一份矮生玉米株高主基因+多基因遗传模式分析
2022-08-04吴元奇邱贵兰李若楠张艳茹
吴元奇,唐 兰,邱贵兰,李若楠,王 伟,赵 丽,张艳茹
(四川农业大学玉米研究所,成都 611130)
玉米是我国也是世界上重要的粮食作物之一[1],随着21世纪土地面积的不断减少,对玉米单位面积产量的要求也不断提高。以美国为代表的发达国家的生产研究表明,通过提高单位面积的种植密度是提高玉米产量的有效途径。株高是农艺性状中制约玉米种植密度最重要的因素之一[2],并且还影响到玉米的抗倒伏性和收获指数[3]。此外,节间长度和节间数目作为影响株高的关键因素,也将影响植物的结构。矮秆玉米主要通过缩短节间长度和减少节间数目来实现,同时矮秆玉米茎节间强劲,不易折断[4]。在“绿色革命”中半矮秆基因和矮秆基因的发现,使得粮食产量有了显著的提高[5],尤其是半矮秆基因SD1和矮秆基因Rht的引入,分别极大地提高了水稻和小麦的产量[6],矮秆基因逐渐也在玉米、大麦和高粱等[7-11]被大量利用,显著提高了世界谷类作物的产量,植株矮化成了作物育种上最有利用价值的性状之一。近年来,与作物株高或节间长度相关的基因和数量性状基因座(QTLs)不断被报道和表征,并日益展现出来巨大的应用潜力。
株高是玉米的农艺性状之一由主基因+多基因控制,有学者认为主基因+多基因应包含在数量遗传之内[12]。联合分离分析(JSA)是一种通过同时使用多代表型信息来拟合分离模型的最大似然方法[13],混合主基因+多基因遗传模型是以Elston的一个主基因+多基因遗传理论为基础予以推导出来的[14]。这个理论解释了一些经典孟德尔遗传学无法解释的数量性状遗传。主基因+多基因模型已经广泛应用于水稻、玉米和大豆等农作物,但主基因+多基因模型在矮秆玉米的研究中较少[15]。本文试图通过对我们新发现的一个矮秆白玉米株高的6个世代的联合分析,解析矮秆株高性状的遗传模型,估算其遗传效应,为研究其矮秆基因奠定基础。
1 材料和方法
1.1 试验材料与田间设计
本试验被测种为白玉米矮生材料8227,该材料是将矮秆四不像和普通白玉米杂交,再与普通白玉米经过多次回交并自交获得;该自交系遗传稳定且无不良连锁遗传性状,田间综合表现良好,具有较大利用价值。测验种为2份中秆自交系8009和8087,以上材料均由四川农业大学玉米研究所提供。
