藻类响应缺铁胁迫的光合机制研究进展
2022-07-10徐明昱白天宇王佳悦田立荣
徐明昱,白天宇,王佳悦,田立荣,2
(1河北师范大学生命科学学院分子细胞生物学重点实验室,石家庄 050024;2河北省细胞信号与环境适应协同创新中心,石家庄 050024)
0 引言
铁(Fe)是地壳中含量第四丰富的元素[1],是地球上大多数生物的基本必需元素。尽管地壳中铁元素含量丰富,然而缺铁却是动植物界普遍存在的现象。生物可利用的铁主要是Fe2+,在开放的含氧环境中,Fe2+很快被氧化成Fe3+。在生理pH值下,Fe3+只形成不溶性或难溶性的氢氧化物或羟-水络合物[2],严重降低了铁在生物体内的利用效率。在开阔的海洋区域,特别是高氮和磷营养、低叶绿素(High Nutrient Low Chlorophyll,HNLC)的海洋区域,藻类等浮游植物的生长和营养物质的利用主要受Fe2+含量的限制[3]。已有研究表明,低铁条件([Fe2+]≈1 nmol/L)在现代淡水和海洋环境中普遍存在,开放海域的铁浓度甚至可以降低到0.05 nmol/L以下[4],铁缺乏已成为现代淡水和海洋生态系统中藻类生长的主要限制因素。藻类在生物圈处于食物链较低底层,最终会通过食物链影响人类健康。根据世界卫生组织的数据,全世界有30%的人口因缺铁导致贫血[5]。因此,开展藻类应对铁缺乏的作用机制研究,对于提高海洋初级生产力和改善人类铁营养状况等具有重要意义。
铁是植物体内超过300种酶的辅助因子,在植物的光合作用、呼吸作用、叶绿素生物合成等重要的生理过程中发挥着不可替代的作用[6-10]。铁作为辅因子主要以血红素、非血红素和铁硫簇3种形式存在,使蛋白质能够执行催化、电子转运和清除活性氧等功能,对呼吸作用和光合作用的能量转移至关重要[11]。关于铁在植物体内的分布、吸收、运输、储存、代谢以及铁蛋白结构与功能等已有大量文献报道[12-17],李俊成等[18]、董晓雨等[19]、云少君等[20]、张妮娜等[21]已有综述总结,本研究不再做详细论述。
光合器官是最富铁的细胞系统之一[22-23]。叶片中80%的铁来自于叶绿体,其中60%的铁位于叶绿体的类囊体膜上,20%贮存在叶绿体基质中[24]。大量研究发现,环境缺铁严重破坏藻类的光合电子传递反应,造成光合组分的损失等不利影响。然而,目前关于藻类光合器官应答缺铁胁迫所做出的生理反应方面的综述还鲜有报道。因此,本研究主要以铁缺乏对光合电子传递系统的破坏为基础,重点阐述藻类如何响应缺铁胁迫,包括铁诱导蛋白的产生、光合系统的结构重置等内容,并讨论未来铁缺乏相关的光合作用机制和逆境胁迫研究方向。
1 铁是叶绿素和藻胆素生物合成及光合电子传递反应所必需的辅助因子
