极小种群野生植物贵州地宝兰群落生态位特征及种间关系
2022-06-21安明态李晓芳
张 央, 李 志, 安明态,①, 李晓芳, 汪 伟
(1. 贵州大学: a. 林学院, b. 贵州大学生物多样性与自然保护研究中心, 贵州 贵阳 550025; 2. 贵州省植物园, 贵州 贵阳 550004)
生态位理论通过对物种间以及物种和环境间的相互关系进行定量研究,能够反映种群对资源和生态空间的利用能力,同时生态位理论也是不同种群适应不同环境的重要生存策略和机制[1]。种间联结反映了群落的结构、功能和动态[2]。开展植物生态位特征和种间联结性研究,是了解群落结构变化和演替的基础[3]。濒危种在群落内种间竞争中通常处于较为不利的地位,竞争能力较差[4]。极小种群野生植物在种群竞争中也可能处在劣势地位。例如:极小种群野生植物玉龙蕨(SorolepidiumglacialeChrist)在种群间竞争中处于劣势,数量稀少,对资源的利用率低,不能形成优势种群[5];浙江安息香(StyraxzhejiangensisS. M. Hwang et L. L. Yu)与伴生种产生资源竞争导致其数量少[6]。但造成植物濒危也并不完全是生态位原因。例如:缙云秋海棠(BegoniajinyunensisC. I Peng, B. Ding et Q. Wang)的生态位宽度大,其生境适宜生存,种间竞争不激烈,生态位和种间的相互作用不是造成其濒危状态的主要原因[7,8]。因此,深入探究植物种内和种间的竞争程度及植物在群落中的作用和地位,对揭示群落中物种共存机制尤其是濒危物种的群落竞争状况及演替趋势等具有重大意义。
贵州地宝兰(GeodorumeulophioidesSchltr.)是兰科(Orchidaceae)地宝兰属(GeodorumJacks.)植物,产于中国贵州、广西和云南,模式标本产地为贵州罗甸,花较大且呈玫瑰红色[9],具有较高的观赏价值。贵州地宝兰自1921年发表以来便销声匿迹,直到2004年才在广西雅长被陆续发现[10],2021年6月,作者在罗甸进行兰科植物调查时首次在模式标本产地发现了开花的贵州地宝兰居群。贵州地宝兰在野外资源量极少,受威胁严重,被评为濒危(EN)等级[11],并被列入《全国极小种群野生植物拯救保护工程规划(2011—2015年)》名录。
