喀斯特地区珍稀濒危铁皮石斛野外回归试验
2022-07-16周玉飞罗晓青王晓敏康专苗张显波
周玉飞 罗晓青 王晓敏 康专苗 张显波


摘要: 野外回歸是珍稀濒危植物多样性保护的主要方式,将珍稀濒危植物铁皮石斛种质扩繁后回归到原生境以实现喀斯特地区铁皮石斛资源多样性保护。根据黔西南州喀斯特地区野生铁皮石斛资源原生境勘察情况,在野生铁皮石斛资源分布地兴义市则戎镇冷洞村开展铁皮石斛野外回归方式和生境条件研究,并监测回归后期铁皮石斛的适应性、生物学特性和生长情况。结果表明,铁皮石斛植于纵向堆砌岩石、植于岩石天然缝隙、植于横向堆砌岩石和捆绑于岩石的存活率分别为95.02%、91.22%、62.57%和37.59%;山脊、山鞍、山顶、山脚和山腰的铁皮石斛存活率分别为93.44%、91.81%、87.01%、84.07%和82.49%;铁皮石斛回归1年后逐渐适应野外生境,在回归2~ 3年内存活率变化不明显,回归4年后存活率明显下降;铁皮石斛回归初期发病率较高,为7.60%,回归1年后发病率变化不明显,回归4~ 5年后铁皮石斛老株多因生境变化出现腐烂现象;铁皮石斛回归初期高位芽萌发率较高,为8.47%,回归1年后高位芽萌发率明显降低,回归3年后基本不萌发高位芽;不同回归方式、生境条件和回归时间对铁皮石斛的分蘖率、新芽高、新芽粗没有明显影响。综上所述,回归方式和生境条件是影响铁皮石斛野外回归成败的主要因素,植于纵向堆砌岩石和岩石天然缝隙的铁皮石斛存活率最高,郁闭度、通风条件、苔藓层等生境因子对铁皮石斛的适应性和生长具有一定影响。
关键词: 铁皮石斛; 珍稀濒危; 喀斯特地区; 野外回归
中图分类号: S181 文献标识码: A 文章编号: 1000-4440(2022)03-0798-08
Re-introduction experiment of rare and endangered Dendrobium officinale in karst area
ZHOU Yu-fei, LUO Xiao-qing, WANG Xiao-min, KANG Zhuan-miao, ZHANG Xian-bo
(Institute of Subtropical Crops, Guizhou Academy of Agricultural Sciences, Xingyi 562400, China)
Abstract: Re-introduction is the main way to protect the diversity of rare and endangered plants. In order to protect the diversity of Dendrobium officinale resources in karst areas, the rare and endangered Dendrobium officinale germplasm was bred and returned to its original habitat. According to the investigation of the original habitat of wild Dendrobium officinale resources in karst area of southwest Guizhou, the re-introdution method and habitat conditions of Dendrobium officinale were studied in Lengdong Village, Zerong Town, Xingyi City, where the wild Dendrobium resources were distributed. And the adaptability, biological characteristics and growth of Dendrobium officinale were monitored at the later stage of re-introdution.The results showed that the survival rates of Dendrobium officinale planted in longitudinal pile rocks, natural crack of rocks, transverse pile rocks and bound to rocks were 95.02%, 91.22%, 62.57% and 37.59%, respectively. Moreover, the survival rates of Dendrobium officinale at ridge, saddle, top, foot and mountainside were 93.44%, 91.81%, 87.01%, 84.07% and 82.49%, respectively. Dendrobium officinale gradually adapted to the habitat after one year of re-introduction, the survival rate did not change significantly after two or three years, but decreased significantly after four years. The incidence rate of Dendrobium officinale was 7.60% at the beginning of re-introduction, did not change significantly after one year, the old Dendrobium officinale was mostly tended to rot due to habitat changes after four or five years. Relatively high germination rate of high bud was 8.47% at the beginning of re-introduction, decreased significantly after one year, and high bud dmost did not germinate after three years. The tiller rate, new bud height and diameter of Dendrobium officinale were not affected by different methods, different habitats conditions and different years. In conclusion, re-introdution methods and habitats conditions are the main factors affecting the re-introduction success or failure of Dendrobium officinale . The survival rate of Dendrobium officinale planted in longitudinal piles rocks and natural crevices of rocks is the highest. The habitat factors such as canopy density, ventilation condition and moss layer have certain effects on the adaptability and growth of Dendrobium officinale .