首先,铁参与光合生物中重要的捕光色素—叶绿素和藻胆素的生物合成过程。已有研究表明,铁是叶绿素和藻胆素合成过程中众多酶的活化剂。例如,叶绿素和藻胆素的合成前体都是δ-氨基乙酰丙酸(δaminolevulinic acid,ALA),ALA合成酶需要铁氧还原酶进行活化。此外,ALA合成叶绿素过程中的α-氨基乙酰丙酮酸合成酶(ALA synthase)、粪卟啉原氧化脱羧酶(coprpporphyrinogen)的活化,以及镁-原卟啉(Mg protoporphyrin)合成原叶绿素酸酯(protochlorophyllide)等过程均需要铁的参与[25]。缺铁胁迫时叶绿素合成相关酶活性显著被抑制,从而限制了叶绿素的合成。藻胆素的合成是由血红素转化为胆绿素IXa(biliverdin IXa),胆绿素IXa被特定的酶部分还原异构形成藻胆素,这个过程是由还原的铁氧还蛋白驱动的过程[26]。缺铁会影响铁氧还蛋白的活性和血红素的合成,进而影响藻胆素的合成。叶绿素和藻胆素不仅作为光合色素捕获太阳能,而且是藻胆体、PSI和PSII等重要蛋白复合物的结构成分,叶绿素和藻胆素含量的降低将直接导致光合效率的下降。
其次,铁是光合电子传递反应中不可缺少的辅因子,铁主要存在于血红素(heme)或者铁硫簇(4Fe-4S簇和2Fe-2S簇)中。以蓝细菌的光合电子传递反应为例,铁是类囊体膜上光系统II(Photosystem II,PSII)、光系统I(Photosystem I,PSI)、细胞色素b6/f复合物(Cyt b6/f)、细胞色素c6(Cyt c6)和铁氧化还原蛋白(Ferredoxin,Fd)等重要蛋白复合物的辅因子[27](图1)。在整个光合电子传递链中,PSI的铁含量最高,每个PSI核心包含3个4Fe-4S簇,即12个铁原子;蓝藻PSI以核心三聚体复合物形式存在,PSI复合物中铁原子的数量达到36个。因此,铁作为光合作用电子传递链上重要蛋白质复合物的关键元素,铁元素的缺乏将直接影响光合电子传递的顺利进行。
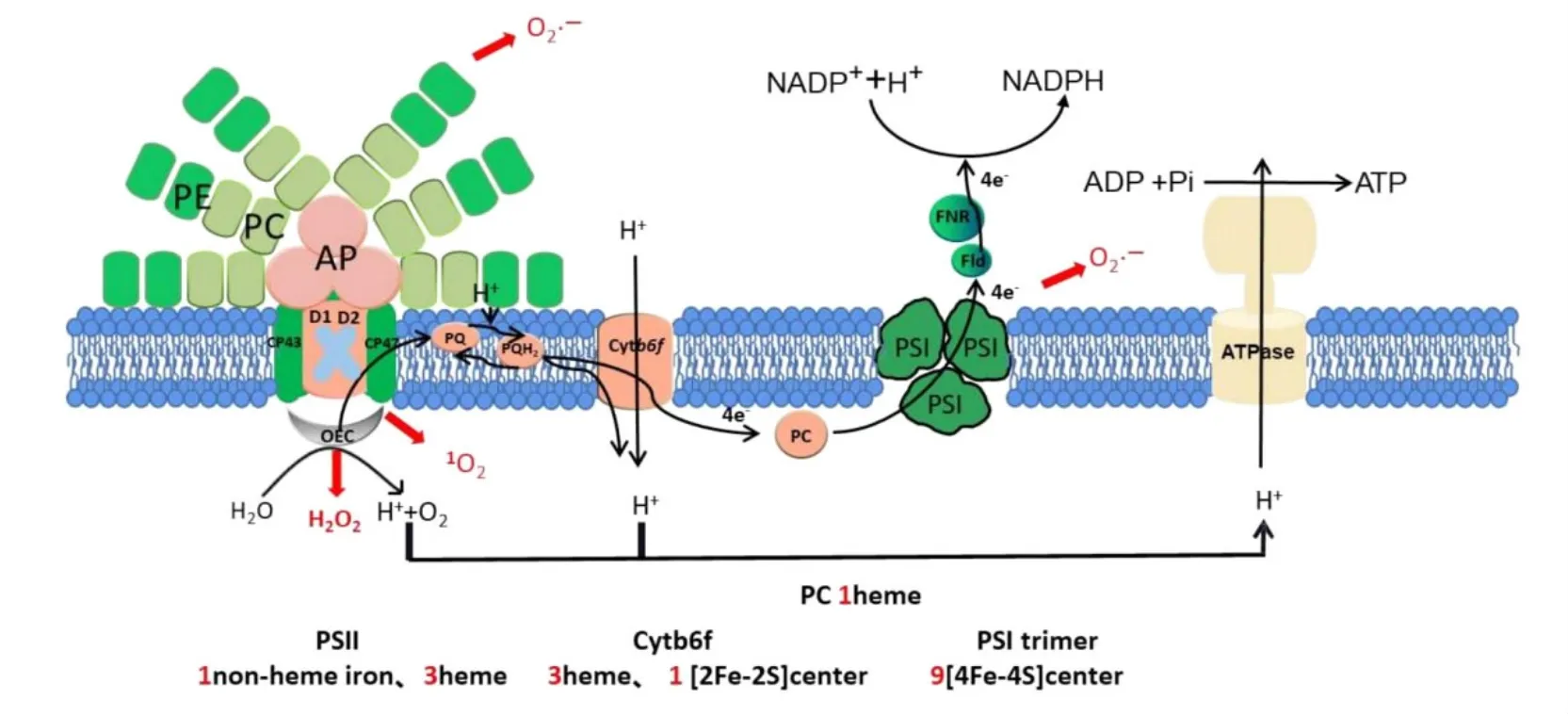
图1 蓝细菌光合电子传递链上Fe原子的存在形式
2 铁缺乏对光合作用的破坏
铁是藻类重要的营养物质,铁元素与光合作用有着密不可分的关系,缺铁会对光合作用产生破坏,使光合作用效率降低,从而导致藻类产量下降。缺铁对光合作用的破坏包括以下几个层次。
2.1 破坏藻类光合元件的形成