试验于2017年在四川雅安濆江农场试验田进行,春播种植8009/8227(组合Ⅰ)、8227/8009(组合Ⅱ)、8087/8227(组合Ⅲ)和8227/8087(组合Ⅳ)的亲本P1、P2,并组配F1;2017年秋在云南播种其F1及及其亲本,正反交组合的F1和各自的亲本回交分别得到 B1(F1×P1),B2(F1×P2),杂交种 F1自交得到 F2,至此8227×8009、8009×8227、8227×8087、8087×8227组合的6世代(P1、P2、F1、B1、B2和F2)均已构建好。
2018年春,将上述4个杂交组合的P1、P2、F1、B1、B2和F2分别种植于新津、雅安和自贡3个环境。3个地点全部采用随机区组试验,行长3.5 m,行距80 cm,株距50 cm,每行7窝,每窝2株,种植密度约50 000株/hm2。P1、P2和F1均2行区种植;B1、B2和F2分别为4、4和7行区种植。新津点因试验用地缺少设置2次重复外,雅安、自贡2点均设置3次重复,常规田间管理。
1.2 取样调查
3个地点(新津、雅安和自贡)的各组合的株高于灌浆期进行调查,测量株高时边行的单株舍弃不测量(避免边际效应),选取中间生长正常的植株进行调查;各个组合的P1、P2和F1调查均为10株,B1、B2和F2世代分别调查为30、30和40株。
1.3 统计分析
根据盖钧镒和章元明等[16-17]提出的“六世代主基因+多基因联合的数量性状分离分析方法”,研究玉米正反交组合株高主基因+多基因的遗传规律。包括24类遗传模型:分别为1对主基因(A模型)、2对主基因(B模型)、多基因(C模型)、1对主基因+多基因(D模型)和2对主基因+多基因(E模型)。其分析软件由南京农业大学章元明教授提供。当多基因模型具有上位性效应时,其一阶遗传参数由E.E.Gamble[18]提出的6参数模型计算。
2 结果与分析
2.1 表型分析
白玉米矮生材料8227与2份综合性状优良的自交系8009和8087按Mather & Jinks[19]的方法组配的正反交6世代,考察其在新津、雅安和自贡3个试点的株高性状,白玉米矮秆8227(P1)与中秆8009和8087(P2)组配正交6世代;中秆8009和8087(P1)与白玉米8227(P2)组配反交6世代。如表1-2。
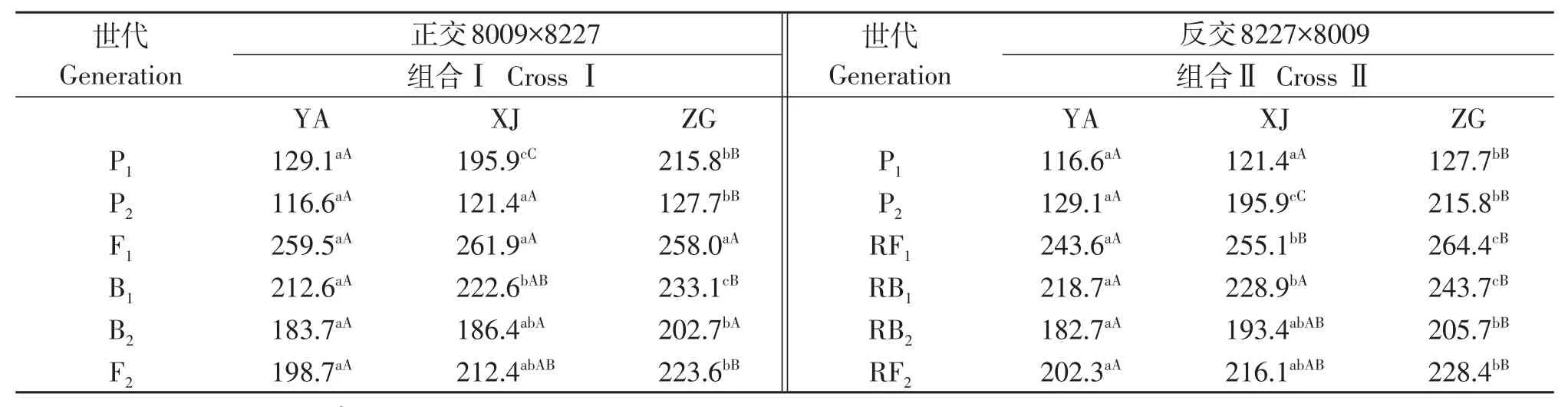
表1 12组合正反交6世代株高平均值Table1 The average plant height of 6 generations in 12 crosses

表2 12组合正反交6世代株高平均值表现Table 2 The average plant height of 6 generations in 12 crosses