Key words: Dendrobium officinale ; rare and endangered; karst area; re-introduction
贵州西南部喀斯特地区石斛资源较为丰富,铁皮石斛( Dendrobium officinale Kimura et Migo)因药用价值高遭到灭绝性采挖,野生铁皮石斛资源被严重破坏 [1] 。国际自然保护联盟(International Union for Conservation of Nature,IUCN)已将铁皮石斛列为极度濒危物种。在中国,铁皮石斛属于国家Ⅰ级保护植物 [2] 。珍稀濒危植物是生物多样性保护中优先保护的对象,植物多样性保护的主要方式是就地保护(In-situ conservation)、迁地保护(Ex-situ conservation)和野外回归(Re-introduction) [3] 。野外回归(又称再引入)是将人工繁殖幼苗引种到原本分布的自然或半自然生境中,建立足够的遗传资源来适应进化改变,可自然维持和更新种群 [4] 。它是一项新兴且长期有效的生物多样性保护模式,也是联系珍稀濒危植物就地保护和迁地保护的重要桥梁 [5] 。近年来,越来越多的珍稀濒危植物野外回归保护在实践中得到了应用,然而关于铁皮石斛野外回归研究鲜有报告。本研究在铁皮石斛野生资源原分布范围内将铁皮石斛种苗回归到原生境中,以保护铁皮石斛生物多样性为目的,探索珍稀濒危铁皮石斛野外回归方式和生境条件,监测回归后期铁皮石斛的适应性、生物学特性和生长情况,为进一步研究铁皮石斛野生资源保护和可持续利用奠定基础,也为其他兰科植物野外回归提供参考依据。
1 材料和方法
2014-2015年,对黔西南州野外石斛资源的分布种类、分布数量、生境特点、繁殖情况等资源状况的调查研究结果显示,铁皮石斛野生资源极为稀少,濒临灭绝 [6] 。2016年3月,将铁皮石斛人工繁育种苗回归到野外,回归试验地点选在兴义市则戎镇冷洞村,该地植被丰富多样且有多种野生兰科植物生长,也是铁皮石斛野生资源主要分布区域之一。
1.1 野外回归试验地的自然生态条件
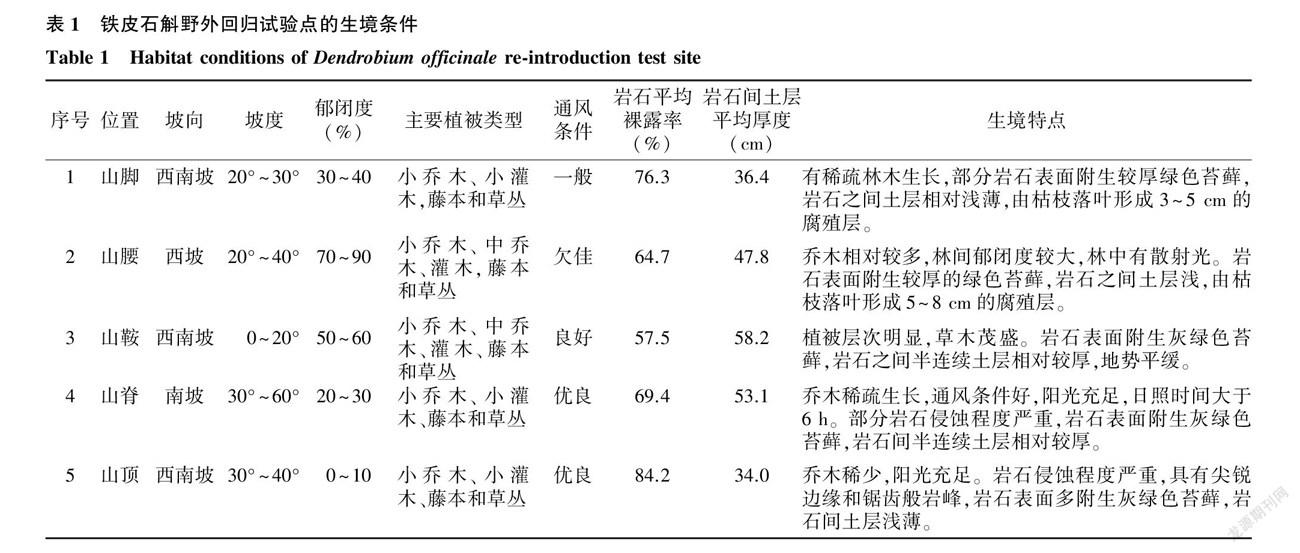
1.1.1 地理位置与气候条件 铁皮石斛野外回归地位于兴义市则戎镇冷洞村(E104°1′~ 104°56′,N24°34′~ 24°54′),海拔1 100~ 1 300 m,属低纬度高海拔区。该地区具有亚热带季风湿润气候特征,热量充足、雨量充沛、雨热同季。年均日照为1 720 h,年平均气温17 ℃,极端最高气温34.9 ℃,极端最低气温-3.1 ℃ ,年平均无霜期340 d,年平均降雨量1 500 mm,80%的雨水都集中在5- 9月份,年蒸发量1 492 mm,年相对湿度81%。
1.1.2 岩石-土壤-植被类型 铁皮石斛野外回归试验地是典型的喀斯特石山区,岩石裸露率为50%~ 80%,岩石上有石生苔藓覆盖。土壤为石灰岩母质发育的石灰土,呈弱碱性,pH 7.1~ 7.9。岩石之间的土层浅薄,土层平均厚度在10~ 60 cm,枯枝落叶腐烂后会形成5~ 20 cm的腐殖层。在植被丰富多样的喀斯特石山林中,常年云雾滋润,林中有散射光,郁闭度0~ 90%,风速1~ 3 m/s 。回归区域坡向为南坡、西南坡和西坡,坡度0°至 60°。植被类型为次生性常绿阔叶落叶混交林,主要有樟科、桑科、大戟科、榆科、漆树科、鼠李科、壳斗科、玄参科、棕櫚科、豆科、楝科、柏科、罗汉松科、苏铁科、清风藤科、蔷薇科、芸香科、禾本科、无患子科、忍冬科、仙人掌科、芭蕉科、兰科、百合科、茄科、五加科、天南星科、石蒜科、景天科、茜草科、苦苣苔科、虎耳草科、蕨科、白蘑科等植物。