目前,研究者在贵州地宝兰的资源现状和分类修订[12]、遗传多样性[13]、菌根真菌[14]以及无性繁殖[15]等方面开展了相关研究,认为其致危因子主要是种群缺乏竞争力、生境片段化、果实少、种子萌发率低以及人为干扰[10,12]。由于贵州地宝兰分布狭窄,野外居群少,加上调查不深入,对其群落生态学研究不足,贵州地宝兰是否因为种间竞争关系过大导致濒危尚不清楚,研究极小种群野生植物贵州地宝兰的生态位特征和种间关系是了解其濒危原因的重要途径。因此,本文对野外分布的贵州地宝兰17个居群所处群落进行调查,分析其群落的物种组成、生态位宽度、生态位重叠、种间联结和种间相关性,揭示生态位特征和种间关系在贵州地宝兰濒危原因中的生态作用机制,为该种的有效保护提供理论依据。
1 研究区概况和研究方法
1.1 研究区概况
贵州地宝兰主要分布在北盘江中上游(六枝—关岭段)、北盘江下游(册亨—望谟段)、南盘江和红水河流域,地理坐标范围为东经105.23°~106.67°、北纬24.81°~26.04°,分布区面积约9 511 km2。海拔范围为411~928 m,其中贵州罗甸县罗妥村为模式标本产地。研究区位于中国东南季风与西南季风交汇处[16],其地势由南向北突升,由南至北地貌依次为常态地貌、喀斯特地貌,土壤类型包括山地黄棕壤和石灰土[17]。区域内年平均气温18.4 ℃,年平均降水量1 100 mm;森林植被多为残存的次生林和次生灌丛[18]。
1.2 研究方法
2021年6月至8月,在贵州地宝兰居群所处的群落分别设置面积10 m×10 m的样地进行调查,共17个样地,样地基本信息见表1。由于研究目标种贵州地宝兰为草本,数量较少,且大部分居群位于空旷的林下,草本层植物稀疏,参考有关兰科植物的样地设置[19],每个样地设置1个面积10 m×10 m的乔木层样方,4个面积5 m×5 m的灌木层样方,在样地四角各设置1个面积2 m×2 m的草本层样方。对乔木层中胸径(DBH)≥5 cm的植物进行每木检尺,调查种名、胸径和株高;灌木层和草本层调查种名、株数、地径、株高和盖度。胸径采用围尺(精度0.1 cm)测量,地径采用游标卡尺(精度0.1 mm)测量,株高采用测高杆(精度0.1 m)或钢尺(精度0.1 cm)测量,盖度采用方格法[20]估测。

表1 贵州地宝兰群落样地的基本信息
1.3 数据分析