从组合I、II的正反交不分离世代P1、P2和F1,可以看出F1、RF1表现明显的杂种优势,P2正反交株高在地点间都表现为极显著差异;就正反交分离世代B1、B2和F2,对于B2、RB2代在雅安、新津2个试点均表现为中亲遗传;RF2、F2分离世代,株高均矮于F1代的株高,表现为自交衰退。
从III、IV的不分离世代正反交P1、P2和F1,可以看出RF1、F1株高表现明显的杂种优势,RF1在3地点株高表现出极显著差异,P1、RP2株高在3个地点都表现出极显著差异,从正反交分离世代B1、B2和F2,可以看出RB1在3地点也表现出极显著差异,F2、RF2株高都矮于F1、RF1的株高,表现为自交衰退,其他分离世代正反交株高较稳定。
2.2 2组合地点间正反交12世代频数分布
从3环境下2组合正反交12世代玉米株高次数分布(图1)可以看出,8227×8009组合和8087×8227组合亲本和杂交后代都表现为单偏态分布,两组合在地点间 B1、RB1都为单偏态分布,B2、RB2在新津、雅安和自贡3个点正反交都表现为双峰偏态分布,8227×8009组合的F2、RF2在新津、自贡和雅安地点间都基本表现为双峰偏态分布,8087×8227在F2、RF2在新津的双峰表现的双峰偏态分布不明显,在雅安和自贡表现的双峰偏态较明显。
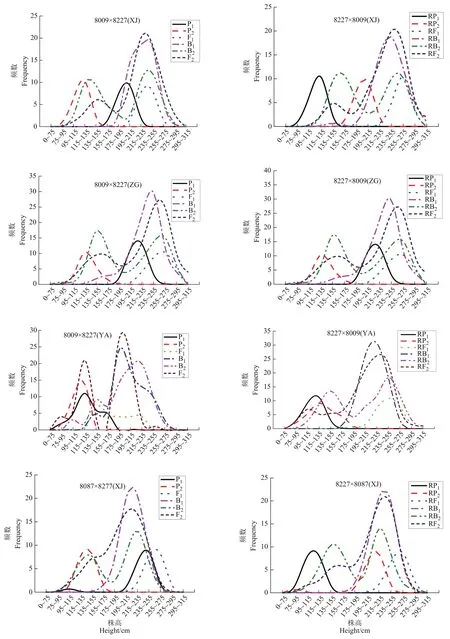
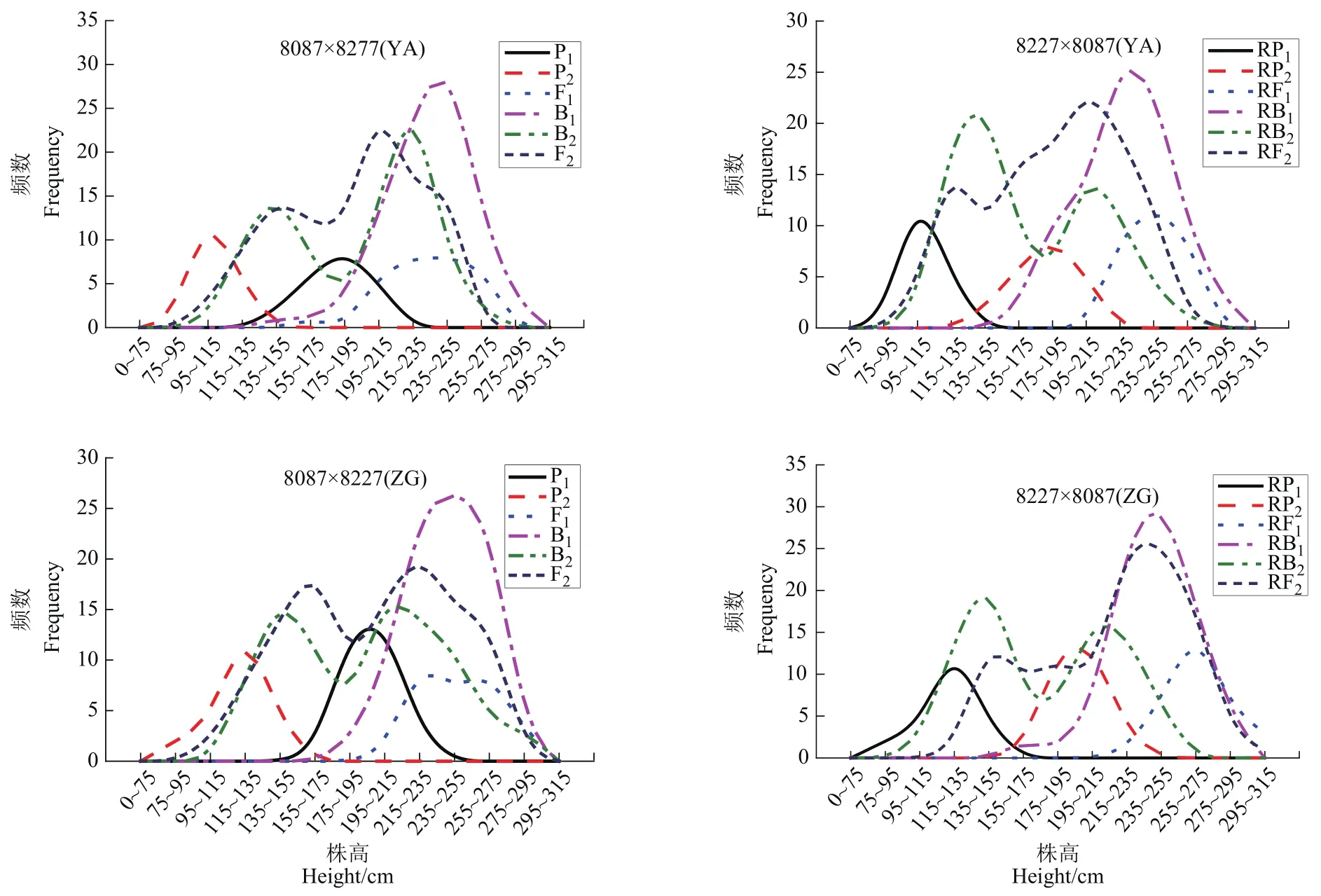
图1 12组合正反交6世代株高频数分布Figure 1 The frequency distribution of plant height in 6 generations of 12 combinations
2.1.3 株高的最适模型
从表3可以看出,于雅安点,组合8227×8087和8227×8009 AIC最小值其模型均为E-1,其余两个组合8009×8227和8087×8227的AIC最小值的模型均为D模型。在新津点,组合8227×8009和8227×8087的AIC最小值均为D模型,其他两个组合8009×8227和8087×8227的AIC最小值模型分别E-1和D-3。在自贡点组合8227×8087的AIC最小值表现为D模型,8227×8009、8009×8227和8087×8227的AIC最小值分别为E、E-1和E-3模型。

表3 2组合正反交株高遗传模型的适合性检验结果Table 3 The results of suitability test of the genetic model of plant height in 2 crosses