1.1.3 回归试验点生境条件 在山脚、山腰、山鞍、山脊、山顶5个试验点种植铁皮石斛,各试验点的坡向、坡度、郁闭度、主要植被类型、通风条件、岩石裸露率、岩石间土层厚度和生境特点见表1。
1.2 试验材料
从黔西南州兴义市、安龙县、册亨县、望谟县等地收集的铁皮石斛种质资源,经组培扩繁并在大棚驯化12个月以上。选择株高≥ 5.0 cm、茎粗≥ 3.5 mm、根≥3 条、叶片≥ 6片的健康无病虫害的合格驯化苗,以3~ 5株为一丛进行野外回归种植。铁皮石斛回归种植数量5 000 丛,除日常管理需要,尽可能减少人为干扰。
1.3 回归生境和回归方式
1.3.1 回归生境 在回归试验区域,通过勘查喀斯特地区山林中的地形、坡向、坡度、郁闭度、土层厚度、植被和岩石裸露等情况,分别在山脚、山腰、山鞍、山脊、山顶共5个试验点设置铁皮石斛野外回归试验。
1.3.2 回归方式 利用天然的喀斯特岩石种植铁皮石斛,主要采取4种回归方式。(1)植于岩石天然缝隙:将铁皮石斛种苗种植于石头表面的自然裂缝中,用树皮将其固定;(2)植于横向堆砌岩石:将铁皮石斛种苗种植于横向堆砌岩石缝隙中,用树皮将其固定;(3)植于纵向堆砌岩石:将铁皮石斛种苗种植于纵向堆砌岩石缝隙中,用树皮将其固定;(4)捆绑于岩石:将铁皮石斛种苗根茎部用麻绳固定和捆绑于岩石四周。
1.4 回归铁皮石斛的适应性和生长调查
调查不同生境条件和不同回归方式下铁皮石斛的适应性和生长情况。
不同生境条件下铁皮石斛的适应性和生长情况:每个试验点选取5个具有代表性的区域,记录铁皮石斛总株数、存活株数、发病株数、萌发高位芽的株数;每个区域随机选取30丛铁皮石斛记录老株数、新芽数、新芽高和新芽粗。不同回归方式下铁皮石斛的适应性和生长情况:选取5个具有代表性的区域,记录总株数、存活株数、发病株数、萌发高位芽的株数;每个区域随机选取30丛铁皮石斛记录老株数(前一年萌发的芽数)、新芽数、新芽高(最长3株新芽的株高平均值)和新芽粗(最长3株新芽的茎粗平均值)。统计存活率、发病率、高位芽萌发率(从茎段萌发高位芽的株数占总株数的百分比)、分蘖率(从根部萌发的新芽数与老株数的比值)、新芽高和新芽粗。
1.5 回归后的铁皮石斛后期监测
2016-2021年,监测野外回归铁皮石斛的适应性、生物学特性和生长情况。观察记录野外回归铁皮石斛对环境的适应情况以及生长周期、开花期、繁殖状况等生物学特性。在铁皮石斛野外回归一年后,从存活率较高的试验点选取10个区域进行监测。每年11月,记录各区域铁皮石斛的存活株数、发病株数、萌发高位芽的株数;每个区域随机选取30丛铁皮石斛记录老株数、新芽数、新芽高和新芽粗。统计存活率、发病率、高位芽萌发率、分蘖率、新芽高和新芽粗。
1.6 数据分析
数据使用DPS 7.05软件进行分析。
2 结果与分析
2.1 不同回归方式下铁皮石斛的适应性和生长情况
2.1.1 不同回归方式下铁皮石斛的适应性 在春季,铁皮石斛萌发新芽并长出新根,根系攀附于岩石表面呈裸露状。植于岩石天然缝隙的回归方式操作简单易行,在喀斯特地区石灰岩表面有天然形成的缝隙,选择不易积水的石缝种植石斛,铁皮石斛定植于石缝后生长的新根附着于岩石表面,大部分回归的铁皮石斛都能较好地适宜这种回归方式。横向堆砌岩石是自然散落形成的岩石堆或早期人工开垦土地时为防止水土流失而人为堆砌的石块,横向堆砌岩石间会有枯枝落叶和流失的土壤填充于石缝,铁皮石斛多数难以存活或不能正常生长,在雨季石斛根系基本腐烂,少数植株从茎段上萌发高位芽并长出气生根。纵向堆砌岩石是早期人工开垦土地时为防止水土流失而人为堆砌的石块,纵向堆砌岩石之间形成的缝隙较多,不易积水且透气性好,铁皮石斛大多生长较好,极少出现发病或死亡的情况。捆绑于岩石的铁皮石斛存活率极低,尤其在阳光照射强的天气,铁皮石斛根系因紧贴岩石表面易被烫伤,雨热同季的气候特点使得植株根系易被感染而腐烂。
