贡嘎山黄崩溜沟泥石流迹地的植被恢复
2022-06-16王晓琴王莉秦世豪
王晓琴 ,王莉* ,秦世豪
1.四川省林业科学研究院,四川 成都 610081;2.成都理工大学旅游与城乡规划学院,四川 成都 610059
由于历史上的不合理采伐和森林的退化,我国的原始林已经遭到不同程度的破坏,造成我国已经出现可采资源匮乏的局面,同时引发一系列环境问题[1]。全球变化和一些自然灾害也对森林产生影响,所以针对不同阶段退化天然林的恢复与重建变得尤为迫切[2]。世界各国都在关注生态系统的恢复与重建,我国在开展的相关工作已经走在全球前列[3,4]。东北和西南是我国的两大林区,在西南的亚高山通常是云杉和冷杉为建群种的森林分布,现存的一些原始林也遭到一定的扰动和干扰,形成大面积的次生林[5],刘兴良等对中国天然次生林进行了诊断、类型分类与评价方法的探讨,为我国次生林恢复提供了科学依据[6]。天然林的恢复也是植被演替过程,特别是原生裸地开始的演替,将会展现出原生演替序列。Clements 的植被演替理认为演替是一个有序,并可以预测的过程,演替是生物群落与环境相互作用导致环境变化的结果[7]。植被恢复是合理的结构、高效的功能和和谐的关系[8]。只要对退化生态系统停止扰动,植被就会向原群落方向进展演替,并且恢复过程与原群落结构相比是由低到高的[9]。鄢武先对卧龙地震灾后植被恢复和大熊猫栖息地选种研究,对植被恢复的植物选择提供了依据[10]。因此,研究植被恢复应首先搞清楚植被的自然恢复全过程,对长时时间序列自然恢复过程的研究,更有助于科学规划和布局,提出针对性自然可行的恢复技术手段,发挥森林生态系统良好的生态效益和经济效益。
1 研究区概况
研究区位于贡嘎山海螺沟海拔 3 000~3 100 m的地段,黄崩溜沟是海螺沟的一条支流,年平均气温4.2℃,年最高气温为23.2℃,年最低气温为-14.0℃。由于受季风和地形的影响,该地区降水比较充沛,降水量大,年降水日数多达261d,蒸发量小。气候特征总体表现为寒凉气候,降水丰沛,雾日、雨日长,日照时数少,日照百分率低,湿度大。从上世纪初至今共爆发了7 次大规模泥石流,堆积的泥石流扇都是在毁坏森林和土壤后形成的,泥石流扇周边的峨眉冷杉林为迹地提供了丰富的种源。在不同时期形成的泥石流迹地上发育着不同的植物群落,形成间断的植被原生演替序列。不同植被恢复时期的群落与海螺沟冰川退缩区相应阶段群落有着高度的相似性,通过对比可以更好地研究泥石流迹地植被自然恢复的过程和机制。虽然贡嘎山黄崩溜沟泥石流迹地植被恢复不是典型的原生演替过程,但是各个自然恢复阶段还是形成了恢复序列。黄崩溜沟泥石流迹地可对比观测不同恢复阶段的群落特征,是研究近百年天然林自然恢复的理想场所。
