不同夜间温度处理对马铃薯试管薯及块茎形成相关基因表达的影响
2022-07-06李广存段绍光简银巧刘建刚金黎平徐建飞
刘 菊 李广存 段绍光 胡 军 简银巧 刘建刚 金黎平 徐建飞
(中国农业科学院蔬菜花卉研究所/农业农村部薯类作物生物学和遗传育种重点实验室,100081,北京)
马铃薯(Solanum tuberosum L.)是继水稻和小麦之后的世界第三大粮食作物[1],对于保障世界粮食安全和推进中国从脱贫攻坚向乡村振兴平稳过渡具有不可替代的作用。温度是影响马铃薯植株生长和块茎产量的最重要环境因素之一,马铃薯喜冷凉,高温条件不仅会对植株营养生长造成明显的伤害,还会影响块茎形成进程[2]。通常情况下,马铃薯块茎形成的最佳日间温度为14℃~22℃[3],日间环境温度的持续升高将降低或完全抑制块茎形成。研究[4]表明,日间高温频率和强度的不断增加会对作物产量产生不利影响,而夜间高温会对全球作物生产产生更严重的威胁。相比于日间温度,马铃薯块茎形成受夜间温度影响更大[5-6],尤其是夜间低温促进块茎发生,块茎形成的最适夜间温度是17℃[7],夜间温度高于20℃会降低植株结薯数,而夜间温度一旦超过25℃,块茎产量就会急剧下降[8]。研究[9]显示,块茎初始形成期,与日间高温相比,夜间高温通过影响植株的同化产物分配,显著降低了大薯(>100g)比例和早期收获指数,造成产量的严重损失。
有研究[10]表明,马铃薯块茎形成是一个非常复杂的生物学过程,涉及光周期、温度、糖类物质、糖转运体和植物激素等相关基因和通路的协同作用。应用多组学技术研究发现,CO/FT(CONSTANS/FLOWERING LOCUS T)介导的光周期调控块茎形成通路中,一些重要信号因子如StCO1、StGI(GIGANTEA)、StLHY(LATE ELONGATED HYPOCOTYL)、StCDF1(CYCLING DOF FACTOR 1)和StSP5G(SELF-PRUNING 5G)也同时响应温度变化。作为块茎形成的直接诱导者,FT类基因StSP6A同时参与光周期和温度调控块茎形成过程[11-12]。前人研究[12-14]已鉴定出,昼夜节律时钟StTOC1(TIMING OF CAB EXPRESSION 1)、蓝光受体蛋白StFKF1(FLAVIN-BINDING KELCH-REPEAT F-BOX 1)和microRNA SES等信号分子在高温条件下,通过下调StSP6A的表达抑制结薯。然而,关于参与夜间温度调控结薯相关的基因和分子调控机制尚不明确。
为进一步探究夜间温度对马铃薯块茎形成和发育的影响,本试验以马铃薯试管薯为研究对象,通过比较不同夜间温度处理下7个马铃薯品种或材料试管薯的形成情况和结薯时间受夜间温度影响显著材料的块茎形成相关基因表达量的变化,探究最适试管薯形成的夜间温度条件,并为筛选块茎形成调控通路中响应夜间温度变化的基因提供参考。
1 材料与方法
1.1 试验材料
供试材料为中国农业科学院蔬菜花卉研究所马铃薯课题组种质资源库所保存的马铃薯‘DM(DM1-3 516 R44)’、‘RH(RH89-039-16)’、‘中薯3号’、‘中薯5号’、‘中薯18号’、‘中薯21号’和‘费乌瑞它(Favorita)’脱毒试管苗。试管苗繁殖所用培养基为MS固体培养基(30g/L蔗糖和7g/L琼脂粉,pH 5.8),无菌条件下,剪取试管苗单个茎段接种于含50mL培养基的培养瓶(容积350mL,高白料玻璃)中,每瓶接种12株,置于21℃±1℃培养箱中,光照强度2000lx,光周期16h/d条件下培养21d,获得基础苗。