铁是光合作用装置中含量丰富的成分,生活在铁营养缺乏的海洋或淡水藻类必须及时调整细胞代谢才能适应低铁环境。藻类在适应缺铁环境时进化出不同的光合响应机制,笔者对国内外已经报道的藻类应对缺铁环境的光合元件进行了汇总(表1),并将蓝细菌和藻类在铁胁迫时光合系统的应答调控进行总结(图2)。可以看出,光合生物处于不同的生态环境时,其应对缺铁环境的光合响应机制不尽相同。例如海洋硅藻(Thalassiosira oceanica)主要通过显著降低细胞内富铁的光合电子传递组分的含量来适应低铁水平,这些组分包括Cyt b6/f和PSI[28]。在陆地和淡水环境中,蓝细菌则通过保持较低的PSI:PSII的比率,以适应低铁的陆生或淡水环境[29]。
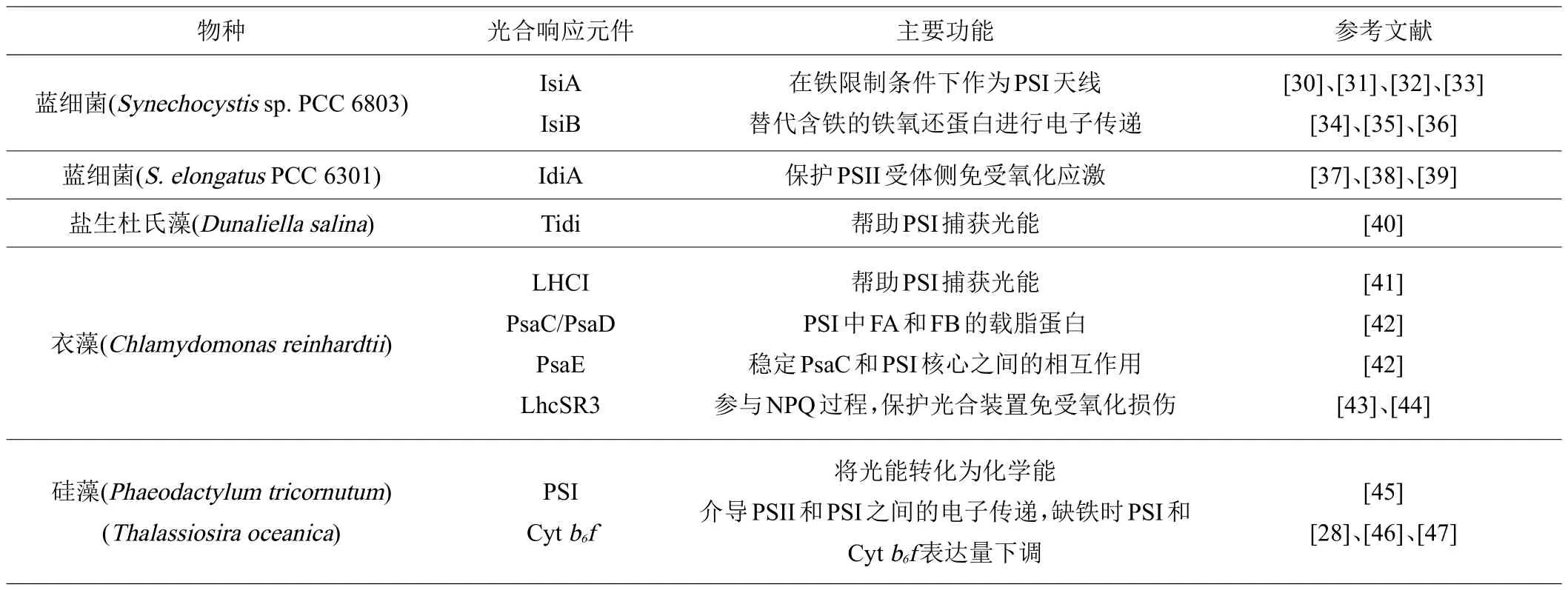
表1 光合生物响应缺铁胁迫相关的光合元件及其功能

图2 藻类光合系统响应铁胁迫的模式图
PSI是整个线性电子传递链中含铁量最高的光合装置,在多个物种中的研究都表明,缺铁对PSI的结构和活性破坏最严重。在莱茵衣藻中,缺铁导致光合器官的重建,除了导致PSI核心亚基的含量显著降低之外,还会造成外周捕光天线的缺失、表达下调或者截短,降低LHCI的结构稳定性,导致LHCI与PSI核心之间的激发能转移效率下降[41-42]。除此之外,在水稻(Oryza sativaL.)中也发现了缺铁对PSI的核心亚基和捕光天线的含量和结构破坏[48]。
除了PSI之外,关于缺铁胁迫对PSII的电子传递活性也进行了相关研究。例如,铁缺乏会造成水分子的氧化和PSII最大量子产率的降低,同时缺铁还会引起PSII的捕光天线LHCII的显著减少,导致PSII活性的下降[49]。同时,在莱茵衣藻中缺铁会诱导产生一种胁迫诱导的捕光天线蛋白LhcSR3(Light Harvesting Complex Stress Related3)。LhcSR3参与莱茵衣藻的非光化学淬灭(NPQ)过程,在缺铁条件下LhcSR3的表达会导致NPQ的增加[50-51]。
2.2 产生氧化应激
PSII和PSI是产生活性氧(Reactive Oxygen Species,ROS)的主要位置(图1),3种活性氧包括超氧阴离子O2·-、单线态氧1O2和过氧化氢H2O2。缺铁导致光合电子传输受损,极大的影响PSII和PSI,从而使其产生大量的ROS。而且,有关ROS解毒酶,如过氧化氢酶、过氧化物酶和一些超氧化物歧化酶均以铁作为辅因子,因此铁限制将导致ROS解毒酶能力的下降,从而加剧氧化应激[52]。同时,在诸多色素蛋白复合物中PSI含铁量最多,因此PSI是受缺铁条件损害最严重的光合结构。藻类缺铁时,PSI及LHCI的蛋白含量、蛋白结构、稳定性均受到影响,P700的光氧化活性显著降低。由于PSI相比其它光合装置(如PSII)受损更严重,由此会造成整个光合线性电子流在PSI处产生瓶颈。