2 研究方法
2021 年在泥石流迹地样地上的不同植被恢复阶段分别布设10×10 m2和20×20 m2的样方3 个,调查各测树因子,按曲仲湘提出的划分标准对优势群的5 个立木级进行实验。对灌木层和草本层分别设立5×5 m2和1×1 m2的样方调查,记录种类和数量。恢复的前中期采用收获法测定生物量,后期采用解析木和生长方程计算生物量,对不同植被恢复阶段的立地作土壤剖面调查,按照土壤发生层次采集土样,测定土壤中N、P、K 等元素的含量。样地形成的年代,在近30 年按照观测资料确定,早于30 年的样地根据树龄推算。光合作用采用美国制造的 CI -301PS 和Li-6400 分别进行测定,生产力是通过测定光合速率并结合样地调查数据获得。
3 结果与分析
3.1 泥石流迹地植被恢复过程中群落动态
泥石流迹地形成的原生裸地由大量的沙粒和大小不一的石块、岩屑组成,有利于植物生长和土壤快速发育[11]。豆科草本植物蜀西黄芪(Astragalus souliei)分布广,固氮作用增强,迹地植被恢复速度快[12]。在迹地形成后的第4 年,先锋树种川滇柳(Salix rehderiana)和冬瓜杨(Populus purdomii)开始侵入迹地。在演替早期,大量的川滇柳侵入迹地,生长速度很快,冬瓜杨在初期比川滇柳生长慢一些。这一侵入过程持续10~15 年后结束。川滇柳在树龄约25 年时达到6.5 m 高,在这段时间里群落均是以川滇柳为主的川滇柳、冬瓜杨群落,随后冬瓜杨生长速度加快,逐渐在高生长上超过川滇柳,在群落中不断扩大覆盖度和土壤养分资源的占有率。川滇柳则逐渐失去这些资源,生长缓慢,死亡率增大,群落逐渐演替成以冬瓜杨为优势的冬瓜杨、川滇柳林。
峨眉冷杉在迹地形成后第5 年开始进入迹地,此时,由于土壤中的养分稀少,土壤微生物缺乏,对依靠外生菌根真菌吸收土壤养分的峨眉冷杉影响很大,所以峨眉冷杉在演替早期的存活率很低,生长十分缓慢。到迹地形成第8 年时,川滇柳和冬瓜杨形成幼树群落,土壤和地表微生物环境得到较大改善,峨眉冷杉侵入后存活率明显增大,生长较好。群落演替到冬瓜杨、川滇柳林阶段,经过冬瓜杨、川滇柳和其他灌木、草本的改造,土壤养分有了明显增加,土壤微生物环境得到改善,川滇柳种群进一步衰退,峨眉冷杉逐渐在树高、胸高断面等方面超过川滇柳,群落演替为冬瓜杨、峨眉冷杉林。糙皮桦(Betula utilis)出现较晚,在恢复后期对立地的养分循环发挥着一定作用,在峨眉冷杉成熟林中形成林下第二层。