1.2 试管薯诱导和处理条件
无菌条件下,以上述基础苗为母苗,选取长势一致且生长良好植株的第2~3节(从顶芽往下数)单个茎段,接种于结薯培养基(普通MS固体培养基添加蔗糖60g/L和琼脂粉7g/L,pH 5.8),每节段含1个腋芽,每个基因型材料接种30瓶,每瓶接种6个茎节。将接种后的培养瓶分别置于日间/夜间温度23℃/13℃、23℃/18℃和23℃/23℃的人工气候箱(宁波江南仪器厂,型号GXZ-500D)进行试管薯诱导培养,每个基因型单个温度处理下放置10瓶,共60株(20株/重复,重复3次),光照强度均为2000lx,光周期均为8h/d。
1.3 试管薯表型数据记录
培养初期每隔3d观察1次植株生长和试管薯形成情况,生长20d后每天观察记录表型变化和试管薯初始结薯时间,诱导培养60d时统计试管薯结薯率和结薯方式,具体统计方法如下:
试管薯初始结薯时间(d):从接种之日到第1个试管薯初始形成(直径约2mm)的天数[15];试管薯结薯率(%)=(结薯植株数/总植株数)×100;试管薯结薯方式包括3种类型,经由匍匐茎顶端膨大形成的有柄薯(apical microtuber);未经匍匐茎发生,直接在腋芽基部膨大形成的无柄薯(basal microtuber);在植株侧枝腋芽处着生的无柄薯(medial microtuber),分类方法参照周俊[16]和Pantelić等[17](图 1)。

图1 ‘RH’试管薯结薯方式示意图Fig.1 Schematic diagram of types of in vitro tuberization in‘RH’
1.4 RT-qPCR取样和相关基因表达分析
在23℃/23℃处理条件下,取‘RH’试管薯初始形成植株的叶片以及同时期23℃/23℃、23℃/18℃和23℃/13℃尚未有试管薯形成植株叶片,液氮处理后置于-80℃冰箱保存备用,3株叶片混样为1个重复,每个处理重复3次。
按照多糖多酚植物总RNA快速提取试剂盒(GeneBetter)说明书提取RNA,1%琼脂糖凝胶电泳检测RNA完整性,BioDrop uLite超微量蛋白核酸分析仪检测RNA纯度和浓度。获得的RNA用反转录试剂盒Script III RT Kit With gDNA Eraser(GeneBetter)进行cDNA的合成,按照Hieff®qPCR SYBR® Green Master Mix(No Rox)(YEASEN)试剂盒说明进行RT-qPCR。在Light Cycler®480实时荧光定量PCR仪(Roche,瑞士)上完成反应,反应程序为:95℃预变性3min;95℃变性15s,60℃退火15s,72℃延伸20s,40个循环。用Primer 3.0(https://primer3.ut.ee/)设计引物,由北京六合华大基因科技有限公司合成,内参基因EF1α及各基因引物序列见表1。
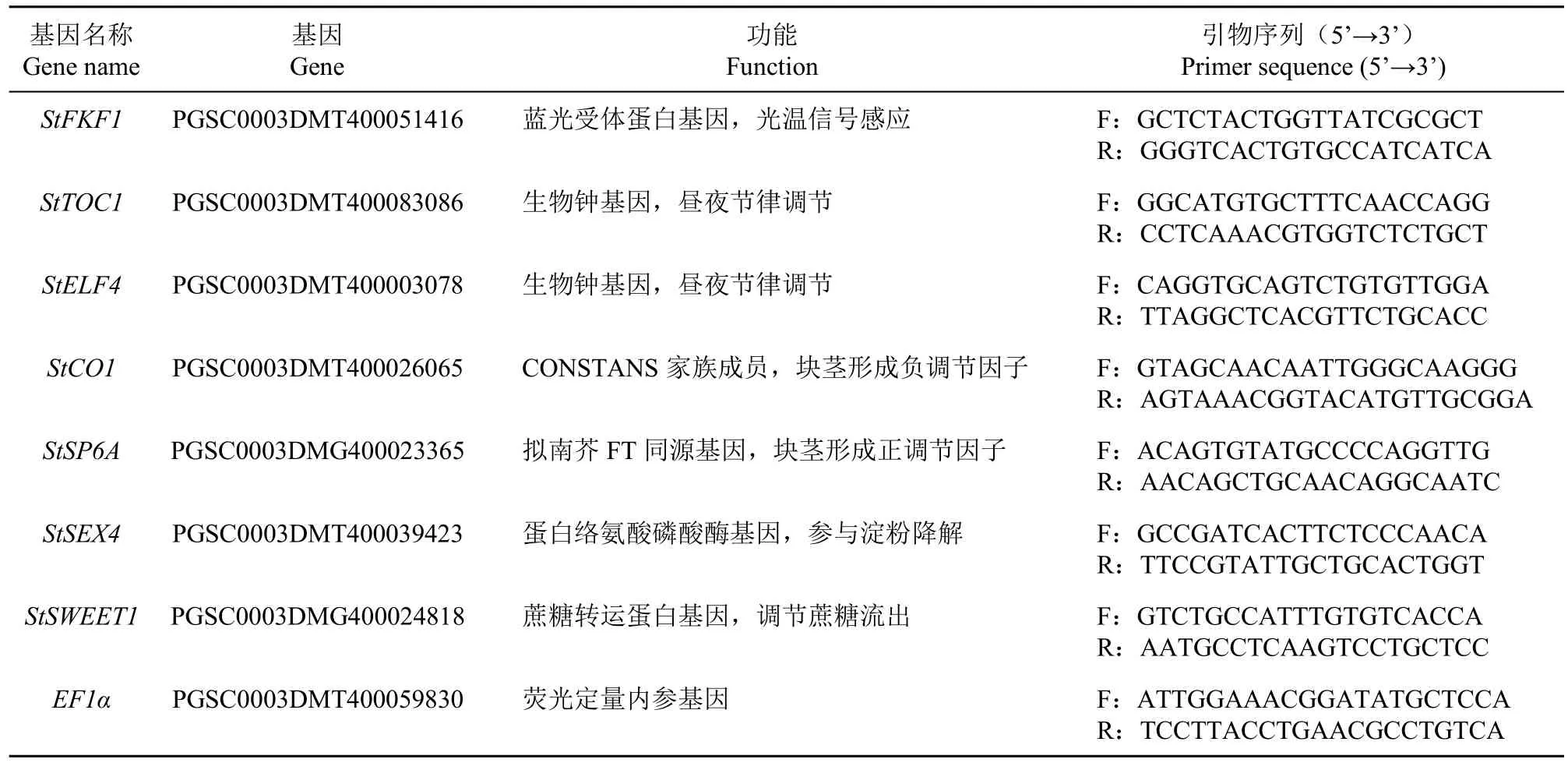
表1 实时定量PCR引物信息Table 1 RT-qPCR primer information