综上,铁作为辅因子为许多蛋白质提供氧化还原电位。缺铁胁迫会引起类囊体膜结构的变化,破坏光合元件的形成,引起光合性能的下降,最终导致植物体活性氧的产生,从而对光合作用产生破坏。
3 藻类应对缺铁胁迫的光合响应机制
由于铁是构成光合电子传递链上各种光合元件的重要金属元素,光合作用装置受到环境中“游离”铁的强烈限制。如上所述,环境缺铁会导致藻类缺绿(叶绿素减少),生长变缓,光合作用效率降低[53]。尽管如此,缺铁并不会造成藻类或植物的直接死亡。这是由于光合生物对铁的强烈需求导致其在适应铁缺乏的过程中,不断调整光合作用过程,以尽量减少缺铁环境对光合作用造成的破坏。
3.1 蓝细菌应对缺铁胁迫的调控机制
蓝细菌提供了地球上初级生产总量的一半以上,大多数蓝细菌生活在海洋和淡水环境中。铁缺乏是水生生态系统中蓝细菌生长的主要限制因素,在缺铁条件下,蓝细菌中PSI和Fd的含量大幅下降。为了适应低铁水平的环境,蓝细菌发展了一套应对缺铁胁迫的调控机制,平衡体内的铁预算和减轻铁消耗,其中最主要的表现是通过降低PSI:PSII比值[54]和表达铁压力诱导蛋白(iron-stress induced proteins A and B,简称IsiA和IsiB)和铁缺失诱导蛋白A(iron deficiency induced proteinA,IdiA)应对铁缺乏。
3.1.1 铁压力诱导蛋白IsiA由IsiA基因编码的IsiA蛋白属于叶绿素结合蛋白,由于IsiA与PSII的内周天线CP43的氨基酸序列同源,因此也称为CP43'[55]。在铁饥饿条件下,不同拷贝数量的IsiA蛋白围绕在PSI核心三聚体周围,形成PSI-IsiA超级复合物,从而扩大PSI的吸收截面,补偿缺铁时的光合效率。已有研究表明,在低铁或者完全缺铁条件下培养蓝藻,PSI周围可以诱导表达不同数量的IsiA蛋白[56-60]。低铁条件或者短时间缺铁时,PSI核心复合物周围会结合18个IsiA蛋白,形成PSI3-IsiA18超级复合物。但是,缺铁时间更长就会影响PSI3-IsiA18的稳定性,导致在PSI三聚体周围形成具有13个IsiA拷贝的单环的复合物[56]。当铁的浓度在纳摩尔每升(nmol)条件下,PSI周围可能产生更多的IsiA蛋白,最多可以达到43个,在PSI核心周围形成两圈环状结构[57]。最近结构生物学家分别利用X-射线晶体衍射和单颗粒冷冻电镜技术解析了完整的PSI3-IsiA18超级复合物的三维结构:PSI-IsiA复合体是一个几乎为圆形的圆盘,具有三重旋转对称性;每个PSI核心三聚体周围有18个独立的IsiA蛋白[59-60],每个IsiA蛋白含有6个跨膜螺旋,结合17个叶绿素(Chls)分子,其中13个Chls的位置与CP43中的Chls是相同的,4个Chls是IsiA所特有的。