随着环境条件不断改善,峨眉冷杉数量不断增加,对资源的占有率不断增大,逐渐对冬瓜杨构成很强的竞争压力,致使冬瓜杨进入衰退状态[13]。这一过程持续进行,直到峨眉冷杉树高超过冬瓜杨时,冬瓜杨种群密度已经大大减少,胸高断面积远远低于峨眉冷杉,群落演替为峨眉冷杉中龄林。在以后的近百年里,如果不发生灾害性干扰事件,峨眉冷杉将一直控制上层林冠,保持建群种地位。参照临近的海螺沟冰川退缩区植被原生演替序列,峨眉冷杉中龄林需要经过50 年后,才能逐步恢复到峨眉冷杉顶级群落,但是土壤还需要更大的时间尺度来恢复,形成顶极群落后群落进入较为稳定的状态(见表1)。
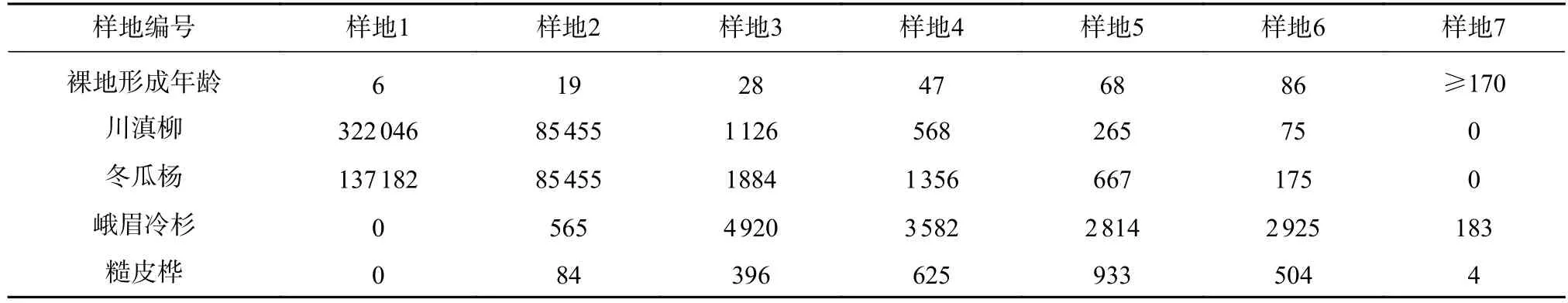
表1 泥石流迹地乔木优势种种群密度(trees·hm-2)Tab.1 Density of dominant tree populations in debris flow sites(trees·hm-2)
在不同植被恢复阶段由于群落组成发生变化,光能在群落中的分配发生了很大的改变[14]。迹地和林缘生长的草本和灌木净光合速率迅速升高,生长加速,但由于叶面积指数低,呼吸作用较强等因素,群落的生产力还是不高,光合作用种间和季节变化都很大。
灌木显脉荚蒾(Viburnum nervosum)、冰川茶藨子(Ribes glaciale)和美容杜鹃(Rhododendron calophytum)林缘的光合速率通常比林下的高出2—3 倍,它们之间的差异比草本植物小。草本双花蟹甲草(Sinacalia davidii)、鹿蹄草(Pyrola rotundifolia)、早熟禾(Poa annua)和扇羽阴地蕨(Botrychium lunaria),林缘的光合速率比林下的一般草本更高,通常高出5 倍左右,其净光合速率季节变化较大。测得林缘醉鱼草(Buddleja lindleyana)的瞬时光合速率最高,可达30 μmol·m-2·s 以上(见表2 和表3)。位于主林层的乔木层植物全年生长时间长,先锋植物净光合速率比峨眉冷杉高很多。乔木净光合速率通常比林下植物高1 倍以上(见表4)。