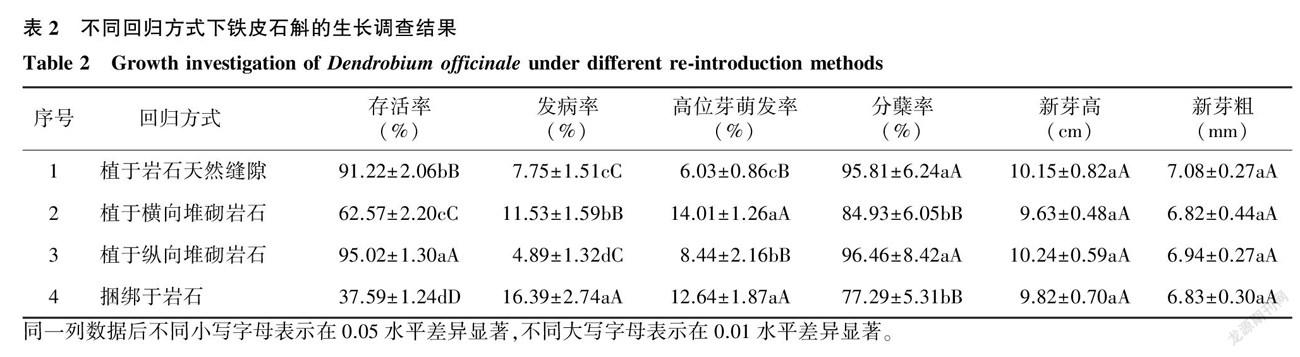
2.1.2 不同回归方式下铁皮石斛的生长情况 由表2可知,回归方式是影响铁皮石斛存活率的主要因素之一。植于岩石天然缝隙和植于纵向堆砌岩石的铁皮石斛存活率最高,分别为91.22%和95.02%;其次是植于横向堆砌岩石的铁皮石斛,存活率为62.57%;捆绑于岩石的铁皮石斛存活率最低,仅为37.59%。从表2可以看出,4种不同回归方式之间铁皮石斛存活率存在极显著差异。其中,病害是影响铁皮石斛存活率的主要原因之一,发病率越高的回归方式铁皮石斛存活率就越低。植于岩石天然缝隙、植于横向堆砌岩石、植于纵向堆砌岩石和捆绑于岩石的铁皮石斛发病率分别为7.75%、11.53%、4.89%和16.39%,4种不同回归方式之间铁皮石斛发病率存在显著差异。高位芽萌发率与植株对环境的适应性相关,植于纵向堆砌岩石的铁皮石斛高位芽萌发率为8.44%,高于植于岩石天然缝隙的铁皮石斛,并存在显著差异;植于横向堆砌岩石和捆绑于岩石的铁皮石斛多因根部腐烂而萌发高位芽,它们的高位芽萌发率分别为14.01%和12.64%,与植于岩石天然缝隙和植于纵向堆砌岩石相比存在极显著差异。植于岩石天然缝隙和植于纵向堆砌岩石的铁皮石斛分蘖率分别为95.81%和96.46%,相比于植于横向堆砌岩石和捆绑于岩石的铁皮石斛存在极显著差异。从铁皮石斛新芽生长情况看,4种回归方式之间铁皮石斛新芽高和新芽粗不存在明显差别。综合铁皮石斛各项生长指标可知,植于岩石天然缝隙和植于纵向堆砌岩石的铁皮石斛存活率、发病率、高位芽萌发率、分蘖率相比于植于横向堆砌岩石和捆绑于岩石的铁皮石斛均存在极显著差异。
2.2 不同回归生境条件下铁皮石斛的适应性和生长情况
2.2.1 不同回归生境条件下铁皮石斛的适应性 根据坡向、郁闭度、植被类型、通风条件和岩石裸露程度等不同生境条件,在喀斯特山林的山脚、山腰、山鞍、山脊、山顶分别布置5个回归试验点,经调查发现,生境条件是影响铁皮石斛适应性的关键因素。5-7月是铁皮石斛野外回归后的快速生长期,这时也正值雨热同期季节。在山脚下,有稀疏的小乔木生长,大部分区域郁闭度较小,铁皮石斛回歸初期阳光直晒植株不易适应。部分附生有较厚的绿色苔藓层的岩石上,铁皮石斛根系多呈水渍状腐烂并逐渐延伸至根茎部,叶片脱落,直至整株腐烂死亡。在山腰,林木相对茂密,通风条件欠佳,岩石表面多附生较厚的绿色苔藓,在高温高湿环境条件下大部分铁皮石斛根茎软化腐烂,而无较厚的苔藓层附生的岩石上,铁皮石斛在回归初期因林间郁闭度大具有较高存活率,但植株生长细嫩。在山鞍,地势平缓,植被丰富且层次分明,林间有散射光,通风条件良好,铁皮石斛生长状况较好。在山脊,有乔木稀疏生长,通风条件好,岩石周边多草丛,阳光充足且光照时间通常在6 h以上,因此铁皮石斛生长状况好,发病率较低。在山顶,乔木稀少,通风条件好,岩石侵蚀程度严重,部分岩石表面附生浅薄灰绿色苔藓,铁皮石斛回归初期阳光直射植株难以适应,有灰绿色苔藓附生的岩石上铁皮石斛大多得以存活并在阳光照射下生长健壮。
2.2.2 不同回归生境条件下铁皮石斛的生长情况 由表3可知,不同生境条件对回归铁皮石斛的存活率和发病率影响较为明显。山鞍和山脊的铁皮石斛存活率最高,分别为91.81%和93.44%,与其他3个试验点相比差异极显著;山顶的存活率仅次于山鞍和山脊,为87.01%;山脚和山腰存活率相对较低,分别为84.07%和82.49%。山脊和山顶由于郁闭度低、通风条件好,发病率最低,分别为3.57%和4.18%,与其他3个试验点相比差异极显著;山鞍通风条件良好但草木茂盛,发病率为7.59%,极显著低于山脚和山腰;山脚和山腰发病率分别为10.82%和12.58%,山腰林间郁闭度大、通风条件欠佳,发病率显著高于山脚。在5个不同的回归生境试验点,铁皮石斛高位芽萌发率、分蘖率差异不显著,高位芽萌发率在8.01%~ 8.87%,分蘖率在88.96%~ 94.19%,铁皮石斛新芽的生长情况总体差异不明显。