2.1.4 遗传参数的估计
从表5可以看出,在新津组合8227×8009的株高由1对加性-显性主基因与加性-显性-上位性多基因控制,表现为D模型。1对主基因的加性效应值为-36.2呈负向效应,显性效应值为63.1呈正向遗传效应,主基因遗传率在B1世达最大、F2世代次之,分别为91.9%、82.0%。雅安为2对主基因由显性-上位性主基因和加性-显性多基因控制,符合E-1模型。其2对主基因加性效应值均为-18.3呈负效应,2对主基因的显性效应值分别为99.1、99.3呈正向效应。在两对主基因中的上位性中,显性×显性互作效应较大且为负向。主基因遗传率在B1、F2世代较大,分别为88.2%、91.5%。在3地点间多基因遗传效应较小。自贡点受2对主基因-显性-上位性主基因控制,表现为E模型。其2对主基因加性效应值均为-28.1具有负向效应,第1对主基因的显性效应值为48.6具有正向效应,第2对主基因显性效应值为-12.3,具有负向效应,其主基因遗传率在B1、F2世代较大,分别为89.5%、92.8%,在3地间表现出较强的主基因遗传力,且表型方差占比显著大于环境方差。

表4 2组合正反交株高遗传模型的适合性检验结果Table 4 The results of suitability test of the genetic model of plant height in 2 crosses

表5 8227×8009组合正反交株高最优模型遗传参数的估计Table 5 Estimates the genetic parameters×the high optimal model of 8227 and 8009 positive and inverse strains
新津和自贡2点的组合8009×8227其株高的遗传模型都由2对主基因-显性-上位性模型和加性-显性多基因控制,为E-1模型。其两对主基因加性效应值均为20.2具有正向效应,显性效应值分别为-6.0、-44.6呈负向效应;在新津其主基因显性效应较自贡值较大,在新津点其主基因遗传率B2、F2较大,为93.9%、93.0%;而在自贡点其主基因遗传率为89.8%、86.7%。该组合在雅安点受1对加性-显性主基因和加性-显性-上位性多基因控制,与D模型一致。且1对主基因的加性效应值和显性效应值分别为51.2、29.6均呈正向效应,主基因B2、F2遗传率较大,分别为73.4%、85.6%。在3个地点间雅安、自贡的环境方差较大,新津的环境方差占比较小,3地点间相对来说表型方差占主要比值。
