干热河谷次生稀树灌木林优势种群空间格局及种间关联性研究
2022-06-16龙成余志祥杨永琼税梅梅
龙成 ,余志祥 ,杨永琼 ,税梅梅
1.攀枝花学院,四川 攀枝花 617000;2.四川攀枝花苏铁国家自然保护区,四川 攀枝花 617000;3.攀枝花市建筑工程学校,四川 攀枝花 617000
种群是构成群落的基本单位,也是理解植物群落结构组成与动态变化机制的关键[1]。种群年龄结构不仅反映不同种群大小和年龄个体的组配情况,也反映了种群数量动态及其发展趋势[2-3]。近年来,国内出现了以种群结构为研究内容的报导。但多数学者,像刘贵峰[4]、涂云博[5]、袁志忠[6]等只是单纯地针对种群结构进行研究,并没有涉及种群在群落中的分布情况。然而,研究植物分布格局对揭示种群的形成和维持机制有着重要的理论意义,与种群结构的研究相辅相成,一直以来都是植物生态学领域的研究热点[7]。国内的学者也相应地做了一系列的工作。蒙荣[8]等运用地统计学的方法,对短花针茅荒漠草原群落优势种群进行了空间格局分析;张金屯[9]和李素清[10]等,则分别应用DCA 排序与格局分析方法相结合、DCA 排序与双项轨迹方差法相结合的方法,完成了对相关种群分布格局的研究;两人又在2008 年共同阐述了随机配对法在亚高山草甸优势种群格局分析中的应用[11]。然而,这些研究都是基于一种或几种固定尺度下进行的,没有把空间格局与尺度变化有机的结合起来。为了克服这些问题,近年来,对于物种空间格局的研究,多采用点格局分析法的K函数[12-15],并且出现了一系列的研究进展。张金屯[16]在阐述植物空间点格局分析时应用K函数对美国密歇根州克林顿县栎林的3 个优势种群进行了研究;杨晓晖[17]等对白刺沙堆进行空间格局分析;宋于洋[18]等则对梭梭种群不同发育阶段的空间格局及其关联性进行了分析。通过这些学者的研究可以说明多尺度点格局分析是格局分析方法的重大突破。但K函数因其存在显著的尺度累积效应(即小尺度累积效应影响大尺度格局分析结果)。因此,通过圆环代替K函数中圆以有效消除尺度累积效应的O-ring 函数便应运而生,并很快应运到各类种群点格局分析中[19]。
干热河谷属生态脆弱区,土壤侵蚀严重[20-22],以“南亚热带为基带的立体气候”高温、干旱、蒸发量大,这里植物物种较为稀少,但中国特有物种和濒危物种在此均有分布[23],发育着以草地和多汁、带刺灌木为主的河谷特殊类型半稀树草原、稀树草原植物群落[24]。国内干热河谷森林群落主要分布在云南和四川两省[24],目前对干热河谷森林群落的研究主要集中在云南省干热河谷群落结构[25]、植物生物量[26]、生物多样性[27-28]、植物对土壤养分和水分的利用[29]等方面。四川攀枝花苏铁国家级自然保护区内1 500 m~2 300 m,主要为干热河谷次生稀树灌木林,是干热河谷特殊类型稀树、半稀树草原植物群落—干热河谷稀树灌木林经人为干扰(放牧)退化后,常年受季节性焚风效应影响,在山体背风坡形成以低矮灌木和草本植物为建群种的次生稀树灌木林群落,并处于向气候顶极(干热河谷常绿针阔叶混交林[24])演替的次生演替初级阶段。