1.3.1 物种组成及优势种分析 依据群落调查结果,分别对乔木层、灌木层和草本层的种类进行统计并计算各种类的重要值[21],根据重要值确定各层的优势种。
1.3.2 生态位分析 生态位宽度采用Levins[22]和Shannon-Wiener[23]2种生态位宽度计算,生态位重叠采用Pianka生态位重叠值[24]计算。

1.3.4 种间相关性分析 采用Pearson相关性分析和Spearman秩相关性分析比较贵州地宝兰群落草本层种间的相关程度[27,28]。
1.4 数据处理
采用EXCEL 2016软件对相关数据进行统计和计算;采用R 4.0.3软件计算和分析生态位特征和种间联结性。
2 结果和分析
2.1 物种组成及重要值
调查结果(表2、表3和表4)显示:17个贵州地宝兰群落共有维管植物41科78属94种(含变种、亚种),其中,乔木层10科14属15种,主要为麻栎(QuercusacutissimaCarr.),重要值为63.90%,占据绝对优势;灌木层21科33属42种,主要以假木豆〔Dendrolobiumtriangulare(Retz.) Schindl.〕、麻栎和浆果楝〔Cipadessabaccifera(Roth.) Miq.〕占据优势,重要值分别为26.61%、23.57%和11.97%;草本层20科39属45种,主要以求米草〔Oplismenusundulatifolius(Arduino) Beauv.〕、仙茅(CurculigoorchioidesGaertn.)、藿香蓟(AgeratumconyzoidesLinn.)和贵州地宝兰占据优势,重要值分别为21.69%、18.21%、16.20%和15.10%,其中贵州地宝兰的重要值在草本层中位列第4,处于中上水平。

表2 贵州地宝兰群落乔木层物种组成及重要值

表3 贵州地宝兰群落灌木层物种组成及重要值

表4 贵州地宝兰群落草本层物种组成及重要值
2.2 生态位特征
2.2.1 生态位宽度 贵州地宝兰群落草本层45个种类中仅18个种类的Levins生态位宽度和Shannon-Wiener生态位宽度不同时为最小值(Levins生态位宽度为1.00且Shannon-Wiener生态位宽度为0.00),这18个主要种类的Levins生态位宽度和Shannon-Wiener生态位宽度见表5。