1.5 数据处理
运用Microsoft Excel 2019软件处理数据并作图,利用 SPSS 17.0软件进行单因素方差分析(ANOVA),选用 Duncan’s检验法对显著性差异(P<0.05)进行多重比较,采用 2-ΔΔCT法计算目的基因相对表达量。
2 结果与分析
2.1 不同夜间温度处理对试管薯初始结薯时间的影响
供试材料试管薯初始结薯时间均受夜间温度调控。3种温度处理下,‘中薯5号’、‘中薯18号’和‘中薯21号’试管薯在23℃/18℃下结薯最早,而‘RH’、‘DM’、‘费乌瑞它’和‘中薯3号’在23℃/23℃下试管薯初始结薯时间最短。除‘费乌瑞它’和‘中薯5号’外,其余基因型试管薯在23℃/23℃和23℃/18℃下的初始结薯时间均显著早于23℃/13℃处理(图2)。对于‘RH’和‘DM’基因型,不同夜间温度处理下的试管薯初始结薯时间差异显著。以上结果表明,块茎初始结薯时间受夜间温度影响,过低的夜间温度不利于块茎形成。

图2 不同夜间温度处理对试管薯初始结薯时间的影响Fig.2 Effects on initiation of in vitro tuberization under different night temperature treatments
2.2 不同夜间温度处理对试管薯结薯率的影响
不同夜间温度处理下,供试材料试管薯的结薯能力不同(图3)。随夜间温度升高,‘费乌瑞它’、‘中薯5号’、‘中薯18号’和‘中薯21号’试管薯结薯率呈先上升后下降的趋势,在23℃/18℃条件下结薯率最高。‘RH’和‘中薯3号’试管薯结薯率变化趋势基本一致,即23℃/18℃和23℃/13℃条件下的试管薯结薯率显著高于23℃/23℃处理,本试验中,‘DM’基因型在3种夜间温度处理下的试管薯结薯率无显著差异。以上结果表明,夜间高温处理会降低试管薯结薯率。
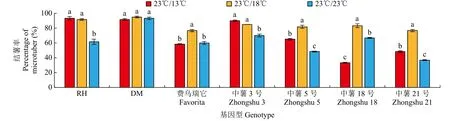
图3 不同夜间温度处理对试管薯结薯率的影响Fig.3 Effects on percentage of in vitro tuberization under different night temperature treatments
2.3 不同夜间温度处理对试管薯结薯方式的影响
在3种夜间温度处理下,大多数供试材料试管薯的结薯方式固定,试管薯多以有柄薯的形式存在,而‘RH’和‘中薯18号’试管薯结薯方式受夜间温度影响而发生变化(图4),随着夜间温度的升高,无柄薯的比例逐渐增加,其中‘中薯18号’在23℃/23℃处理下,存在较高比例基生无柄薯,这表明‘RH’和‘中薯18号’植株在夜间温度升高环境下更倾向于不经由匍匐茎伸长而直接在茎节腋芽处膨大形成试管薯。
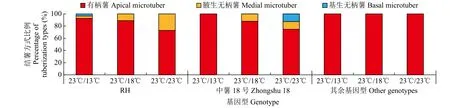
图4 不同夜间温度处理对试管薯结薯方式的影响Fig.4 Effects on type of in vitro tuberization under different night temperature treatments
2.4 不同夜间温度处理下块茎形成相关基因的表达分析