目前,基于永久固定样地的干热河谷植物群落动态及演替的研究还较鲜见,对四川攀枝花苏铁国家级自然保护区内干热河谷次生稀树灌木林的研究更是尚未起步。研究基于四川攀枝花苏铁国家级自然保护区干热河谷次生稀树灌木林内1 hm2固定样地,通过分析优势种群组成、空间分布格局及种间关联性,以期较为全面地掌握该群落内优势种群现状、分布状况及发展动态,旨在为干热河谷稀树灌木林乃至整个干热河谷生态系统的演替和生物多样性维持机制的研究奠定物种动态及空间格局方面的基础。
1 研究区概况
研究地位于川、滇两省交界横断山脉东南前缘向云贵高原过渡地带的四川攀枝花苏铁国家级自然保护区(101°32′15″~101°35′46″E,26°36′31″~26°38′24″N),是金沙江河谷较为典型的地段,地势陡峭,河谷深切,山体相对高度大,地形封闭。区内保护对象为干热河谷常绿针阔叶混交林生态系统及其内部的野生动植物、地貌景观(河谷景观)等。保护区总面积13.58 km2,受干热河谷气候效应影响,研究区属于南亚热带半干旱河谷气候类型,冬季气候温和,日照充足,热量丰富,年均气温19.7℃~20.5℃,是四川省年均气温最高的地区。5 月最热,12 月或1 月最冷。6 月上旬至10 月为雨季,11 月至翌年5 月为旱季,年无霜期超过300 d。保护区地势由西北向东南倾斜,北面丰家梁子横亘、纵列“鸡爪梁子”众多,最高海拔2 259.6 m (团山包),最低海拔1 120 m (猴子沟与保护区边界交点),相对高差1 139.6 m。区内大部分岩石主要为震旦系灯影组石灰岩和奥陶系白云质石灰岩,土壤发育具有较为明显的垂直分布特征,1 500 m 以下海拔高度分布红色石灰土,1 500 m 以上为棕黄色石灰土。保护区内地带性顶极植被为干热河谷常绿针阔叶混交林,研究样地主要优势物种为攀枝花苏铁(Cycas panzhihuaensis)、铁橡栎(Quercus cocciferoides)、滇榄仁(Terminalia franchetii)和蒙桑(Morus mongolica)等[23]。
2 研究方法
2.1 样地概况及调查方法
2015 年7—10 月,参照美国史密森热带森林科学研究中心(CTFS)监测样地的建设方法,用全站型电子速测仪(GTS-102N,Topcon Corporation,Tokyo,Japan),在四川攀枝花苏铁国家级自然保护区干热河谷次生稀树灌木林内建立了面积1 hm2(100 m ×100 m)的固定样地,并将样地划分为400 个5 m × 5 m的小样方,标定并调查样方内所有胸径(DBH) ≥1 cm的木本植物,调查内容包括:记录每株植物的学名、胸径大小、树高以及坐标等,不能现场鉴定的植物种,制成标本内业鉴定。
