Bɑcillus sp.T124对六价铬的生物还原
2022-05-29姜毅汤鼎周宇诚丁聪聪赵兴青柴育红
姜毅,汤鼎,周宇诚,丁聪聪,赵兴青*,柴育红,2
(1.常州大学环境与安全工程学院,江苏 常州 213164;2.常州大学怀德学院,江苏 靖江 214500)
由于在工业中的广泛使用和排放,铬(Cr)成为了一种常见的环境污染物。Cr 在环境中以多种价态存在,其中最常见和最稳定的是Cr(Ⅵ)和Cr(Ⅲ)。据报道,Cr(Ⅵ)较Cr(Ⅲ)毒性更高,因为Cr(Ⅵ)具有很高的稳定性和细胞通透性。因此,Cr(Ⅵ)已被许多国家指定为首要环境污染物之一。而Cr(Ⅲ)是维持人体新陈代谢和体内稳态所必需的,并且Cr(Ⅲ)具有较低的毒性、迁移率和生物可利用度,这是因为其容易在环境中形成不溶的氢氧化物或氧化物。因此,Cr(Ⅵ)污染的修复策略主要集中在将Cr(Ⅵ)还原为Cr(Ⅲ)。
近些年,已研究出多种将Cr(Ⅵ)转化为Cr(Ⅲ)的方法,包括物理化学修复以及生物修复。与生物修复相比,物理化学法快速有效,但存在操作复杂、设备昂贵和二次污染等缺点。而生物修复由于成本低、环境友好等突出优势而备受关注。
现已经鉴定并分离出多种具有Cr(Ⅵ)去除能力 的 微 生 物,如W4、sp. M6、LY10、、sp.GSKRLMBKU-03等。细菌还原Cr(Ⅵ)的机理主要有两种:一是间接还原,即通过细菌的代谢产物还原Cr(Ⅵ);二是直接还原,即通过细菌铬酸盐还原酶的酶促反应将Cr(Ⅵ)还原为Cr(Ⅲ)。ACKERLEY等研究发现大肠埃希氏菌可产生一种氧不敏感性的酶——NfsA,这种酶可将Cr(Ⅵ)直接还原为Cr(Ⅲ)。还 有 研 究 发 现MR-1和sp.能 将Cr(Ⅵ)还 原为Cr(Ⅲ)。然而,Cr(Ⅵ)生物修复过程的有效性主要取决于还原的最终产物,先前的研究很大程度上忽略了Cr(Ⅲ)产物的形成过程及对最终产物的表征分析。
还原产物的表征至关重要,我们不仅需要了解还原机理,还需要探究还原产物的形态和种类。耐Cr菌株sp.T124分离自长期受铬污染的土壤,本研究对该菌株还原Cr(Ⅵ)的效率和机理进行探究,并在最佳条件下评估了该菌株对Cr(Ⅵ)的还原能力。通过扫描电镜-能谱仪(SEM-EDS)分析菌株和还原产物的形态,利用X射线衍射(XRD)和拉曼光谱分析还原产物的构成,利用傅里叶红外光谱(FT-IR)和X射线电子能谱(XPS)分析还原产物的官能团变化和元素构成,以探索菌株的性质和还原产物的特征,从而了解Cr(Ⅵ)的还原机理。
1 材料与方法
1.1 Cr(Ⅵ)还原菌株
从江苏某电镀工厂周边收集的Cr(Ⅵ)污染土壤中分离出耐受Cr(Ⅵ)且表现出最高Cr(Ⅵ)还原能力的菌株T124,16S rDNA 基因序列分析将其鉴定为芽孢杆菌,命名为sp.T124。将菌株序列提交至GenBank 登记获得登录号为MN511807。LB 液体培养基组成为:鱼粉蛋白胨10 g·L,酵母浸膏5 g·L,NaCl 5 g·L。将KCrO溶解于去离子水中以制备Cr(Ⅵ)的储备溶液(10 g·L)。完全抑制细菌生长的最低金属浓度被认为是最低抑制浓度(MIC),菌株T124的MIC值被确定为300 mg·L。
1.2 Cr(Ⅵ)浓度的测定
