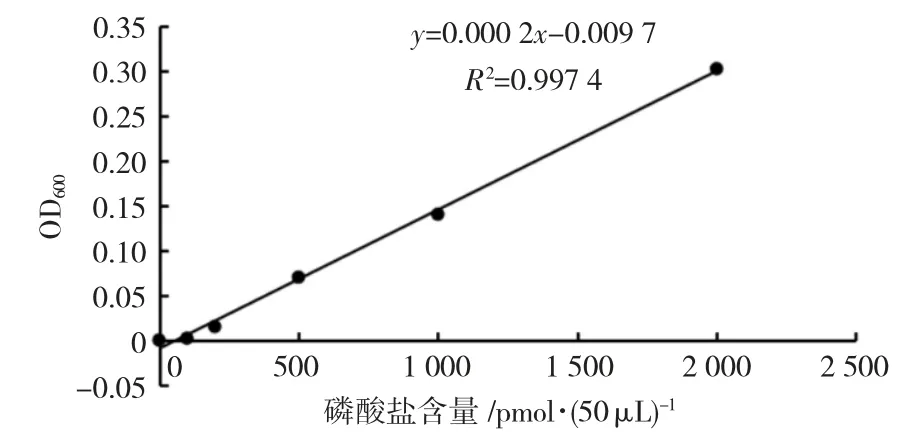
图5 磷酸盐标准曲线
3.3 CiPP2C37-like 磷酸酶活性对温度的响应
通过不同温度对应的吸光度利用磷酸盐标准曲线计算酶活力用于绘制酶活温度曲线。在20 ℃到40 ℃之间,随着温度的增高,酶活性逐渐增强;40 ℃之后,随着温度的升高酶活性逐渐下降(图6)。表明CiPP2C37-like 反应的最适温度为40 ℃左右。
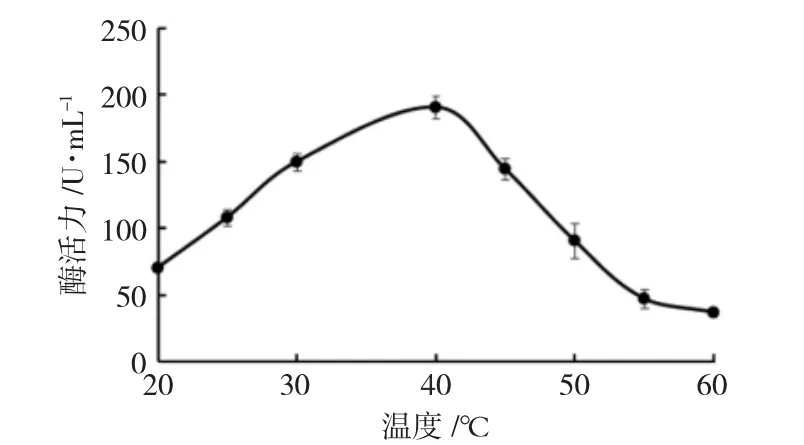
图6 温度对酶活力的影响
3.4 CiPP2C37-like 磷酸酶为碱性磷酸酶
当反应体系的pH 值小于7.5 时,CiPP2C37-like磷酸酶活性较弱,当反应体系pH 值高于7.5 时,酶活性缓慢下降(图7)。综上,CiPP2C37-like 磷酸酶活性在碱性条件下较强,其最适pH 值为7.5,属于碱性磷酸酶。
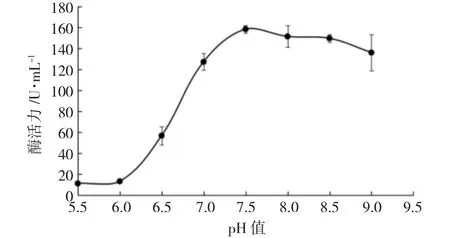
图7 pH 值对酶活力的影响
3.5 CiPP2C37-like 磷酸酶活性对金属离子的响应
以不含金属离子的酶活反应体系为对照,发现Li+和Ba2+对CiPP2C37-like 磷酸酶的活性起到了较为明显的促进作用,K+和Na+对其活性的影响不明显。当反应体系中加入Cu2+时,酶活性被极大的抑制(图8),推测原因可能是Cu2+与酶的活性中心结合,改变了酶的构象,影响了酶与底物的结合能力,降低了磷酸盐的生成速率,导致酶活性降低。
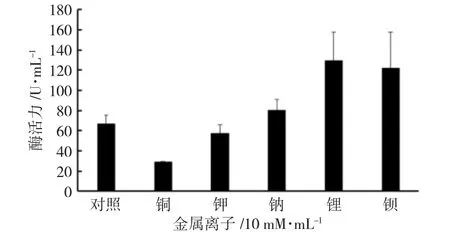
图8 金属离子对酶活的影响
3.6 CiPP2C37-like 蛋白与磷酸肽的亲和性
通过双倒数曲线作图计算CiPP2C37-like 蛋白与人工合成磷酸肽(随试剂盒提供)的Km值。根据拟合趋势线y=0.000 4x+0.0005,算得Km值为0.8(图9)。