2.2 群落优势种群组成
采用《植物群落学》[30]和《植物群落学实验手册》[31]上的方法计算木本植物各物种的重要值,并以重要值表征物种在群落中的优势程度[33]。优势物种取重要值大于1.5 的树种。其中,重要值(IV)=[相对多度(RD)+相对频度(RF)+相对显著度(RP)]/3。相对多度(RD)=单个种群多度(D)/所有种群多度之和 × 100;相对频度(RF)=种群频度(F)/所有种群频度之和 × 100,种群频度(F)=种群个体出现样方数/总样方数;相对显著度(RP)=种群显著度(P)/所有种群显著度之和 × 100,种群显著度(P)=种群所有个体胸径高断面积之和。
2.3 群落优势种群空间格局及种间关联
采用单变量(Univariate)和双变量(Bivariate)Oring 统计函数分别计算干热河谷次生稀树灌木林群落优势种群空间格局和种间关联性[32]。UnivariateOring 统计函数计算公式如下:

式(1)中,n1为格局1 点的数目,为以第i个点为圆心,半径为r,宽为w的圆环,Point1[X]为格局1 中,X区域内点的数目,Area[X]为区域X的面积。
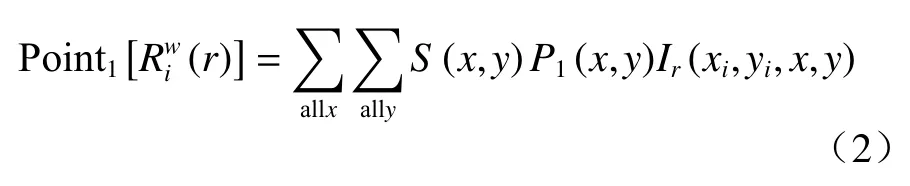
式(2)中,(xi,yi)为格局1 中第i个点的二维坐标,S(x,y)为公式中一变量,若坐标(x,y)在研究区域内,则S(x,y)=0;P1(x,y)表示落在每个单元格内格局1 点的数目。Ir是一个以格局1 中第i个点为中心,半径为r,宽为w的圆环而变化的变量,其表达式如下:
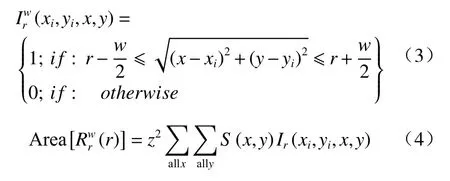
式(4)中,z2表示一个单元格的面积大小。
BivariateO-ring 统计函数公式如下:
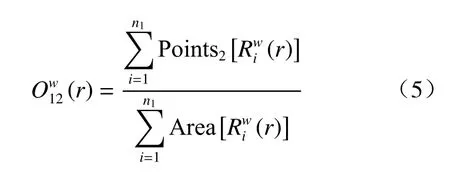
式(5)中,Points2[X]为格局2 中,X区域内点的数目。其他各项意义与(1)式均相同。
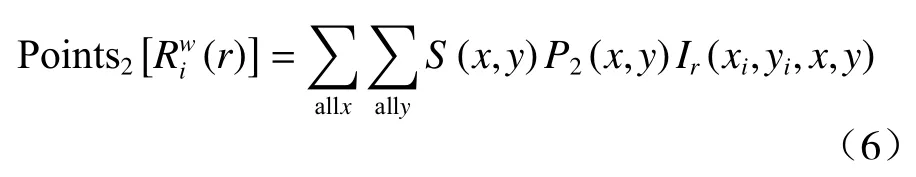
式(6)中,P2(x,y)表示落在每个单元格内格局2 点的数目。其他各项均与式(2)相同。此外,式(6)中Ir(xi,yi,x,y)的表达式与式(3)相同。
在Univariate 和BivariateO-ring 统计分析中,如果O11(r)或O12(r)值大于置信区间上限(即上包迹线),则该种群在该距离尺度显著聚集或两种群在该距离尺度呈显著正相关;如果O11(r)或O12(r)值在置信区间上下线之间(上下包迹线间),则该种群在该距离尺度随机分布或两种群在该距离尺度无显著相关性;如果O11(r)或O12(r)值或O12(r)值小于置信区间下限(即下包迹线),则该种群在该距离尺度均匀分布或两种群在该距离尺度呈显著负相关。本研究单变量和双变量O-ring 统计均采用Programita 2004 模拟19 次以得到95%的置信区间。