图3 蓝细菌PSI3-IsiA18超级复合体的三维结构和IsiA单体结构
目前推测IsiA蛋白的功能主要表现在:IsiA通过在PSI三聚体和单体周围形成多聚体环,充当PSI的辅助天线,增加PSI的吸收截面[61-62];IsiA作为额外的叶绿素储存库[63]:IsiA能够在缺铁期间储存高达50%的叶绿素,一旦环境补充铁,细胞从含铁环境恢复时,叶绿素可以从IsiA复合体中释放出来,用于光系统的重建。因此,IsiA色素-蛋白质复合物不仅在蓝细菌受到铁胁迫时发挥作用,而且在支持光合结构从缺铁状态转变成正常状态的恢复中,发挥关键作用[64];还有研究认为IsiA中含有淬灭的叶绿素分子,可能通过淬灭缺铁细胞不能利用的过量光能提供光保护。然而,时间分辨荧光光谱显示,在PSI-IsiA超复合物中由IsiA到PSI之间存在清晰的激发能量转移,表明IsiA在超复合物中起着能量供体的作用,而不是能量淬灭剂[60]。
总之,IsiA蛋白在帮助蓝细菌适应缺铁胁迫的生存环境时发挥了重要调节作用。但是,IsiA蛋白仅在进化较低等的蓝细菌和原绿藻中存在,在其它真核藻类和植物中并没有发现IsiA的同源基因以及类似PSIIsiA的超级复合物。
3.1.2 铁缺失诱导蛋白A(IdiA)在蓝细菌中铁缺乏时色素-蛋白质复合物的组成发生了重大变化,会产生与PSII相关的铁缺失诱导蛋白IdiA。IdiA蛋白分子量为35 kDa,与二聚体的PSII蛋白复合物结合[66]。研究表明,蓝细菌能正常表达IdiA蛋白。在缺铁条件下,IdiA的表达高度上调[67],可以保护PSII的受体侧免受氧化应激。缺乏IdiA的蓝细菌突变体在铁缺乏下更容易被ROS破坏[68-69]。因而,IdiA在保护光系统II受体侧免受氧化损伤方面发挥重要作用。基因序列比对和蛋白质免疫印迹分析表明,几乎在所有蓝细菌中都存在IdiA的同源蛋白,但是在绿藻或者高等植物中没有检测到IdiA的同源物[47-48]。
3.1.3 替代蛋白 一些藻类应对缺铁环境时可以产生一些特殊蛋白质,替换或保护受缺铁影响的功能蛋白,例如在缺铁条件下,由不含铁的黄素氧还蛋白(flavodoxin,Fld)在功能上取代含铁的铁氧还蛋白Fd。Fld由IsiB基因编码,是以黄素单核苷酸作为辅基的移动电子载体[70]。Fld的氧化还原性质与Fd性质基本匹配,能够在大多数反应中替代Fd[71]。基因组数据分析表明,缺乏Fld的微生物通常位于富含铁的沿海地区,而含有Fld的海洋微生物则位于容易缺铁的海洋环境中[72]。