向含100 mg·LCr(Ⅵ)的95 mL LB 液体培养基中加入5 mL 活化12 h 后的菌液(OD=1),将其置于37 ℃的恒温振荡箱中以180 r·min的转速培养48 h。设置无细胞的培养基作为对照,以排除生物作用。所有实验均设置3 个平行。分别在第0、2、4、6、8、10、12、24 h 和48 h 时取出2 mL 样品置于比色管中。样品中的Cr(Ⅵ)浓度通过1,5-二苯碳酰二肼法测定:将0.2 g 1,5-二苯碳酰二肼溶解在50 mL 丙酮中,随后加入12.5 mL 85%的磷酸、12.5 mL 95%的浓硫酸以及50 mL 的去离子水以获得显色剂;将2 mL显色剂添加至取出的2 mL 待测样品中并用无菌水稀释至50 mL,在分光光度计上540 nm 处测定混合溶液中的Cr(Ⅵ)浓度。
1.3 菌株T124还原Cr(Ⅵ)的影响因素
为了对菌株T124还原Cr(Ⅵ)的环境条件进行优化,在不同碳源和不同共存离子的条件下测定其对Cr(Ⅵ)的还原率。不同碳源(葡萄糖、果糖、蔗糖、乳糖、乙酸钠和甘露醇,15 mg·L,pH 7.0)下菌株对Cr(Ⅵ)的还原实验在Minimal salt medium(MSM)培养基(NaHPO6.0 g·L,KHPO3.0 g·L,NaCl 0.5 g·L,MgSO·7HO 0.246 g·L,CaCl0.5 g·L)中进行,在含有不同碳源的培养基中分别加入100 mg·L的Cr(Ⅵ)和5 mL 活化后的菌液,将培养基置于恒温振荡箱(37 ℃、170 r·min)培养2 d 后取出测定Cr(Ⅵ)浓度。

1.4 SEM-EDS和XRD分析
通过离心(8 000 r·min、20 min)收集反应完全后的细胞和沉淀物,然后用磷酸盐缓冲液洗涤3 次。冷冻干燥后的样品研磨后喷金即可上机。利用扫描电子显微镜&能谱仪(SEM & EDS,日本电子株式会社JSM-6360 LA型)对菌株的细胞形态和矿物沉淀的结构形态进行观察,并且利用能谱仪(EDS)对细胞表面成分和矿物沉淀成分进行详细的元素分析。
将离心后的沉淀物冷冻干燥并研磨成粉末,然后与光谱纯KBr 以1 mg 样品与100 mg KBr 的比例混合压片。利用D-MAX2500 型X-射线衍射仪(XRD,日本理学)对沉淀物中的矿物成分进行测定。测试参数为电压40 kV、电流40 mA,2角度范围为10°~90°,扫描速率为0.5°·min。使用MDI Jade 软件(版本6,Materials,Data,Inc,美国)对XRD数据进行分析。
1.5 拉曼光谱分析
利用LabRAM HR 型拉曼光谱仪(Horiba-Jobin,日本)分析与细菌细胞相关联的还原产物。制样方法同1.4,将2 mg样品粉末直接粘在双面碳导电胶上,测试参数为激发波长532 nm,分辨率为(2±1)cm,测量范围为250~3 000 cm,使用最大倍数(50 倍)重复收集。
1.6 FT-IR和XPS分析
将反应完全后的样品以10 000 r·min离心20 min,得到的细胞重悬于100 mL 0.05 mol·L的磷酸盐缓冲液中(PBS,pH 7.0),充分搅拌,形成均匀的细胞悬液,然后再以10 000 r·min离心15 min。重复上述步骤3 次。将离心后的沉淀冷冻干燥并研磨成粉末,然后与光谱纯KBr 以1 mg 沉淀粉末与100 mg KBr 的比例混合压片,在10 t·cm下静压2 min。采用Perkin-Elmer-Spectrum One 型傅里叶变换红外光谱仪(Nicolt A vatar 370,美国尼高力仪器公司)收集波长范围为4 000~400 cm处的红外光谱数据,扫描准确度为4 cm。