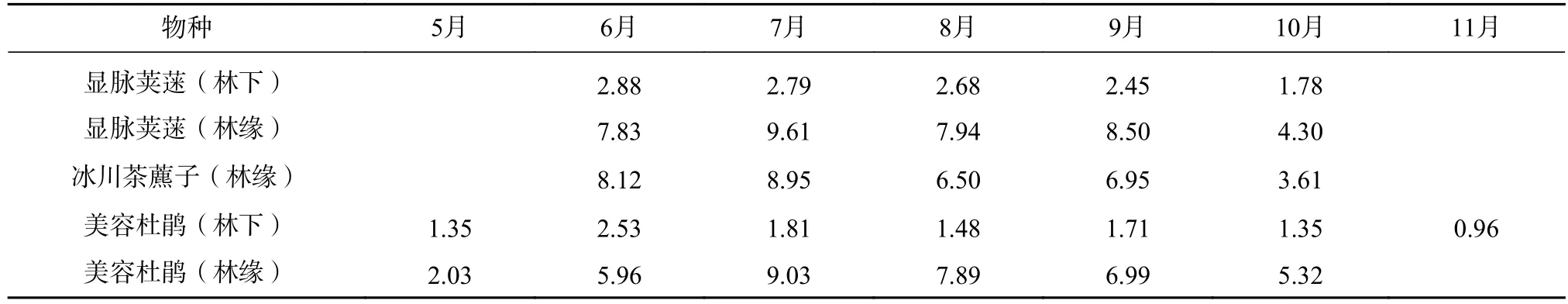
表2 主要灌木光合速率的年变化(μmol·m-2·s)Tab.2 Annual variation of photosynthetic rates of main shrubs(μmol·m-2·s)
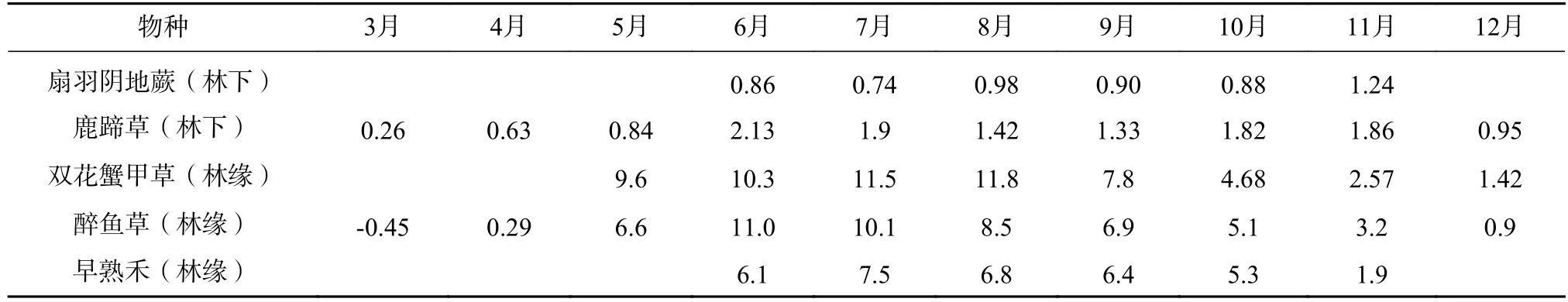
表3 主要草本植物光合速率的年变化(μmol·m-2·s)Tab.3 Annual variation of photosynthetic rate of main herbs(μmol·m-2·s)
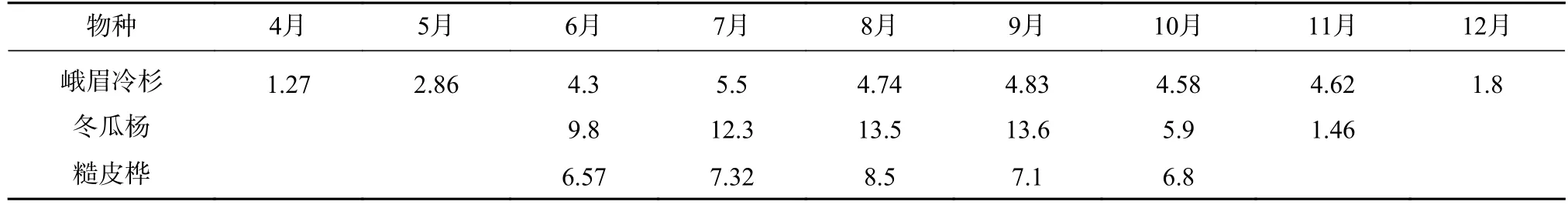
表4 乔木主要树种光合速率的年变化(μmol·m-2·s)Tab.4 Annual variation of photosynthetic rates of main tree species(μmol·m-2·s)
3.2 林分组成和结构以及生物量和生产力的变化
位于贡嘎山东坡海拔3 000 m 附近的峨眉冷杉成熟林(样地7)为地带性植被,在不同时期经泥石流破坏的迹地,现分别恢复到落叶阔叶幼树林(样地1)和小树林(样地4)以及峨眉冷杉中龄林(样地6),将上述4 类样地的生物量和生产力进行比较,可获得生态系统自然恢复的一般特征。
原生裸地环境变化大,缺乏营养元素,先锋群落结构单一,生物量和生产力都不高群落结构单一,生物多样性不高,植物光能利用效率最低,草本植物比较发达[15]。植被经47 年恢复,生物多样性提高,植物生长季光能利用效率有所升高,在形成郁闭的小树林后,生物量和生产力显著提高,林下灌木和草本生长良好,种间竞争加剧,群落组成在这一时期虽然变化最大,乔灌草层次结构趋于合理。峨眉冷杉中龄林种内竞争加剧,自疏加强,建群种数量极不稳定,年龄结构单一,生物多样性降低。在植被恢复的各个阶段,峨眉冷杉成熟林生物多样性最高,生物量和生产力也最高,光能利用效率最高,生物量和生产力在林分中分配合理(见表5)。
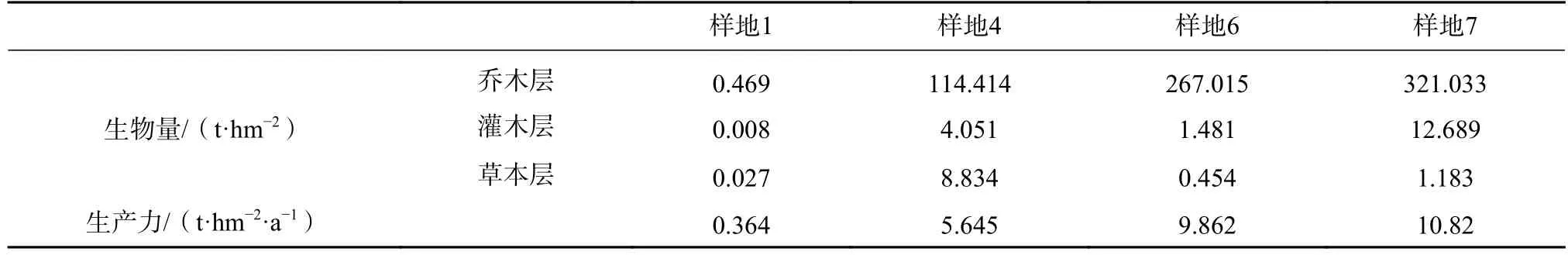
表5 泥石流迹地不同恢复阶段群落的生物量和生产力Tab.5 Community biomass and productivity at different restoration stages in debris flow sites