由表5可以看出:贵州地宝兰群落草本层18个主要种类的Levins生态位宽度为1.00~14.85,Shannon-Wiener生态位宽度为0.00~2.76。18个主要种类的2种生态位宽度存在一定差异,但生态位宽度排名前8的种类一致,由高到低依次为贵州地宝兰、仙茅、求米草、藿香蓟、海金沙〔Lygodiumjaponicum(Thunb.) Sw.〕、淡竹叶(LophatherumgracileBrongn.)、金丝草〔Pogonatherumcrinitum(Thunb.) Kunth〕、芒(MiscanthussinensisAnderss.),表明这8种植物在草本群落中资源利用能力较强。该排序与重要值的排序基本一致,总体表现为重要值大的种类其生态位宽度也较大。草本层中贵州地宝兰的重要值位列第4,其Levins生态位宽度和Shannon-Wiener生态位宽度却位列第1,分别为14.85和2.76,表明贵州地宝兰在群落草本层的资源利用能力强于其他种类。
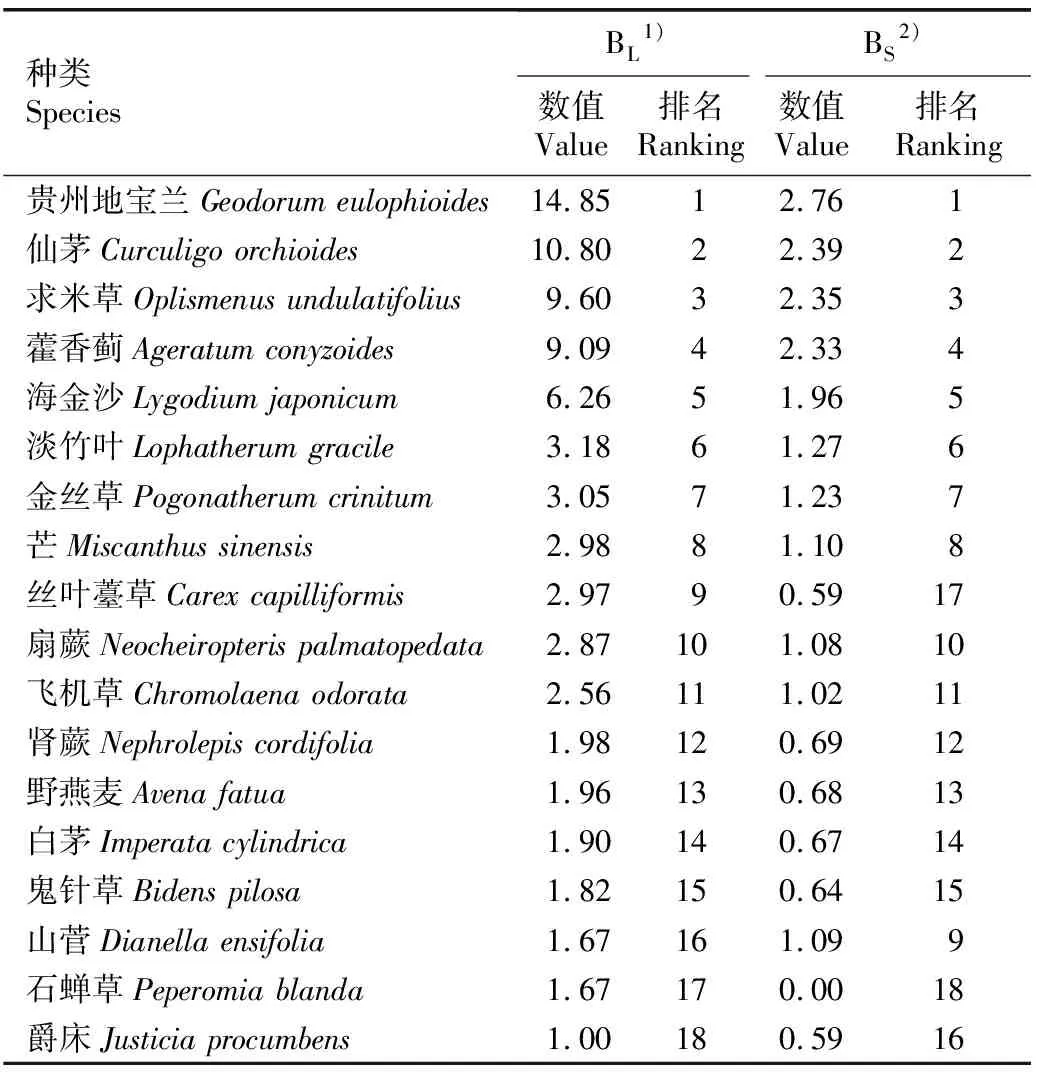
表5 贵州地宝兰群落草本层主要种类的生态位宽度
2.2.2 生态位重叠 贵州地宝兰群落草本层种类的Pianka生态位重叠值见表6。由表6可以看出:贵州地宝兰群落草本层45个种类中有23个种类形成的种对存在生态位重叠,共计100个种对,占总种对数(253)的39.5%,Pianka生态位重叠值在0.00~0.85之间,其中,Pianka生态位重叠值大于0.50的种对有19个,占总种对数的7.5%。贵州地宝兰与其他22个种类均存在生态位重叠,Pianka生态位重叠值为0.15~0.79,其中与仙茅、求米草、藿香蓟和海金沙的Pianka生态位重叠值较大,分别为0.79、0.66、0.61和0.57,表明贵州地宝兰与这4个种类在群落草本层中存在资源竞争关系。
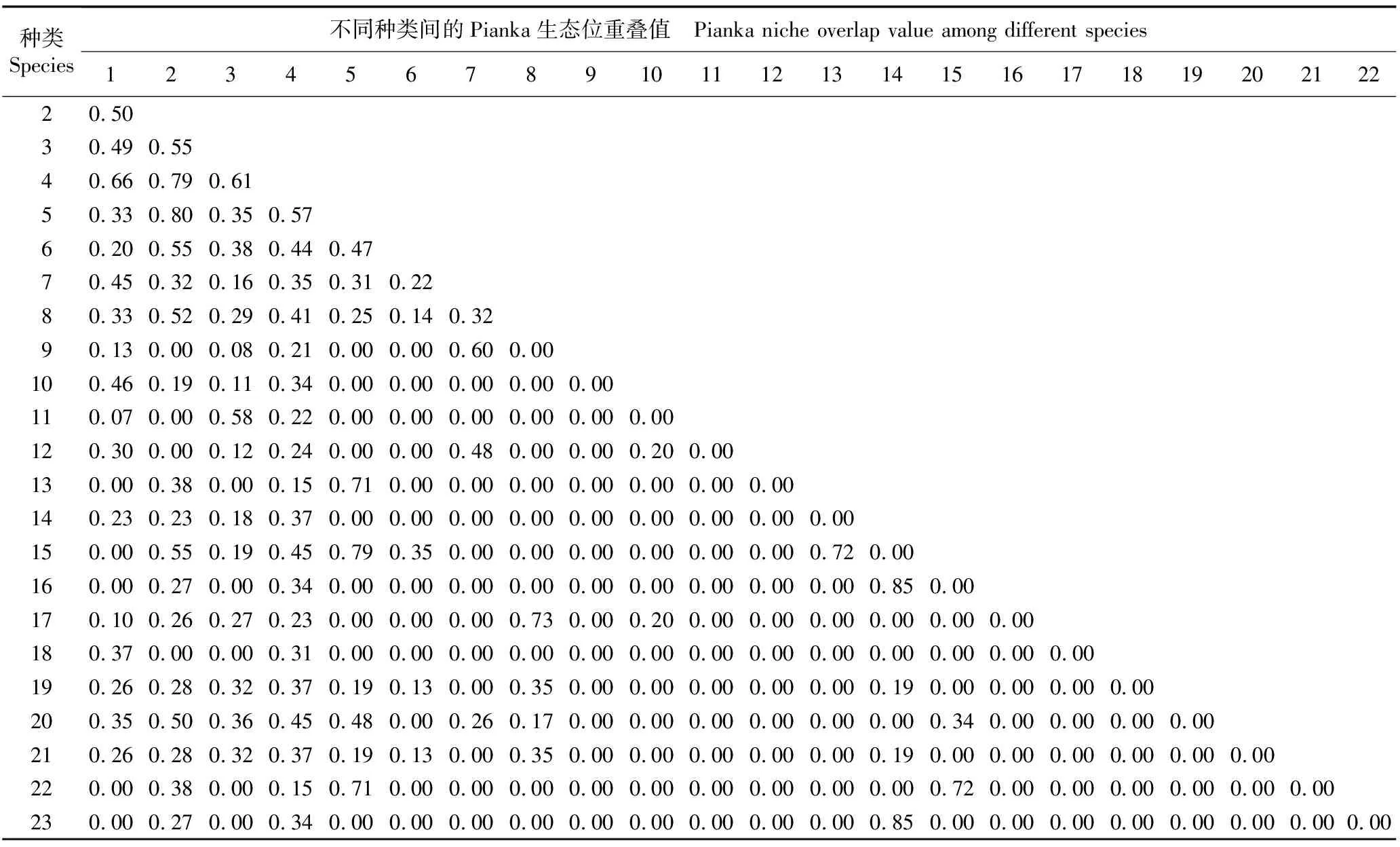
表6 贵州地宝兰群落草本层种类的Pianka生态位重叠值1)
2.3 关联性分析