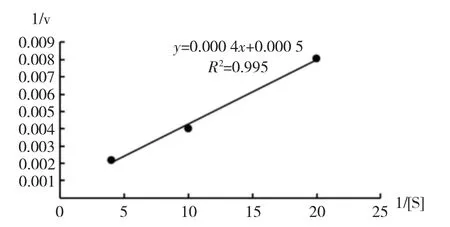
图9 Km 值的双倒数曲线图
3.7 CiPP2C37-like 与拟南芥中PP2CA 亲缘关系
根据NCBI 查询模式植物拟南芥的PP2C 基因,选择了10 个已知功能的基因进行了系统进化树分析,发现CiPP2C37-like 基因序列与拟南芥中PP2CA 亲缘关系最近(图10)。已有研究发现PP2CA在模式植物拟南芥中负调控ABA 信号通路[21-23],故预测CiPP2C37-like 蛋白可能在中间锦鸡儿的ABA 信号通路中起作用。
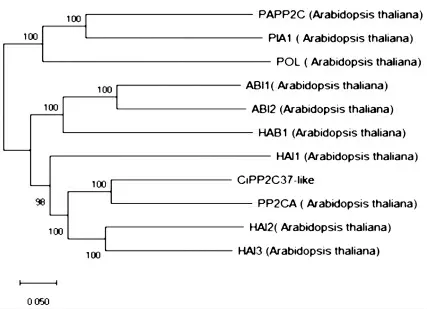
图10 CiPP2C37-like 蛋白与拟南芥中PP2C 的系统进化分析
4 结论
利用原核表达系统,经过镍柱纯化得到了可溶的、有酶活性的CiPP2C37-like 磷酸酶。确定了CiPP2C37-like 磷酸酶与底物反应的最适温度为40 ℃左右,最适pH 值为7.5。Li+和Ba2+能够明显增强酶活性,Cu2+对酶活性具有抑制作用,其对丝苏氨酸磷酸酶测定系统中的磷酸肽的Km值为0.8。通过系统进化树分析,发现CiPP2C37-like 在所有已知功能的PP2C 相关基因中与PP2CA 的亲缘关系最近,在拟南芥中已揭示部分PP2CA 在ABA信号途径中的作用机理,PP2CA 的缺失导致拟南芥种子萌发中ABA 高度敏感,抑制ABA 调控的气孔闭合[22],通过ABA 的受体抑制PP2CA 的蛋白活性和E3 连接酶调控PP2CA 降解,进而调控ABA 相关基因表达[23],故预测CiPP2C37-like 蛋白可能在ABA 信号通路中起负调节作用,为日后CiPP2C37-like 蛋白质在中间锦鸡儿中的作用机理研究提供参考。