2.3 野外回归铁皮石斛后期生长情况监测
2.3.1 野外回归铁皮石斛的适应性 2016-2021年对野外回归铁皮石斛的适应性调查发现,从回归方式看,植于岩石天然缝隙和植于纵向堆砌岩石2种回归方式的铁皮石斛存活率较高,植于横向堆砌岩石和捆绑于岩石的铁皮石斛大部分已死亡。从生境条件看,山脊和山顶的铁皮石斛生长势较好,存活率较高,山鞍的铁皮石斛存活率次之,山脚和山腰的铁皮石斛大部分已死亡。2016-2017年是野外回归铁皮石斛的生长适应期,存活率和发病率受回归方式和回归生境影响最大;2018-2019年是野外回归铁皮石斛的生长稳定期,大部分铁皮石斛基本适应野外环境,尤其在通风良好、阳光充足的区域,铁皮石斛适应性好、植株生长健壮;2020-2021年野外回归铁皮石斛的生长逐渐衰退,随着生境条件变化,存活率出现明显降低,老株茎根部易出现水浸状腐烂现象。铁皮石斛除了受大生境影响外同样受小生境影响,在同一试验点不同小区域的铁皮石斛适应性存在明显差异。
2.3.2 野外回归铁皮石斛的生物学特性 在喀斯特石山区,回归种植于喀斯特岩石上的铁皮石斛株高4~ 20 cm,茎粗5~ 8 mm。铁皮石斛每年可萌发2次新芽,春芽在3- 5月萌发,萌发率98.7%,秋芽在7- 8月萌发,萌发率9.6%。铁皮石斛回归种植第一年萌发的新茎下会长出新根,新根附生于岩石表面,向四周伸展不易拔起,长度不一,根长2~30 cm。一年生新茎的叶片基本不脱落,茎干生长势旺盛;二年生茎的叶片基本完全脱落,茎不再伸长生长,茎干白色;三年生茎无叶片,白色叶鞘逐渐消失,茎干呈赤裸状;四年生茎、五年生茎和六年生茎表型与三年生茎表型相似,呈无叶和无白色叶鞘包裹的赤裸状,但五年生茎和六年生茎相继出现枯萎死亡。铁皮石斛开花期在5月下旬-6月中旬,盛花期在6月上旬,花朵完全展开后1~2 d是授粉最佳时间,授粉花朵第2 d开始逐渐萎蔫,7 d后子房明显膨大,11-12月果实成熟,自然结果率极低,约0.1%,未发现明确的授粉昆虫。铁皮石斛果实成熟后果皮裂开,成熟种子散落,种子在适宜的湿润环境中可萌发,但极难生长成植株,铁皮石斛主要还是通过无性分蘖繁殖。在野外自然生境条件下,铁皮石斛会遭受更多的病害、虫害、鼠害等,受害部位多在叶片、茎干、果实;主要病害有软腐病、黑斑病、炭疽病、根腐病等;主要害虫有蜗牛、蛞蝓、蚜虫、蝗虫、象甲等;鼠害较为严重且难以防控,它们主要啃食铁皮石斛茎干。
2.3.3 2016-2021年野外回归铁皮石斛的生长情况 由表4可知,2016-2021年野外回归铁皮石斛的存活率、发病率、高位芽萌发率出现明显变化,而分蘖率、新芽高、新芽粗变化不明显。2016年铁皮石斛种苗经大棚驯化后回归种植到野外,存活率为89.70%;铁皮石斛生长一年后于2017年再次萌发新芽,存活率为83.61%;铁皮石斛经历1年的野外生长已基本适应野外自然环境,2017-2019年铁皮石斛存活率变化较小;2020-2021年铁皮石斛存活率出现明显下降,2020年存活率为67.16%,2021年存活率为58.80%。2016年铁皮石斛野外回归初期发生大面积病害,发病率最高,为7.60%;2017-2021年铁皮石斛发病率无显著差异,发病率维持在3.19%~ 4.73%。2016年铁皮石斛因环境不适应,萌发高位芽的概率最高,为8.47%;2017-2021年铁皮石斛逐渐适应生长环境,高位芽萌发率明显降低,尤其是2018年以后萌发高位芽的铁皮石斛极少,2020年和2021年铁皮石斛基本没有萌发高位芽。2016-2021年铁皮石斛的分蘖率、新芽高、新芽粗总体上差异不显著。