选取参与马铃薯块茎形成的光受体基因(StFKF1)、生物钟相关基因(StELF4和StTOC1)、CO/FT光周期调控通路关键基因(StCO1和StSP6A)以及糖代谢和糖转运体调控相关基因(StSWEET1和StSEX4)进行不同夜间温度条件下结薯植株和非结薯植株的表达分析(图5)。

图5 不同夜间温度处理下块茎形成相关基因表达量的分析Fig.5 Expression analysis of tuberization related genes under different day/night temperature treatments
以结薯最早的23℃/23℃处理下结薯植株叶片中基因表达水平为对照,结果表明,3种温度处理下的非结薯植株与对照相比,StFKF1在23℃/18℃和23℃/13℃处理下的表达量显著上升,而StSEX4在23℃/18℃和23℃/13℃处理下的表达量显著下降。StTOC1和StCO1仅在23℃/13℃处理下的表达量显著升高,其他处理与对照无显著差异。StELF4在3种温度条件下均显著下调表达,与之相反,StSP6A和StSWEET1的表达均显著上调。
通过对3种夜间温度处理下的非结薯植株叶片基因表达变化分析发现,随着夜间温度降低,StFKF1表达量显著增加,23℃/13℃条件下最高;StELF4表达量在23℃/18℃和23℃/13℃之间差异显著;StCO1表达量在23℃/23℃和23℃/13℃条件下差异显著;StSEX4表达量在23℃/23℃和23℃/18℃条件下差异显著;StTOC1和StSP6A表达量的显著性变化趋势相同,23℃/23℃和23℃/18℃处理间无显著差异,但均显著低于23℃/13℃处理;StSWEET1在23℃/23℃条件下表达量显著低于其他2种温度处理。
3 讨论
3.1 不同夜间温度处理对试管薯初始结薯时间和结薯率的影响
温度是影响马铃薯块茎形成和发育的重要环境因子,与日间温度相比,夜间温度对马铃薯的生长和产量建成有更重要的影响[2]。夜间温度尤其是夜间高温对水稻和小麦植株农艺性状和产量等均有不利影响[18-19]。马铃薯试管薯研究表明,环境温度过高或过低都不利于块茎形成,15℃~18℃最适宜试管薯的诱导培养[20],适度低温(17±1)℃培养的试管薯始薯期变短,结薯数增多[21]。本试验中,23℃/18℃处理下绝大多数供试基因型试管薯初始结薯时间显著早于23℃/13℃处理,且23℃/18℃下的试管薯结薯率较高,该结果与张武等[22]和邱甜等[23]研究结果一致。前人研究[24]发现,在诱导结薯时不同马铃薯品种对温度的适应性不同,炸片加工品种‘大西洋’和彩色薯肉品种‘转心乌’试管薯的最佳诱导温度分别为20℃和25℃。本试验中,‘RH’和‘DM’等材料试管薯在23℃/23℃下结薯最早,而‘中薯5号’和‘中薯18号’等在23℃/18℃下结薯最早,表明不同基因型马铃薯试管薯形成对夜间温度的敏感性不同。由此可见,夜间温度对马铃薯试管薯形成存在显著影响,适宜的夜间温度可以缩短块茎初始形成时间和提高结薯率,过高或过低的夜间温度都不利于试管薯形成。
3.2 不同夜间温度处理对块茎形成相关基因表达的影响