目前,从蓝细菌解析到的PSI3-IsiA18-Fld3蛋白复合物三维结构中发现,Fld与PSI的结合位点与Fd相同,但是,Fld的尺寸比Fd更大,导致PSI-Fld的亲和力要高于PSI-Fd。与PSI-Fd较小且相对灵活的结合方式相比,PSI-Fld之间的强结合不利于电子传递,因为这需要电子供体与受体之间的快速结合和解离。所以,Fld的效率是低于Fd的[73]。铁缺乏时Fld取代Fd成为电子受体,应对缺铁条件下Fd的含量大幅下降,通过防止光合成分的损失使其在有限的铁条件下维持一定的光合作用,这是蓝细菌和部分藻类应对缺铁环境的一项重要策略。然而,这种补偿机制在陆地植物和衣藻中已经丧失[74]。
3.1.4 环式电子传递 为了优化光合性能,最大限度地减少光氧化损伤,光合生物在不断变化的光照条件和逆境胁迫下进化出一些重要的适应机制,以有效平衡光能吸收与细胞能量需求之间的关系,例如光合电子传递的两种模式之间的转换:线性电子传递(linear electron flow,LEF)和循环电子传递(cyclic electron flow,CEF)。LEF产生ATP和NADPH,而CEF只驱动光合磷酸化提供额外的ATP。CEF具有非常重要的生理意义,不仅能够增加ATP/NADPH比例,提高卡尔文循环的效率;而且,CEF还在光保护中起着重要作用,能够帮助光合生物快速响应环境变化[75]。
由于缺铁胁迫会降低叶绿素的生物合成速度,破坏含铁的色素蛋白复合物(例如PSII和PSI)的结构和功能,尤其PSI受缺铁条件损害最严重,长期缺铁导致LEF产生大量ROS,并导致整个光合线性电子流在PSI处产生瓶颈。在低铁条件下,一些蓝细菌可能利用CEF途径减少氧化损伤以及积累在PSI的电子流。研究发现,在低铁胁迫条件下,聚球藻7942(Synechococcus sp.PCC7942)的LEF受损,聚球藻可以将部分PSII和PSI之间的电子传递解耦联。电子传递主要以围绕PSI的循环模式运行,从而产生质子梯度和ATP,NADP+可能少部分被PSII光还原[76]。在聚球藻8102(Synechococcus WH8102)中并不包含IsiA基因,这使得该生物体可能已经进化出适应缺铁的海洋环境生存的其它机制。相较于需要12个铁原子的PSI,聚球藻具有的质体醌末端氧化酶(plastoquinol terminal oxidase,PTOX)仅需2个铁原子就可以介导CEF,大大降低了光合电子传递对铁的需求[77]。
3.2 杜氏盐藻产生Tidi蛋白