3 讨论与结论
3.1 影响铁皮石斛野外回归的主要因素
铁皮石斛回归试验区位于植被丰富的喀斯特石山林,地理位置与气候条件都适宜铁皮石斛生长 [7-8] ,因此回归方式和回归生境是影响铁皮石斛野外回归成败的主要因素。试验点林间郁闭度、通风条件、苔藓层也是影响铁皮石斛存活率和发病率的关键因素,据调查统计,在植被丰富的喀斯特石山林中郁闭度为30%~ 70%时最适宜铁皮石斛存活 [9] ,但部分铁皮石斛在郁闭度小于30%或郁闭度大于70%的生境条件下也有自身的适应机制。在郁闭度小于30%的生境条件下,铁皮石斛回归初期因缺乏水分补给存活率较低,而生长于草丛或表面附生少量苔藓的岩石的铁皮石斛因水分散失慢,雨水来临后又得以存活,存活铁皮石斛在生长期间获得充足阳光,植株生长更为健壮。在郁闭度大于70%的生境条件下,植于岩石缝隙无较厚苔藓层的铁皮石斛在回归初期因阳光照晒时间短而具有较高的存活率,但长期缺乏充足光照的铁皮石斛生长细嫩且难以长期存活。有研究结果表明,在珍稀濒危植物野外回归过程中,有些物种幼苗在回归初期需要林下遮阴,回归后期又需要充足的光照条件,有些物种则相反 [10] 。生境条件是影响铁皮石斛高位芽萌发的主要因素,在不适宜的环境中铁皮石斛多萌发高位芽,铁皮石斛适应环境后极少萌发高位芽甚至不萌发高位芽。而不同回归方式、不同回归生境和不同回归时间对铁皮石斛的分蘖繁殖和新芽萌发影响较小。兰科植物是被子植物中物种最为丰富的,也是濒危的植物类群,其对生态系统变化极为敏感 [11-12] 。铁皮石斛野外回归4~5年后老株茎根部出现水浸状腐烂现象,原因可能是回归试验区生态系统逐渐恢复,林木生长茂盛,林间郁闭度增大,绿色苔藓层快速生长,通风条件也受到一定影响,铁皮石斛所处生境条件变化在一定程度上影响了其适应性。
3.2 铁皮石斛野外回歸的意义和存在的问题
珍稀濒危物种野外回归研究对保护生物资源具有重要的科学意义和实践价值。Wright等 [13] 对濒危蜘蛛兰属( Caladenia )植物的种子萌发、移栽培养、共生萌发、苗圃培养进行了研究,并将蜘蛛兰属植物的种子、休眠块茎或植株重新引入自然栖息地。澳大利亚南部对12种全国性濒危野生兰花种群进行原生境分布点和适生生境调查, 并建立生境数据库和GIS模型,定位其潜在生境及种群分布 [14] 。Reiter等 [15] 通过引种回归成功的兰花与所处生态环境之间的相关分析,确定了兰花重新引种成败受传粉者和菌根2个关键生态因素影响,并认为栖息地被破坏、杂草入侵、草食动物采食、非法采集、传粉者减少、病原体侵染和气候变化是影响兰花生存的关键因素。中国对于濒危植物回归的研究起步较晚,但也取得了一些探索性研究进展 [16] 。刘仲健等 [17] 对中国特有的兰科濒危物种杏黄兜兰进行了保育生态学研究,通过对原产地气候、植被和其他环境因素与杏黄兜兰之间的关系以及杏黄兜兰迁地栽培和繁殖无性后代重返原产地的生物学特征调查发现,杏黄兜兰可以进行迁地保护和迁地繁殖植株回归原产地。周艳等 [18] 将国家Ⅰ级保护植物白花兜兰繁殖培育3年后野外回归到茂兰保护区,白花兜兰成活率随着回归时间延长逐渐降低,但长势渐好。张哲 [19] 对3种蝴蝶兰属植物保育的研究结果表明,应加强对生境植物群落和传粉昆虫的保护,大尖囊蝴蝶兰三级电站种群应加强原生境就地保护和种群监测,海南蝴蝶兰椰子村种群应迁地保护。