利用PHI-5000 Versaprobe Ⅲ型X 射线光电子能谱仪(XPS,日本)分析沉淀物中Cr的价态。测试参数为扫描周期2 周期,扫描次数5 次,范围200µm,分析仪模式为55 eV和280 eV,能量步长为0.05 eV。
1.7 统计分析
采用SPSS 23 进行数据统计分析,采用单因素方差分析(One-way ANOVA)和最小显著差异检验(LSD)确定变量之间的差异显著性(<0.05)。采用Origin 2020绘图。
2 结果与讨论
2.1 Bɑcillus sp.T124还原Cr(Ⅵ)
如图1a 所示,在接种细菌后的12 h 内,溶液中的Cr(Ⅵ)浓度快速降低,接种12 h时的Cr(Ⅵ)去除率达到了26.9%;在12~48 h 内去除率仍在升高,但是速率降低,营养盐被消耗导致的细菌生长量和活性降低可能是Cr(Ⅵ)还原速率降低的原因之一。有研究表明,由碳源氧化产生的电子原本用于生物合成,但当有Cr(Ⅵ)存在时,电子会被用于还原Cr(Ⅵ),从而减慢了生长速度。另外从图1b中可以明显看出反应48 h后紫色明显变淡,这表明Cr(Ⅵ)浓度明显降低,菌株对Cr(Ⅵ)的还原效果显著。XU 等在对小球藻去除Cr(Ⅵ)的研究中发现,在最初的30 min内小球藻快速去除Cr(Ⅵ),随后去除速率趋于平稳。DAS 等也发现,在存在的45 h 内,初始Cr(Ⅵ)浓度越低,其还原速率越高,在Cr(Ⅵ)浓度为10 mg·L时的还原速率快于Cr(Ⅵ)浓度为50 mg·L和100 mg·L时的情况。当Cr(Ⅵ)初始浓度为100 mg·L时,sp.T124 具有较高的Cr(Ⅵ)去除能力,在48 h 内对Cr(Ⅵ)的去除率为43.2%。在对照组中Cr(Ⅵ)浓度从101.32 mg·L降低至87.72 mg·L,这可能是Cr(Ⅵ)在弱碱性条件下被还原和设备仪器的误差导致的,因此可以忽略非生物因素对Cr(Ⅵ)去除的影响,说明sp. T124 可以从溶液中去除Cr(Ⅵ)。

图1 Bɑcillus sp.T124在48 h内对Cr(Ⅵ)的还原和还原前后溶液颜色的变化Figure 1 Reduction of Cr(Ⅵ)by Bɑcillus sp.T124 within 48 h and change of solution color before and after reduction
2.2 碳源和共存离子对Bɑcillus sp.T124 还原Cr(Ⅵ)的影响
菌株生长过程中必需的物质之一是碳源,不同碳源可能会对菌株还原Cr(Ⅵ)有不同影响,在MSN 介质中研究了6 种不同碳源(葡萄糖、果糖、蔗糖、乳糖、乙酸钠和甘露醇)作为电子供体对菌株T124还原Cr(Ⅵ)的影响,结果如图2a 所示。在接种sp. T124 的2 d 后,葡萄糖、果糖、蔗糖、乳糖、乙酸钠和甘露醇作为电子供体对Cr(Ⅵ)的去除率分别为53.50%、76.59%、52.24%、10.28%、53.50%和19.24%(<0.05),表明在这6 种碳源中,果糖是最合适的电子供体。葡萄糖、蔗糖以及乙酸钠作为电子供体时的还原效果也较好,乳糖和甘露醇则不适合作为菌株还原Cr(Ⅵ)的电子供体。TAN 等发现果糖作为电子供体时,sp. CRB-B1 对Cr(Ⅵ)的去除率最高,葡萄糖和蔗糖次之,其结论与本实验相同。而DAS等在利用还原Cr(Ⅵ)时则发现葡萄糖是最合适的碳源,其次是果糖。DEY等发现利用甘油作为电子供体的效果最好,而蔗糖作为电子供体的效果最差。这说明不同的菌株对碳源的适应程度不同,更偏向于选择果糖或葡萄糖作为电子供体。