由表6可以看出,新津和自贡2点的组合8227×8087其株高遗传模型都受1对加性-显性主基因和加性-显性-上位性多基因控制,与D模型一致。1对主基因在新津和自贡的加性效应值分别为-27.3和-40.7均呈负向遗传效应,其显性效应值分别为52.1和34.9均呈正向效应,新津点的主基因遗传率B1、B2和F2为88.5%、54.3%和74.9%;自贡主基因遗传率B1、B2和F2分别为82.9%、2.6%和71.5%,B2在雅安多基因遗传为53.5%相比另外两个地点遗传效应较高。雅安受2对加性-显性-上位性主基因和加性-显性-上位性多基因控制,为E模型。2对主基因加性效应值均为-16.4呈负向效应,其显性效应值分别为49.9和15.1具有正向效应,主基因遗传率B1、B2和F2为61.4%、1.4%和59.4%,主基因之间的加性×加性互作效应为正值,显性×显性互作效应为负值,环境方差在雅安较高,在其他两地点值较低,表型方差在3地点值都较高。
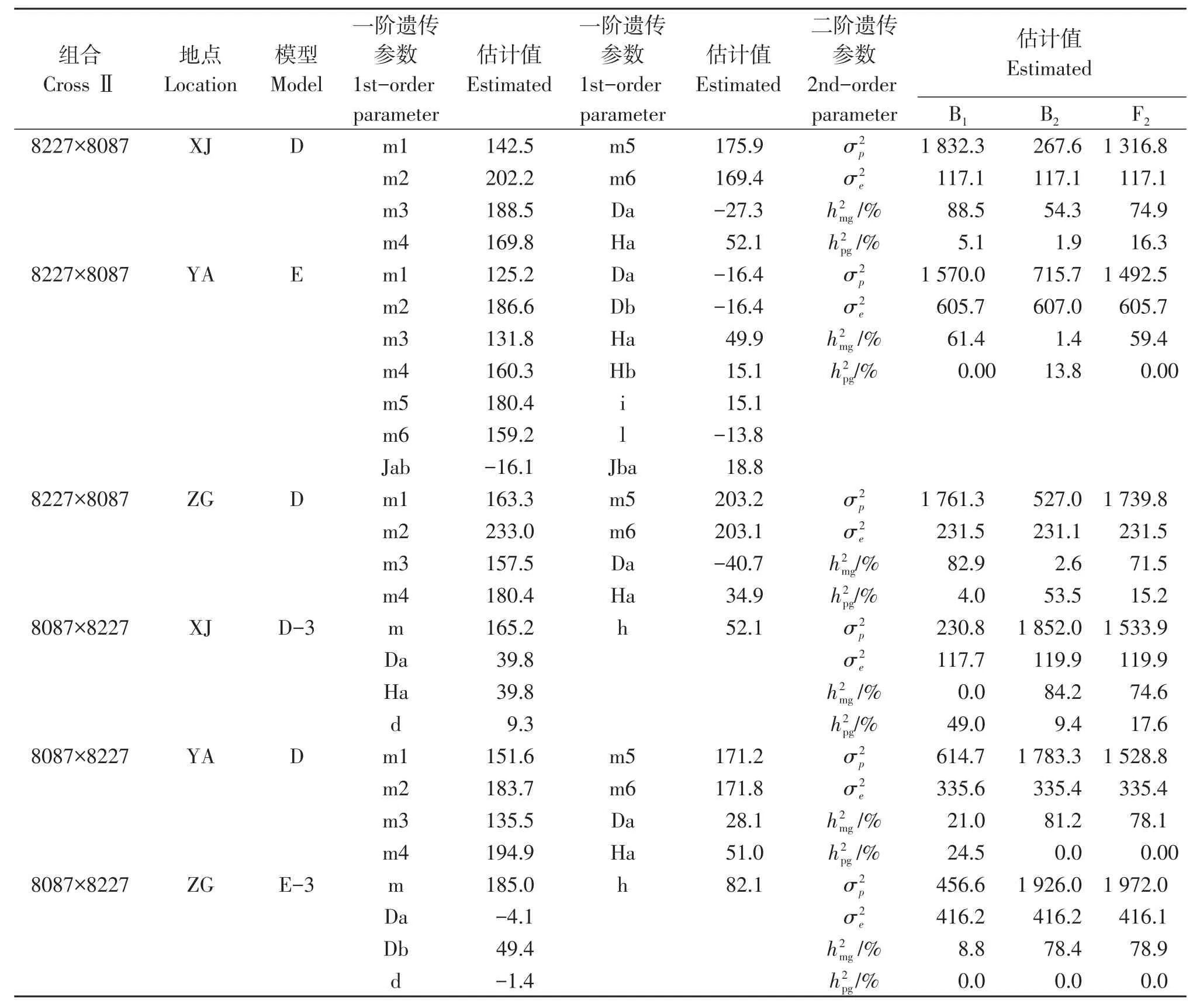
表6 组合8227×8087株高正反交组合最优模型遗传参数的估计Table 6 Estimates of the genetic parameters of the optimal model of the combination 8227×8087 high positive and inverse combinations
组合8087×8227的株高在新津点受1对完全显现主基因和加性-显性多基因控制,与D-3模型一致。主基因加性效应和显性效应累计值均为39.8呈正向效应,主基因遗传率B2、F2较大为84.2%、74.6%,B1以多基因遗传为主为50%。雅安受1对加性-显性主基因和加性-显性-上位性多基因控制,为D模型。1对主基因的加性效应值和显性效应值分别为28.1与51.0呈正向效应,上位性效应中显性×显性互作效应较大为负向,主基因遗传率B1、B2和F2为21.0%、81.2%和78.1%。自贡点受2对加性主基因和加性-显性多基因控制,与E-3模型一致。主基因遗传率B1、B2和F2为8.8%、78.4%和79.0%。3个地点间雅安、自贡的环境方差较大,新津的环境方差值较低。
3 讨论与讨论
3.1 该矮生玉米的株高属于主基因+多基因遗传模型