2.3.2 种间联结分析 对贵州地宝兰群落草本层中形成生态位重叠的23个种类根据其出现与否建立2×2联列表,然后进行χ2检验,结果见图1。由图1可以看出:贵州地宝兰群落草本层上述23个种类构成的253个种对中,仅矛叶荩草〔Arthraxonlanceolatus(Roxb.) Hochst.〕与白茅〔Imperatacylindrica(Linn.) Beauv.〕1个种对呈极显著正联结,占总种对数的0.4%;63个种对呈不显著正联结,占总种对数的24.9%;2个种对呈显著负联结,占总种对数的0.8%;165个种对呈不显著负联结,占总种对数的65.2%;22个种对呈无联结,占总种对数的8.7%。不显著联结和无联结的种对共有250个,占总种对数的98.8%,表明贵州地宝兰群落草本层种类间的联结性较弱,各种类间关系较为松散,种类呈相对独立的分布格局。贵州地宝兰与石蝉草〔Peperomiablanda(Jacquin)Kunth〕呈无联结,与其他21个种类呈不显著联结,其中,与求米草、仙茅、鬼针草(BidenspilosaLinn.)、爵床(JusticiaprocumbensLinn.)和地胆草(ElephantopusscaberLinn.)5个种类呈不显著正联结,与藿香蓟、海金沙、金丝草、芒、野燕麦(AvenafatuaLinn.)、淡竹叶和狗尾草〔Setariaviridis(Linn.) Beauv.〕等16个种类呈不显著负联结。

◆: 极显著正联结Extremely significant positive connection; : 不显著正联结Non-significant positive connection; ▽: 显著负联结Significant negative connection; : 不显著负联结Non-significant negative connection; : 无联结No connection.
2.4 种间相关性分析
对贵州地宝兰群落草本层中形成生态位重叠的23个种类进一步进行Pearson相关性分析和Spearman秩相关性分析,结果见图2。
Pearson相关性分析结果(图2-A)表明:贵州地宝兰群落草本层上述23个种类构成的253个种对中,21个种对呈极显著(p<0.01)正相关,占总种对数的8.3%;10个种对呈显著(p<0.05)正相关,占总种对数的4.0%;25个种对呈不显著正相关,占总种对数的9.9%;5个种对呈极显著负相关,占总种对数的2.0%;11个种对呈显著负相关,占总种对数的4.3%;171个种对呈不显著负相关,占总种对数的67.6%;10个种对呈不相关,占总种对数的4.0%。