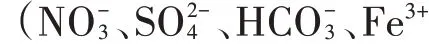

图2 不同碳源和不同共存离子对Bɑcillus sp.T124还原Cr(Ⅵ)的影响Figure 2 Effects of different carbon sources and co-existing ions on the reduction of Cr(Ⅵ)by Bɑcillus sp.T124
2.3 SEM-EDS和XRD分析
细菌细胞的SEM-EDS分析结果见图3。在5 000倍放大倍数下,不含Cr(Ⅵ)的细菌细胞为长杆状,表面平坦且无孔(图3a)。EDS的结果表明,未经Cr(Ⅵ)处理的细菌样品中不含Cr(Ⅵ)。如图3b 所示,经Cr(Ⅵ)处理后的sp.T124 的细胞形态无明显变化,没有破裂和穿孔,这表明浓度为100 mg·L的Cr(Ⅵ)不会对细菌产生严重的毒害作用,该菌株可以耐受高达100 mg·L的Cr(Ⅵ);另外还观察到一些无定形物质结垢附着在细胞表面,其可能是Cr(Ⅵ)生物还原过程中的还原产物。本研究进一步对附着在细胞表面的无定形物质进行了EDS分析,结果在无定形物质中检测到了元素Cr。由于EDS 检测的X 射线能量很高,并且可以穿透薄细菌细胞样品,所以在消除其他干扰成分后,细胞表面的无定形颗粒物可以被视为含Cr物质。SEM-EDS证实了检测到的细胞表面有含Cr化合物的存在。LI等在用SEM-EDS分析经过Cr(Ⅵ)处理48 h 的W1 细胞时发现,浓度为200 mg·L的Cr(Ⅵ)不会对菌株产生毒害作用,菌株的细胞形态没有变化,并且还发现细胞表面的无定形颗粒为含Cr 物质。而有些细胞会因为Cr 的存在而发生形态变化,如CHEN 等在 分析了AR8 的细胞形态后发现,未经Cr(Ⅵ)处理的细胞具有规则光滑的表面,而经Cr(Ⅵ)处理后的细胞是不规则、凹陷、起皱的形态,并且带有一些穿孔。sp.T124 在经过高浓度的Cr(Ⅵ)处理后,细胞形态未有明显变化,这表明sp. T124 具有Cr(Ⅵ)的高耐受性。
为了确定Cr(Ⅵ)还原过程中Cr(Ⅲ)的形成,通过XRD 扫描了经过Cr(Ⅵ)处理的细菌细胞,结果如图3c 所示。经过Cr(Ⅵ)处理生成的产物在2为21.3°、31.66°和38.44°处显示出3 个明显的CrO沉淀峰,在使用标准JCPDS(参考代码01-084-0312)对比后证实了CrO的存在,故可认为产物中存在Cr(Ⅲ),且其是由Cr(Ⅵ)被sp.T124部分还原形成的。CHEN 等在利用XRD 扫描经Cr(Ⅵ)处理的细菌细胞后发现,在2为24.54°、33.66°、36.26°、39.83°、41.55°、44.27°、50.31°和54.95°处也出现了明显的CrO沉淀峰。DHAL等在研究从铬铁矿土壤中分离出的sp.还原Cr(Ⅵ)时也发现了相似的Cr(Ⅲ)沉淀。MOHITE 等报道了介导的Cr(Ⅵ)到Cr(Ⅲ)纳米颗粒的生物转化,并通过XRD分析得到了证实。而在KARTHIK等的报道中发现,strain G12处理Cr(Ⅵ)后,Cr 的峰主要出现在了27.4°、31.7°、45.8°和53.9°,对应于这些峰的晶体组成与CrCl相似,因此推断Cr(Ⅵ)确实被还原为了Cr(Ⅲ),且Cr(Ⅲ)与Cl复合在细菌表面形成了CrCl。