根据前人不同的试验材料研究表明玉米株高为主基因+多基因控制的数量性状,根据株高遗传模型其最少有一对到两对主效基因+微效多基因控制,不同的多种遗传结构模型可合理地解释群体在玉米高度上的差异。由本试验两组正反交世代样本频数分布研究表明,正反交P1、P2、F1和B1总体表现为单峰偏态分布,而正反交B2和F2总体表现为双峰分布,研究结果表明玉米株高遗传存在1对或2对主基因及微量多基因遗传。
3.2 该矮生玉米的株高遗传可能因遗传背景、环境、组合不同而有所差异
本研究通过将白玉米矮生材料8227和自交系8009、8087组配正反交组合6世代,对1年3点共计12组合的株高进行了遗传分析,结果显示12组合的株高与5种遗传模型一致,D(5个组合);E-1(3个组合);E(2个组合);D-3(1个组合);E-3(1个组合),由此可见,12组合玉米株高的模型不一致,在正反交组合之间,或者相同组合在不同地点中的表现以及不同组合在相同地点中的表现,玉米株高的遗传模型都存在着一定的差异性。J.A.Peiffer[20]运用盖钧镒的主基因+多基因的数量模型在多环境下对株高进行了联合世代分析,研究结果因环境的不同而不尽相同。本研究与许多学者[21-24]的研究发现相一致,株高的遗传表现为主基因+多基因混合遗传。王铁固等[21]研究发现,组合Ⅰ、Ⅱ株高的遗传,春播以主基因为主,夏播则以多基因为主。何文昭等[22]以郑58和HD568组配得到的220个重组自交系,在3个环境下对株高和穗位高的遗传特性进行剖析,对株高而言,在环境Ⅰ(2015年夏,河南新乡)符合F-2遗传模型,而在环境Ⅱ(2016年,夏河南新乡)和环境Ⅲ(2016年春,吉林公主岭)则符合G-0遗传模型;对穗位高,在环境Ⅰ和Ⅱ符合F-2遗传模型,而在环境Ⅲ符合G-0遗传模型;该结果均说明株高和穗位高的遗传模型受环境的影响很大。蒋锋等[24]利用甜玉米为材料的研究中发现株高的最适模型为E-0。许莹莹等[25]利用玉米自交系K12和Fc521研究发现在正交组合中,E-1模型为株高的最适模型,D-2模型为穗位高的最适模型;在反交组合中,株高的最适模型为E-3,穗位高的最适模型为D-2。但本试验与少数研究者[25-26]的结果不相同,他们发现株高遗传规律为多基因控制,不存在主基因效应。赵刚等[26]在研究微胚乳超高油玉米时,对2个组合株高的进行遗传分析,研究表明株高在组合I中的最适模型为C模型。彭静等[27]以玉米自交系092作母本,51作父本,研究发现株高的最适模型为C模型。因此玉米的株高遗传因不同的遗传背景、环境和组合而有所差异。
3.3 该矮生玉米的株高遗传存在有一定的母体效应