2.4 聚集规模与优势度回归分析
当种群呈聚集分布时,通常把偏离随机置信区间上限的最大值作为最大聚集强度指标,而聚集规模则为以最大聚集强度为半径的圆[39]。以IBM SPSS Statistic 19.0 软件完成聚集规模与优势度的回归分析。本研究所有绘图均采用Origin Pro 8.0 完成。
3 结果与分析
3.1 群落优势种群组成
干热河谷次生稀树灌木林群落1 hm2样地内重要值>1.5 的优势物种为攀枝花苏铁(C.panzhihuaensis)、铁橡栎(Q.cocciferoides)、滇榄仁(T.franchetii)、蒙桑(M.mongolica)、岩柿(Diospyros dumetorum)和山槐(Albizia kalkora)。除攀枝花苏铁为灌木生长型其余优势物种皆为乔木或小乔木型(见图1)。
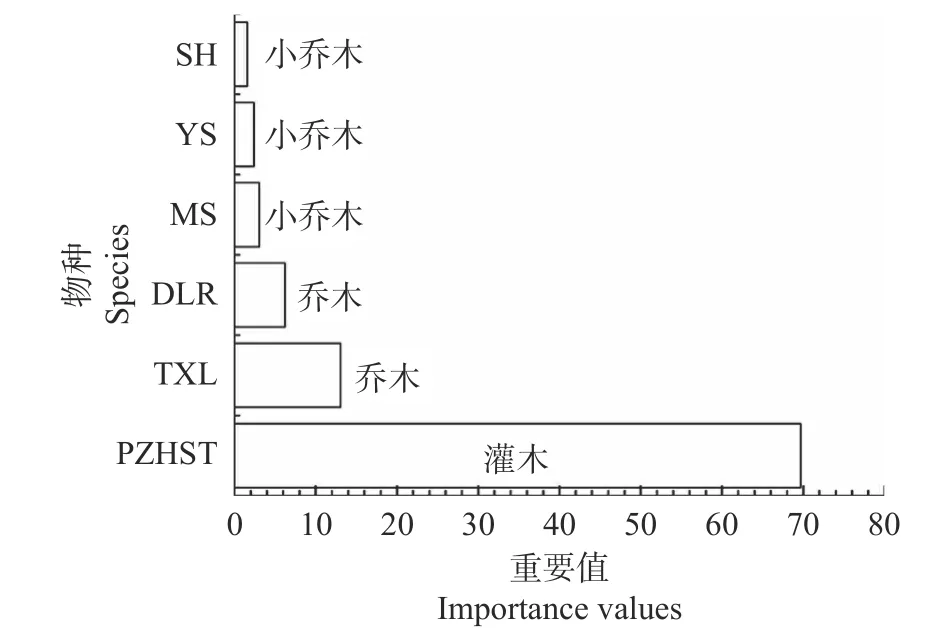
图1 干热河谷次生稀树灌木林群落优势种群重要值Fig.1 Importance value of dominant populations in secondary savanna shrub forest in dry-hot valley
3.2 群落优势种群空间分布格局
由图2 可知,各优势种群的空间分布格局均以随机分布为主。攀枝花苏铁种群(A)仅在1 m 处呈显著聚集分布,其余尺度范围均呈随机分布。其他种群如铁橡栎种群(B)在(1 m、24 m、37 m、44 m),滇榄仁种群(C) (3 m、5 m、9 m、11 m、15~23 m、25 m、27 m、40 m、43 m),蒙桑种群(D)在(13 m、15 m、20 m、26 m、32 m、35 m、44 m),岩柿种群(E)在(8 m、13~14 m、21 m、30 m、37 m)和山槐种群(F)在(4~5 m、24 m、31~32 m) 也有显著聚集现象出现。虽然攀枝花苏铁和山槐种群无均匀分布现象,但其他优势种群在不同尺度上均有均匀分布现象出现,例如,铁橡栎(2 m、11 m、30 m、40 m、50 m),滇榄仁(8 m、37 m、44 m、46 m),蒙桑(17 m、19 m、24 m、31 m、36~37 m、40 m、50 m),岩柿(39 m、42 m、44 m、46~47 m)。

图2 干热河谷次生稀树灌木林群落优势种群空间分布格局(A:攀枝花苏铁,B:铁橡栎种群,C:滇榄仁种群,D:蒙桑种群,E:岩柿种群,F:山槐种群。)Fig.2 Spatial distribution pattern of dominant populations in secondary savanna shrub forest in dry-hot valley (A:C.panzhihuaensis,B:Q.cocciferoides,C:T.franchetii,D:M.mongolica,E:D.dumetorum,F:A.kalkora)