3.3 泥石流迹地的草本植物
对泥石流迹地7 个恢复阶段中的4 个群落的草本植物进行调查,共有约34 科59 种,以峨眉冷杉中龄林数目最少,有11 科16 种;冬瓜杨与峨眉冷杉林混交林和峨眉冷杉成熟林最高,分别为21 科32 种和20 科33 种(见表6)。虽然在一些恢复阶段草本的生物量和生产力不高,但是,在泥石流迹地的植被恢复过程中,草本总物种数表现为由少到多的增加趋势[16]。
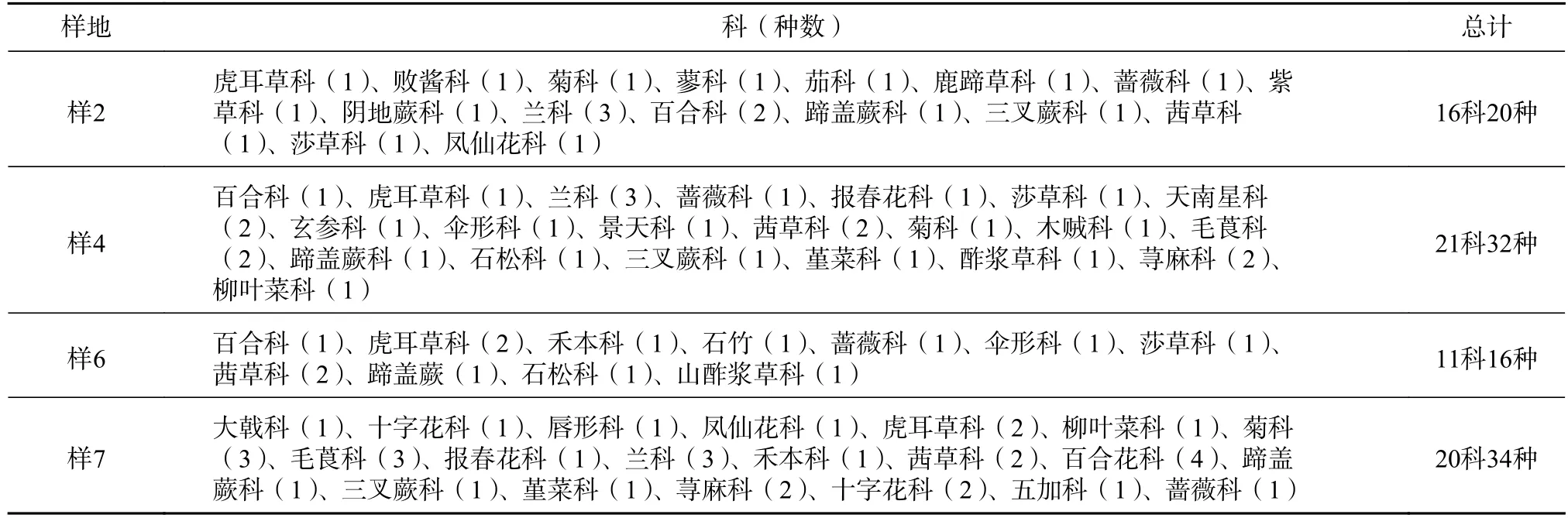
表6 泥石流迹地不同恢复阶段群落草本分布Tab.6 Distribution of herbaceous community at different restoration stages in debris flow sites
4 结果与讨论
在高山峡谷区泥石流扇上发生的是原生演替,植被自然恢复过程特点明显:整个演替过程比较简单,最初草本群落阶段豆科植物的固氮作用至关重要,演替初期、中期和后期共有三个建群种,分别是川滇柳、冬瓜杨和峨眉冷杉;顶极群落建群种峨眉冷杉自迹地形成后的第5 年即开始进入,从邻近原始峨眉冷杉林传播的种子能萌发出幼苗,但成活率很低,在迹地形成第20 年以后群落恢复速度加快,迹地形成后第80 年即能恢复到峨眉冷杉中龄林;在整个演替过程中,建群种数量变化在减小,从原生裸地开始,自然恢复到顶级群落需要150 年以上,但是形成地带性土壤需要更漫长的时间[17];草本植物物种组成和密度随演替进程变化明显,生物多样性变化较大。
联合国已将2021—2030 年定为“生态系统恢复的十年”。森林恢复方式的选择上,倡导天然林自然恢复。天然林的恢复需要对自然恢复过程中植被和环境变化的进行连续观测,探索恢复过程和机理,才能进行植被恢复技术集成与模式优化筛选,构建不同目的的物种配置模式,建立生态系统保护和管理的指标体系,确保群落更加有序和稳定,有效扭转生态系统退化趋势,为可持续发展做出贡献[18,19]。
森林恢复还需要关注生物多样性的保护,需要衡量森林恢复能在多大程度上实现其生态系统服务的目标。与结构简单的人工林相比,天然林可以更好地支持生物多样性保护和实现地表碳贮存、土壤保持、水源涵养的生态系统服务[20,21]。对长时间序列的天然林恢复过程的研究,有助于探索不同恢复目标之间的协同与权衡关系,为制订森林恢复的政策提供科学支持。