植物濒危的主要原因除人类过度采收利用、原生境丧失以外,还有植物自身的生物学特性、遗传多样性、物种进化等限制和影响 [20-21] 。大多数珍稀濒危植物存在生殖障碍或自然繁殖极为困难的问题,如濒危植物凹叶厚朴( Magnolia officinalis )雌雄异熟和异位,单株日开花量少等导致传粉过程受阻,自花授粉和同株异花授粉引起花粉管生长出现障碍 [22] 。极危植物宝华玉兰( Magnolia zenii )和渐危植物天目木兰( Magnolia amoena )的种子成熟脱落后常被动物取食,干燥后萌发率大大降低或难以萌发 [23-24] 。极度濒危植物凹脉金花茶( Camellia impressinervis )由于原生境被破坏,种群数量不断减少导致遗传多样性降低 [25] 。兰科植物花部器官高度特化,雌雄蕊集中于合蕊柱上,雌雄蕊由蕊喙隔离防止自交,对传粉者高度依赖 [26] 。铁皮石斛自然结实率低,种子自然萌发也极为困难,主要通过分蘖繁殖实现种群更新。在自然生境条件下,铁皮石斛幼苗发育及成年植株生长均需共生菌提供营养,是典型的菌根植物 [27] 。虽然目前铁皮石斛种子无菌繁育技术十分成熟,但种苗培育至少需要2年以上,种苗培育时间长、成本高,通过共生萌发技术和共生菌育苗技术研究建立菌-苗共生体系将成为铁皮石斛野外回归研究中的一个重要突破点 [28-30] 。其次,原生境被破坏使得原本自然繁殖率极低的铁皮石斛难以维持自身的种群繁衍。增大种群数量和增加铁皮石斛遗传多样性有助于铁皮石斛形成自我维持种群。为降低回归种群带来的遗传风险,应尽量采用本地种源繁育的种苗 [31] 。此外,铁皮石斛野外回归后期种群监测是一项长期而艰巨的研究工作,包括生理生态适应性、病虫鼠害、遗传风险、种群更新及生物多样性等,这项监测任务需持续到铁皮石斛种群实现正常稳定繁衍 [32] 。
参考文献:
[1] 侯欢欢,安明态,金 勇,等. 基于贵州省兰科植物丰富度与分布特征的保护优先区研究[J]. 中国野生植物资源, 2021, 40(1): 88-94.
[2] CHASE M W, CAMERON K M, FREUDENSTEIN J V, et al. An updated classification of Orchidaceae[J]. Botanical Journal of Linnean Society, 2015, 177(2): 151-174.
[3] REN H, JIAN S G, LIU H X, et al. Advances in the reintroduction of rare and endangered wild plant species[J]. Science China Life Sciences, 2014, 57(6): 603-609.
[4] MAUNDER M. Plant reintroduction: an overview[J]. Biodiversity and Conservation, 1992, 1(3): 51-61.
[5] REN H, ZHANG Q M, LU H F, et al. Wild plant species with extremely small populations require conservation and reintroduction in China[J]. Ambio, 2012, 41(8): 913-917.
[6] 吳明开,刘作易,罗晓青. 贵州珍稀兰科植物[M]. 贵阳:贵州科技出版社, 2014.
[7] ZHANG S B, CHEN W Y, HUANG J L, et al. Orchild species richness along elevational and environmental gradients in Yunnan, China[J]. PLoS One, 2015, 10(11): e142621.