3.3 群落优势种群种间关联性
由图3 可知,与攀枝花苏铁种群负相关尺度范围较大的种群分别为铁橡栎种群(1~17 m;19 m;21 m)(见图3A-B)与滇榄仁种群(1~17 m;21 m) (见图3AC),岩柿种群(1 m;8 m;13 m)则最小(见图3A-E),蒙桑种群次之(4 m;11 m;13~14 m;16 m;24 m) (见图3A-D),相较而言,山槐种群与攀枝花苏铁种群在较小尺度范围负相关分布则较为分散(1~3 m;5~6 m;11~13 m) (见图3A-F)。铁橡栎种群与攀枝花苏铁种群在大于27 m 的尺度范围上出现正相关,且在33~50 m 范围内出现连续正相关(见图3A-B),山槐种群与攀枝花苏铁种群出现正相关则在大于31 m 的尺度上,且存在集中尺度范围的正相关现象(36~38 m;40 m;42~45 m;47 m;49~50 m) (见图3A-F)。其余种群在较大尺度上则对攀枝花苏铁种群存在零星尺度上的正相关作用(滇榄仁:31 m;40 m;42 m;47 m;49 m;蒙桑:34 m;44~45 m;47 m;岩柿:31 m;34 m;42 m)。铁橡栎种群与岩柿种群仅出现负相关(见图3BE),与其余优势种群呈现正、负相关随尺度增大而交替出现的现象(见图3A-B;B-C;B-D;B-F)。铁橡栎种群与山槐种群正相关作用在较小尺度上率先出现(6 m;10 m),随着尺度的增大,在15 m 处出现负相关,随后又在38 m 和48 m 尺度上出现正相关(见图3B-F)。铁橡栎种群与蒙桑种群负相关作用尺度范围较大(7~8 m;11 m;14 m;16~20 m;24~27 m;29 m;32 m;35 m;41 m;44 m),正相关作用仅出现在34 m、48~50 m 尺度上(见图3B-D)。铁橡栎种群与滇榄仁种群、蒙桑种群与岩柿种群以及岩柿种群与山槐种群均表现出正、负相关交替出现(图3B-C;D-E;E-F)。滇榄仁种群与蒙桑种群仅在7~8 m 尺度上表现出正相关,其余尺度上背离置信区间的均为负相关(见图3C-D),与岩柿种群则表现出较小尺度先呈正相关,随着尺度增大逐渐表现为负相关(见图3CE)。山槐种群与滇榄仁种群和蒙桑种群负相关首先出现在较小尺度上,尺度增大后逐渐呈正相关(见图3C-F;D-F)。
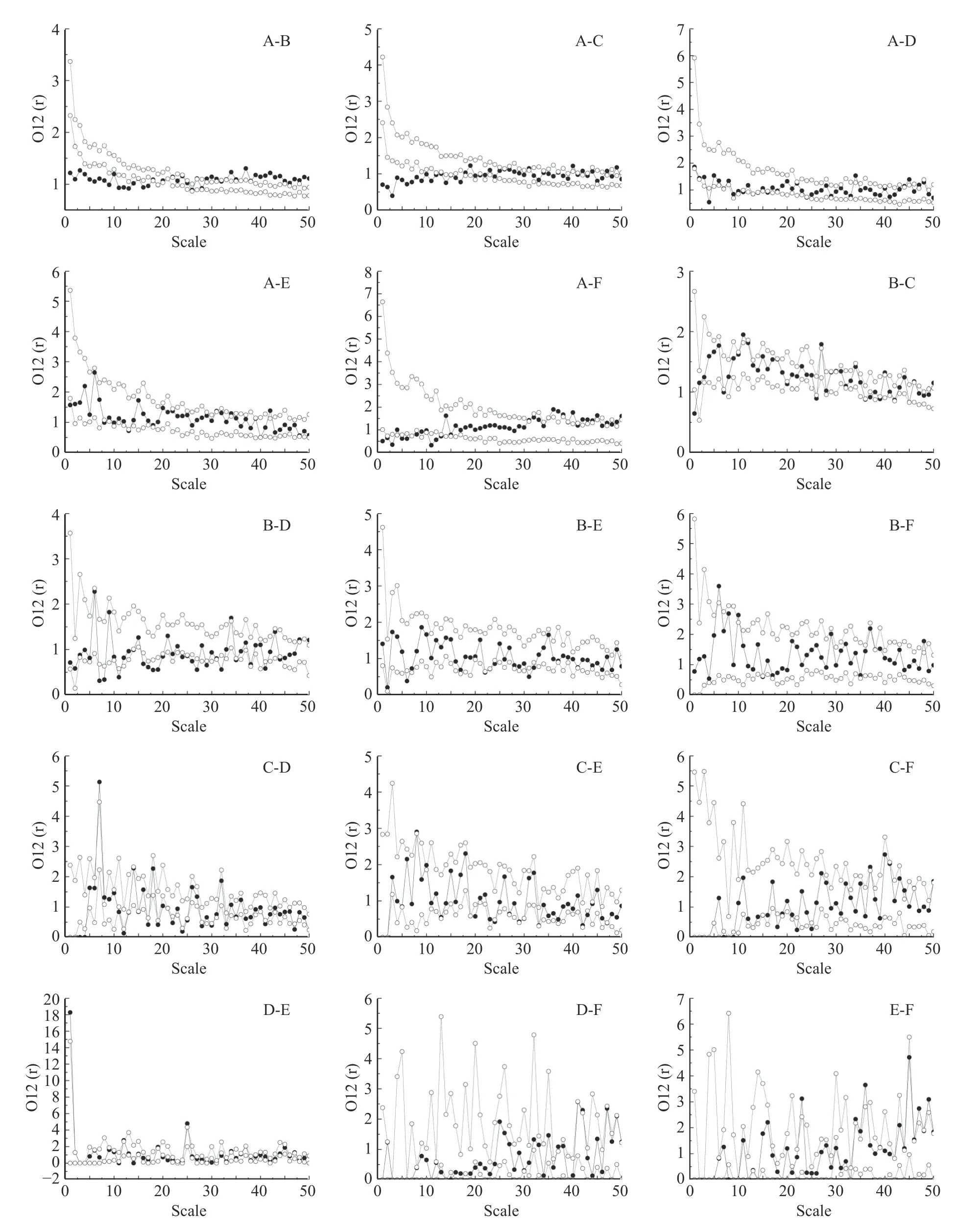
图3 干热河谷次生稀树灌木林优势种群种间关联性Fig.3 Interspecific association of dominant populations in secondary savanna shrub forest in dry-hot valley
3.4 群落各优势种群聚集规模与优势度的回归分析