温度会对昼夜节律时钟基因的转录水平产生影响。拟南芥中,环境温度影响生物钟基因TOC1、GI、CCA1和LHY等的表达[25]。对马铃薯的研究表明,与22℃相比,30℃条件下叶片中StTOC1表达升高[26],StTOC1是块茎形成的负调节因子,下调StTOC1的表达可以减轻高温对马铃薯产量的不利影响[13]。本试验中,夜间23℃和18℃处理下,结薯与非结薯植株间叶片StTOC1表达量无显著差异,但夜间13℃时,非结薯植株叶片StTOC1表达水平显著升高,表明该温度下,StTOC1显著抑制块茎形成,这与夜间13℃下试管薯初始结薯时间最晚的表型结果相符。高气温环境下,马铃薯叶片生物钟基因StELF4表达量降低[14]。本试验结果表明,在非结薯植株中,夜间低温能够降低StELF4的表达量。
温度会影响糖代谢相关基因的表达。研究[14]发现,与22℃/20℃处理相比,29℃/27℃处理下的块茎中蔗糖外排载体蛋白StSWEET1和块茎中淀粉降解相关基因StSEX4的表达量均显著降低,阻止了蔗糖向质外体泄漏,减少了高温下块茎中的淀粉含量损失。马铃薯叶片生成的光合产物主要以蔗糖形式经韧皮部运输至块茎[27-28]。本试验中,低温条件下,非结薯植株叶片中StSEX4表达下调,而StSWEET1表达量显著上升,有助于叶片中的淀粉积累和同化产物向块茎转运。
CO/FT介导的光周期调控通路在马铃薯块茎形成过程中发挥着重要作用。拟南芥中,AtCO(Arabidopsis CONSTANS)的转录调控是光周期调控决定花期的重要因素[29],同样CO类转录因子StCO影响马铃薯块茎结薯时间,长日照(或者短日照加暗间断)下StCO沉默株系结薯提早[30]。StCO在块茎形成过程中同样响应温度变化,在马铃薯热敏感品种Kufri Chandramukhi和耐热品种Kufri Surya的块茎形成期进行昼/夜24℃/20℃和24℃/24℃处理,结果发现,处理14d后,2个品种在高温条件下叶片StCO表达量均显著高于适温处理[31]。本试验中,23℃/13℃处理下的非结薯植株叶片StCO表达量显著高于其他处理,这也印证了StCO在温度调控结薯时间上起抑制作用[26]。
马铃薯FT家族基因StSP6A在块茎形成过程中发挥重要作用。短日照条件下,叶片产生的StSP6A转运蛋白通过韧皮部运输到匍匐茎顶端诱导块茎形成[11]。Lehretz等[12]研究揭示了高温条件下,miRNA(SES)通过下调StSP6A表达抑制结薯。StSP6A转录水平受温度调控已在许多研究[32]中得到证明。马铃薯叶片中StFKF1表达量受土壤高温影响较大,高温环境可通过下调StFKF1的表达,从而降低StSP6A含量而使结薯受阻[14]。本试验结果表明,StFKF1和StSP6A表达量受夜间温度调控,夜间13℃下,StFKF1和StSP6A表达量均显著高于其他处理,推测这可能是因为对于非结薯植株来说,由于处于即将形成试管薯的准备过程,夜间低温刺激了叶片StSP6A的表达,从而导致23℃/23℃处理下的已结薯植株叶片StSFKF1和StSP6A的表达量相对较低,关于StSP6A在不同夜间温度下调控试管薯形成的机制还有待深入研究。
4 结论
夜间低温(13℃)会延缓马铃薯试管薯初始结薯时间,夜间高温(23℃)降低试管薯结薯率,而夜间适温(18℃)可促进试管薯形成。生物钟基因StTOC1和StELF4、糖代谢相关基因StSWEET1和StSEX4及光周期通路基因StFKF1、StCO和StSP6A均表现出显著的温度响应变化,表明夜间温度可通过影响多种代谢途径基因的表达调控块茎形成过程。