在单细胞绿藻杜氏盐藻(Dunaliella salina)中发现,缺铁诱导产生约45 kDa的铁缺乏诱导蛋白(thylakoid iron deficiency-induced protein),简 称Tidi[40]。Tidi与真核生物中的捕光天线LHCI序列高度相似,属于叶绿素a/b结合蛋白(chlorophyll a/bbinding protein,CAB)家族。生化和光谱数据显示Tidi可以与PSI-LHCI进行功能连接,即Tidi吸收的能量可以传递到PSI的反应中心。因此,Tidi的功能主要是增加缺铁条件下PSI的功能天线尺寸,与蓝细菌中的IsiA蛋白功能相似,二者都是通过扩大PSI的捕光能力帮助藻类适应缺铁的极端环境。Tidi是目前在真核藻类中发现的与铁调控相关的唯一一个CAB蛋白家族成员,在其它真核藻类和植物中还未发现类似Tidi或者新的捕光天线蛋白。
3.3 硅藻适应缺铁胁迫的生存策略
硅藻作为海洋环境中最主要的浮游植物,贡献了地球每年20%的原初生产力[78]。在海洋的广大区域,特别是在高营养低叶绿素(HNLC)地区,硅藻的生长和营养物质的利用主要受铁含量的调节。已有研究发现,硅藻可以在极端缺铁导致大量褪绿的情况下仍能保持光合活性和细胞分裂,说明其光合系统能很好地适应铁的限制条件[79]。研究表明,硅藻可能通过以下不同的生理策略来减少铁需求:(1)通过降低细胞色素浓度以降低光捕获效率[47];(2)通过降低富含铁的PSI和Cyt b6f组分改变光合装置对铁的需求[28,46];(3)利用不含铁的酶替代含铁的酶,如黄酮氧还蛋白[80];(4)硅藻还开发了特定的调节机制,便于在环境应激条件下驱散多余的能量[81-82]。
4 展望
环境中可供藻类吸收利用的有效铁较少,非常低的生物利用率和高的光合需求使铁成为限制藻类生产力的主要微量元素之一。铁作为重要的金属辅因子,与光合作用有着密不可分的联系,充分了解缺铁条件下藻类光合系统的适应机制,对了解藻类逆境生长有着重要意义。在以后的研究中,以下问题值得被关注。
(1)蓝细菌应对缺铁环境的光合作用方式似乎更加灵活,更加多样。蓝细菌在铁胁迫条件下不仅可以诱导表达一些特殊的铁防御蛋白和具有相似生物学功能的替代蛋白,还可以利用一些特殊的光合电子传递机制应对不利的缺铁环境。真核藻类和植物同样面临着环境缺铁的挑战,然而关于真核藻类和植物的光合系统应对环境中缺铁的作用机制目前仍不清楚,值得进一步挖掘。
(2)了解光合生物如何利用铁,以及光合系统如何应对铁缺乏,对于提高海洋藻类生产力和指导农业生产至关重要。过去对藻类缺铁的研究已经揭示了叶绿体中的铁蛋白是如何应对缺铁的环境胁迫;今后应集中研究应对缺铁的反应调控途径,例如在蓝细菌中观察到叶绿体对缺铁的适应很可能受到多重转录、转录后和翻译后机制的调节[82]。缺铁引起的藻类响应逆境的信号网络途径还有待进一步研究,这对揭示光合系统适应铁缺乏的分子机制具有重要促进作用。