干热河谷次生稀树灌木林群落6 个优势种群的聚集规模与优势度(重要值)显著线性相关(见图4),计算得出R=0.99,P=0.00 < 0.01,表明二者在0.01水平上显著相关。回归方程为:y=(127.49 ± 9.04)x+(4.78 ± 1.88),Adj-R2=0.98,P=1.46993E-4 < 0.01,回归方程判定系数良好,具有代表性。
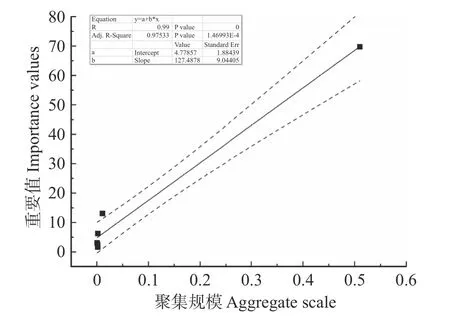
图4 聚集规模与重要值的回归分析Fig.4 Regression analysis of aggregate scale and importance value
4 讨论与结论
干热河谷次生稀树灌木林群落以攀枝花苏铁优势度最高(见图1)。除攀枝花苏铁外,其余优势种群均是乔木或小乔木,说明该群落尚处于演替的初级阶段[38]。随着森林次生演替进程,群落内部环境(例如,光照)将逐渐适于乔木树种生长,乔木型种群或会逐渐占据优势地位[42]。
从空间格局研究结果来看,聚集尺度最小的种群为攀枝花苏铁种群,仅在1 m 处聚集,说明当前群落内环境对该种群影响较小,幼龄个体仅产生小规模聚集化以发挥种群群体效应,应对不良环境和外来种的侵入[40,43]。因此,母株周围幼龄植株个体具有较高的存活率,种群更新较为良好。虽然攀枝花苏铁种群在其余尺度上皆呈随机分布(见图2-A),但在铁橡栎和滇榄仁等优势种群的负相关作用影响下(见图3A-B~A-F),在某些尺度上已出现以群体效应低于不良环境的聚集分布(见图2-A)[43]和种间竞争作用过后的均匀分布(见图2-A)[44]。此外,虽然攀枝花苏铁种群聚集尺度较小,但较强的聚集强度使其具有优势种群中最大的聚集规模,而该群落中种群优势度与聚集规模成正比(见图4),故干热河谷次生稀树灌木林群落内种群的聚集效应成为种群优势与否的关键因素[44]。群落中其余5 个优势种群空间分布格局中随机分布尺度比例均较攀枝花苏铁种群低,但均大于50%,说明干热河谷次生稀树灌木林群落6 个优势种群空间格局均以随机分布为主,这可能与森林次生演替初级阶段群落内环境异质化程度不高,生态位分化程度较低或尚未分化相关[46]。相较而言,攀枝花苏铁对当前群落内环境的适应程度其余5 个优势种群较高,加之其种群较大的聚集规模[43],均为其最终成为群落现阶段优势种群奠定基础。在聚集分布方面,滇榄仁种群聚集分布尺度占比最高,这可能与其生长发育对水分需求较高有关[45],聚集可使其产生适宜种群生长的微环境(例如,荫蔽条件下水分条件较好等)以避免不良环境(例如,干热河谷蒸发量大,土壤含水量较低等[20-22])的影响,使种群得以正常发育。而该种群较小的均匀分布比例说明其也存在来自其他优势种群(例如,攀枝花苏铁和蒙桑等)的竞争压力[44]。在均匀分布比例方面,各优势种群均较低(小于20%),其中蒙桑种群相对较高(见图2-D),该种群经历过两次内部密度调节,铁橡栎和滇榄仁种群与蒙桑种群的种间竞争是其两次种内密度调节的主要原因。通过密度调节适应群落内资源和环境,又因其仅次于滇榄仁的聚集分布比例,表明该种群在一些尺度上也存在以聚集抵御不良环境[43]以及其他优势种群(例如,铁橡栎和滇榄仁等)负相关作用(见图3B-D;C-D)的现象[44]。