组合8227×8009和8009×8227不仅在遗传模型存在一些差异,而且其遗传效应差异还十分明显,从而说明存在一定的母体效应,该组合8227×8009,在新津、雅安和自贡模型为D、E-1和E模型,组合8009×8227在新津、雅安和自贡模型则分别为E-1、D和E-1;组合8227×8009在B1、B2和F2世代不同地点主基因遗传效应值都较大,而8009×227在B1以多基因遗传效应较大,其可能受环境的影响,也存在一定的母体效应,其B2、F2则以主基因遗传为主;8227×8009中,一阶各项遗传参数在自贡、新津均大于8009×8227组合,雅安点相反,说明联合世代分析其亲本不同也会对模型结果产生影响,并且也受一定的环境影响,如果多基因主要集中在母本就会相应的增加多基因的遗传效应,由于其主基因遗传效应值在亲本间遗传效应相对较稳定,因此不会由于亲本不同而受到影响。
对于组合8227×8087和8087×8227不同组合、不同地点之间也表现出模型的差异,在组合8227×8087在地点间符合D、E和D模型,组合8087×8227在3个地点分别符合D-3、D和E-3模型。组合8227×8087遗传参数,3个地点的加性效应都为正值,8087×8227组合则都为负值;就其遗传率在自贡和新津试点而言,对于8227×808组合,B1和F2代的选择效率相对最高;对于8087×8227组合,则以B2和F2代的选择效率最高,该结果表明株高性状受环境影响小,宜在早期世代进行选择,赵延明[23]他认为株高受环境影响大,宜在晚代进行选择,造成两组合选择效率最高世代不一致的原因可能是株高性状受细胞质遗传的影响,综上所述,初步判断该矮生玉米株高在不同组合正反交的遗传效应中存在一定的母质效应。
3.4 主基因加多基因六世代联合的数量性状分离分析方法是探明白玉米矮生自交系株高的可行方法
本研究发现表明株高遗传以主基因为主,多基因为辅,且主基因效应值都比较高,还存在加性和显性效应,显性效应最大效应值为99.32,加性效应最大效应值为-40.71;上位性效应则因组合而异,这可能与试验材料不同有关。这与赵延明[23]的结果相一致,但与霍仕平等[28]的玉米株高以加性效应为主的结果相反。该矮生玉米符合主基因加多基因的联合分离分析方法,探明了矮生玉米的遗传效应,其至少有一对到两对主基因控制,存在微量的多基因遗传,该性状的遗传模式进行分析时,应考虑株高的遗传受试验材料、环境和母体效应的影响,从而做出正确的遗传分析结果。本研究对白玉米矮生新材料8227的株高进行了较为全面的分析,发现该自交系在株型育种上具有较大的应用前景,并掌握了其遗传规律,为后面进一步基因定位和分析鉴定了基础。