不显著相关和不相关的种对共有206个,占总种对数的81.4%,表明贵州地宝兰群落草本层中大部分种类间的相关性不显著。贵州地宝兰仅与白茅存在极显著正相关,与求米草、丝叶薹草(CarexcapilliformisFranch.)和爵床存在显著正相关,与海金沙、荩草〔Arthraxonhispidus(Trin.) Makino〕、扇蕨〔Neocheiropterispalmatopedata(Baker) Christ〕和大白茅〔Imperatacylindricavar.major(Nees) C. E. Hubbard〕存在显著负相关,与其他14个种类不显著相关或不相关。
Spearman秩相关性分析结果(图2-B)表明:贵州地宝兰群落草本层上述23个种类构成的253个种对中,18个种对呈极显著正相关,占总种对数的7.1%;14个种对呈显著正相关,占总种对数的5.5%;36个种对呈不显著正相关,占总种对数的14.2%;19个种对呈极显著负相关,占总种对数的7.5%;24个种对呈显著负相关,占总种对数的9.5%;134个种对呈不显著负相关,占总种对数的53.0%;8个种对呈不相关,占总种对数的3.2%。不显著相关和不相关的种对共有178个,占总种对数的70.4%。贵州地宝兰仅与白茅存在极显著正相关,与求米草、丛毛羊胡子草(EriophorumcomosumNees) 、丝叶薹草、爵床和地胆草存在显著正相关,与其他16个种类不显著相关或不相关。
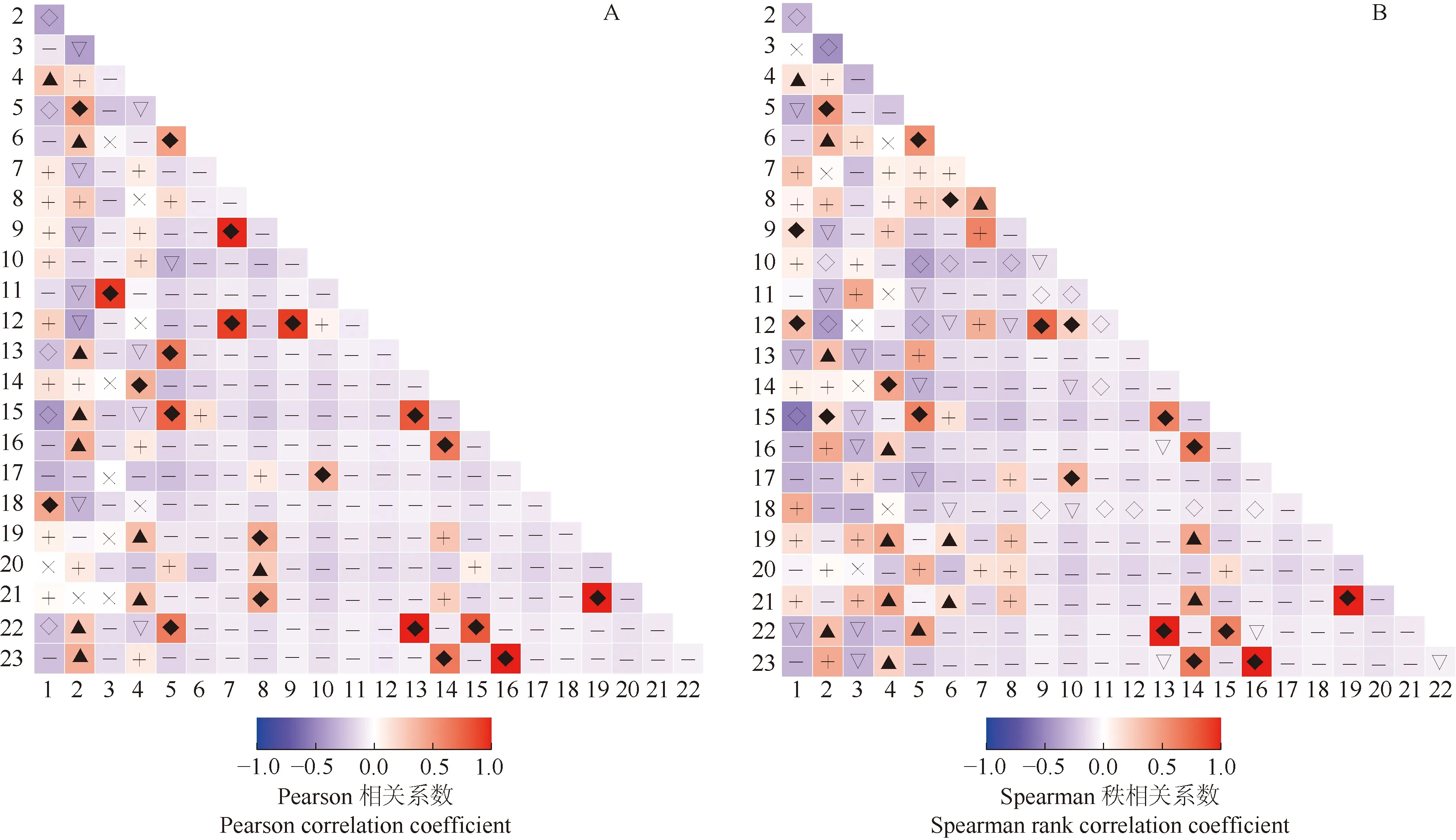
◆: 极显著正相关Extremely significant positive correlation; ▲: 显著正相关Significant positive correlation; +: 不显著正相关Non-significant positive correlation; ◇: 极显著负相关 Extremely significant negative correlation; ▽: 显著负相关 Significant negative correlation; -: 不显著负相关Non-significant negative correlation; ×: 不相关No correlation.