[8] 陈 芳. 基于GIS的贵州铁皮石斛气候适宜性研究[D]. 贵阳: 贵州大学, 2017.
[9] 罗晓青,周玉飞,王晓敏,等. 贵州喀斯特地区铁皮石斛仿野生种植研究[J]. 中药材, 2021, 44(3): 15-19.
[10] MASCHINSKI J, HASKINS K E. Plant reintroduction in a changing climate: promises and perils[M]. Washington: Island Press, 2012.
[11] 周 翔,高江云. 珍稀濒危植物的回归: 理论和实践[J]. 生物多样性, 2011, 19(1): 97-105.
[12] 陈宝玲,宋希强,余文刚,等. 濒危兰科植物再引入技术及其应用[J]. 生态学报, 2010, 30(24): 7055-7063.
[13] WRIGHT M, CROSS R, DIXON K, et al. Propagation and reintroduction of Caladenia [J]. Australian Journal of Botany, 2008, 57(4): 373-387.
[14] QUARMBY J P. Recovery plan for twelveth reatened orchids in the lofty block region of south Australia 2010:ISBN 1921238801 FIS90534 [S]. South Australia: Department for Environment and Natural Resource, 2010.
[15] REITER N, WHITFIELD J, POLLARD G, et al. Orchid re-introductions: an evaluation of success and ecological considerations using key comparative studies from Australia[J]. Plant Ecology, 2016, 217(1): 81-95.
[16] 周志华,金效华. 中国野生植物保护管理的政策、法律制度分析和建议[J]. 生物多样性, 2021, 29(12): 1583-1590.
[17] 刘仲健,刘可为,陈利君,等. 濒危物种杏黄兜兰的保育生态学[J]. 生态学报, 2006, 26(9): 2791-2800.
[18] 周 艳,冯佑鸿,李依蔓,等. 濒危植物白花兜兰野外回归研究[J]. 贵州科学, 2018, 36(5): 10-13.
[19] 张 哲. 海南三种蝴蝶兰属植物的保育生物学研究[D]. 海口:海南大学, 2018.
[20] 刘 锋,杨 異,安明态,等. 贵州佛顶山国家级自然保护区兰科植物多样性及其保护研究[J]. 北方园艺, 2022,1(2): 62-70.
[21] EDWARD O, GUERRANT J, THOMAS N, et al. Reintroduction of rare and endangered plants: common factors, questions and approaches[J]. Australian Journal Botangy, 2007, 55(3): 362-370.
[22] 王 洁. 凹叶厚朴繁育系统研究及其濒危的生殖生物学原因分析[D]. 北京:中国林业科学研究院, 2012.
[23] 王 珊. 极危植物宝华玉兰繁殖生物学特性研究[D]. 南京:南京林业大学, 2020.
[24] 張庆宝. 木兰属植物资源收集和天目木兰种苗特征与抗性研究[D]. 杭州:浙江林学院, 2009.
[25] 赖彦池. 凹脉金花茶的保护遗传学研究[D]. 桂林:广西师范大学, 2018.
[26] 张 哲,任明迅,向文倩,等. 东南亚兰科植物的物种多样性、生活习性及其传粉系统[J]. 广西植物, 2021, 41(10): 1683-1698.
[27] 高 越,郭顺星,邢晓科. 兰科植物种子共生萌发真菌多样性及共生萌发机制研究进展[J]. 菌物学报, 2019, 38(11): 1808-1825.
[28] DING J T, TU H Y, ZANG Z L, et al. Precise control and prediction of the greenhouse growth environment of Dendrobium candidum [J]. Computers and Electronics in Agriculture, 2018, 151(8): 453-459.
[29] CORMICK M, BURNETT R, WHIGHAM D. Protocorm-supporting fungi are retained in roots of mature Tipularia discolor orchids as mycorrhizal fungal diversity increases[J]. Plants, 2021, 10(6): 1-13.
[30] QUENTIN F, LORENE G, SONIA L M, et al. Orchids and their mycorrhizal fungi: an insufficiently explored relationship[J]. Mycorrhiza, 2020, 1(30): 5-22.
[31] BRUNDRETT M C, TEDERSOO L. Evolutionary history of mycorrhizal symbioses and global host plant diversity[J]. New Phytologist, 2018, 220(4): 1108-1115.
[32] RAYBUM A P. Recognition and utilization of positive plant interactions may increase plant reintroduction success[J]. Biological Conservation, 2011, 144(5): 1296.
(责任编辑:陈海霞)
收稿日期:2022-02-28
基金项目:贵州省科技计划项目[黔科合支撑(2020) 4Y011号、黔科合支撑(2021)一般227、黔科合重大专项(2019)3001];贵州省科学技术基金项目[黔科合LH字(2014)7700号]
作者简介:周玉飞(1984-),女,贵州兴义人, 硕士,助理研究员,研究方向为兰科植物保护与利用。(E-mail)411725823@qq.com