此外,干热河谷次生稀树灌木林群落内植株聚集规模是改变当前群落种群优势地位的主要驱动力(见图4)。攀枝花苏铁种群也因其较大的种群聚集规模(见图4)、较多的幼龄个体和大龄个体而成为该群落内最为优势的种群(见图1),也是群落中的建群种。聚集规模大的种群,能够更好地抵御干热河谷不良气候条件影响[20-22,43],最终发育成为群落现阶段的优势种群。同时也说明在常年焚风效应干扰下,干热河谷次生稀树灌木林群落内部各种群均已形成自身特有的生存规律,这与其他类型森林群落中物种的适应性报导相似[34,41,43,49]。
干热河谷次生稀树灌木林群落种间关联性以无相关性为主,负相关次之,正相关所占比例最低(见图3)。除攀枝花苏铁与铁橡栎种群无关联性尺度比例较低外(18%),其余种群间无关联性尺度所占比例均大于50%,这可能与群落尚处于次生演替初级阶段,落内生态位分化亦未完成[47],相对丰富的资源和较小的群落及种群密度使群落密度尚未达到最大环境承载力[48],种群内及种群间因空间、资源而发生的竞争规模相对较小相关。在负相关方面,攀枝花苏铁、蒙桑分别与铁橡栎和滇榄仁种群负相关尺度占比较高(大于30%),这一方面可以反映蒙桑、铁橡栎以及滇榄仁种群均匀分布格局的成因[44],另一方面也使滇榄仁和蒙桑(聚集尺度占比较大)、攀枝花苏铁和铁橡栎(聚集规模较大)等种群的聚集分布得到了很好地解释[43]。另外,群落内植物种间还存在占比较低的正相关作用,这与其他类型森林群落植物间存在相互促进生长的报导相似[49-50],说明群落内各种群也可能进化出以相互促进生长的方式来抵御干热河谷不良气候条件的行为。其中,较为显著的正相关作用物种对为攀枝花苏铁种群与铁橡栎种群、山槐种群以及岩柿种群与山槐种群等(见图3),表明铁橡栎、山槐、岩柿种群直接或间接促进攀枝花苏铁种群的生长发育,这在攀枝花苏铁种群就地保护管理方面可加以利用。
此外,由于干热河谷次生稀树灌木林尚处于次生演替初级阶段,群落内物种周转较快,当前群落内优势物种的地位是否会随着演替进程而发生变化?空间格局研究结果中,各优势种群多数尺度均呈随机分布,是否能说明干热河谷次生灌木林次生演替初期阶段群落具有“中性”或“偏中性”特征?这种以随机分布为主的格局分布特征是否会随森林次生演替的进行而发生变化?干热河谷“焚风效应”又在其中起到怎样的作用?对于这些问题的解答,尚须日后深入研究。
致谢四川攀枝花苏铁国家级自然保护区管理局提供试验场地,该单位工程师李贵能,攀枝花学院王定宏、谢宇宣、尼苦曲哈、晚凤等同学参加野外调查,特此感谢!