3 讨论和结论
植物群落物种组成受植被生物学、干扰因子、地形和生境异质性等因子的综合影响[29]。本次调查发现,贵州地宝兰在北盘江下游(册亨—望谟段)、南盘江和红水河流域的分布点为常态地貌,土壤类型以山地黄棕壤为主,土层深厚,肥力较高,植被以麻栎林为主;在北盘江中上游(六枝—关岭段)的分布点主要为喀斯特地貌,土壤类型以石灰土为主,土壤瘠薄,水土流失较为严重,植被以次生林为主。由于各贵州地宝兰群落样地的地貌和林分的不同,不同群落的物种组成差异较大,具体表现为喀斯特地貌物种组成丰富,主要以浆果楝和山麻杆(AlchorneadavidiiFranch.)为主,常态地貌物种组成单一,主要以麻栎和假木豆为主。综合17个样地来看,贵州地宝兰在草本层的重要值仅位列第4,并不占据绝对优势,在与求米草、仙茅和藿香蓟的竞争中可能处于劣势。
生态位宽度受种类在样方中出现频度的影响,且与重要值并不存在绝对的相关关系[30],种群的生态位宽度越大,对环境的适应能力强和资源利用能力也越强[31]。本研究中,贵州地宝兰在草本层中的重要值和生态位宽度计算结果显示:其重要值位于第4,但Levins生态位宽度和Shannon-Wiener生态位宽度均为最大,分别为14.85和2.76,表明贵州地宝兰的资源利用能力强于该群落草本层其他种类。本研究结果也表明:植物种类的重要值与其生态位宽度并无直接关系。重要值大的种类对群落的构建及生态位功能起决定性作用[30],例如:贵州晴隆县立新村居群所在群落草本层中金丝草的重要值最大,为优势种,对该居群所在群落的草本层构建起着决定性作用,但由于金丝草在其他群落中分布频度低,导致其整体重要值及生态位宽度不高。
生态位重叠反映了同一生态系统中不同种类对资源需求的相似程度[32],生态位重叠值越大,种类之间对资源的利用程度越相似,竞争可能越激烈[33]。本研究中,贵州地宝兰17个群落草本层的45个种类中仅有23个种类形成的种对存在生态位重叠,共计100对,占总种对数的39.5%,表明贵州地宝兰群落草本层种类间整体上对资源利用竞争较小。草本层形成生态位重叠的23个种类中,贵州地宝兰与其他22个种类均存在生态位重叠,但Pianka生态位重叠值大于0.5的仅有仙茅、求米草、藿香蓟和海金沙4种,说明贵州地宝兰与这4种植物具有相似的资源利用方式,在未来群落物种共存及演替过程中可能会存在竞争关系。本研究还发现,物种的生态位宽度越大,与其他物种的生态位重叠可能性越大,贵州地宝兰、仙茅、求米草和藿香蓟这4个生态位宽度较大的种类与群落中其他种类均形成了较为广泛的生态位重叠,达到了68对,占生态位重叠种对数的68.0%。该结果与红豆杉〔Taxuschinensis(Pilger) Rehd.〕[34]的群落生态位研究结果相似,二者均表明生态位宽度较大的种类,其生态位重叠也较大。
种间联结性则反映了群落中种间的相互作用及物种与环境因子间的耦合关系[35]。本研究中,贵州地宝兰群落草本层种类间总体联结为不显著负联结,意味着草本层还处于不稳定的演替阶段,种间相对独立,在受到外界干扰时种类和数量易出现波动。χ2检验结果表明:贵州地宝兰群落草本层种间呈不显著联结和无联结的种对共有250个,占98.8%,种间联结性较弱,各种间关系较为松散,物种呈相对独立的分布格局。Pearson相关性分析和Spearman秩相关性分析结果均表明:贵州地宝兰与白茅存在极显著相关,与爵床、丝叶薹草和求米草存在显著正相关,说明在相同环境条件下贵州地宝兰与这4种植物在一定程度上是“互利互惠”的关系。
在进行种间关系研究时,通常以χ2检验为基础,定性判断种间联结性的显著性,并以Pearson相关性分析和Spearman秩相关性分析等方法进行检验,才能全面、准确地反映植物群落的种间联结性[36]。本研究中,Spearman秩相关性分析呈极显著和显著相关的种对共有75个,大于Pearson相关性分析呈极显著和显著相关的种对数(47),即Spearman秩相关性分析的灵敏度比Pearson相关性分析高。这与刘润红等[37]对岩溶石山青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕群落研究结果一致,这是因为Spearman秩相关性分析是一种非参数检验,更符合自然界物种呈集群分布的特点,而Pearson相关性分析是一种参数检验,更多地要求物种服从连续的正态分布[36]。
综上所述,对贵州地宝兰的保护应注重群落种间关系的维持,建议在居群分布点建立保护小区(点)进行就地保护,同时进行动态监测,加强对放牧活动的管控力度。本研究仅从群落生态学的角度揭示了贵州地宝兰群落现阶段的生态位特征和种间关系,但对于群落间物种共存及种间关系的长期维持机制尚不明确,这也是其他珍稀濒危植物值得长期关注和研究的内容。