图3 Cr(Ⅵ)处理前(a)后(b)Bɑcillus sp.T124的SEM-EDS图像(×5 000倍)和Cr(Ⅵ)处理后(c)Bɑcillus sp.T124的XRD图像Figure 3 SEM-EDS patterns(×5 000 times)of Bɑcillus sp.T124 of before(a)Cr(Ⅵ)treated cells and after Cr(Ⅵ)treated cells(b)XRD pattern of Bɑcillus sp.T124 of Cr(Ⅵ)treated cells(c)
2.4 拉曼光谱分析
拉曼光谱法是表征涉及分子结构和重金属基络合物的强大工具,因此为了检测细菌还原后Cr 可能的价态,通过拉曼光谱分析了经Cr(Ⅵ)处理后的细菌细胞。如图4 所示,Cr(Ⅵ)处理后的细胞及其相关产物在600 cm处显示出清晰的特征峰,该峰与标准Cr(Ⅲ)相对应。拉曼光谱分析的结果表明Bɑcillus sp. T124 确实还原了Cr(Ⅵ),并且细胞固定的Cr 为Cr(Ⅲ)。WANG 等利用共振拉曼光谱法和表面增强拉曼光谱法在单细胞水平上分析了Shewɑnellɑ oneidensis 对Cr(Ⅵ)到Cr(Ⅲ)的 影 响。RAVINDRANATH 等用表面增强拉曼光谱法证实了S. oneidensis MR-1 细胞内Cr(Ⅵ)的减少。KARTHIK 等也 报 道 了Cellulosimicrobium funkei AR6 在经Cr(Ⅵ)处理后,细胞在拉曼图谱中600 cm附近区域显示出强烈的特征峰。对Bɑcillus ɑmyloliquefɑciens 的Cr(Ⅵ)还原产物(沉淀物)进行拉曼光谱分析发现,在600 cm附近区域存在Cr(Ⅲ)峰。因此,拉曼光谱的分析结果进一步证实了Bɑcillus sp.T124将Cr(Ⅵ)还原为Cr(Ⅲ)。
2.5 FT-IR和XPS分析

XPS 是研究Cr(Ⅵ)还原过程中sp. T124细胞中的官能团和元素Cr 化合价的有效分析方法。如图5b 所示,从sp. T124 的XPS 调查光谱中监测到3 个主要峰,分别位于286.31、400.63 eV 和532.83 eV 处,分别代表C 1s、N 1s 和O 1s。此外,在579 eV和48 eV处发现了两个弱峰,分别代表Cr 2p和Cr 3p。如图5c 所示,在587.63 eV(Cr 2p)和577.50 eV(Cr 2p)附近出现了两个不同的峰(从Cr 光谱获得),表明Cr(Ⅵ)还原产物的化合价主要为Cr(Ⅲ)。同时也证明sp.T124 是通过还原而非吸附的方式去除了环境中的Cr(Ⅵ)。YAO 等在利用XPS 分析CY-1 还原Cr(Ⅵ)和Hg(Ⅱ)的产物化合价时得出了与本研究相似的结论。BAI等在利用XPS分析经过Cr(Ⅵ)处理的时发现Cr 2p和Cr 2p存在分裂,形成了不同的还原产物,这证明了使Cr(Ⅵ)减少的途径是还原而非吸附。以上结果进一步证明sp. T124 对Cr(Ⅵ)的去除是通过生物还原这一途径实现的,并且Cr的还原产物主要为Cr(Ⅲ)。
2.6 Bɑcillus sp.T124对Cr(Ⅵ)的去除机制
由本课题组前期研究可知,Cr(Ⅵ)还原的过程主要在细胞外进行。对于特定的还原位点,菌株的每个组分均具有还原Cr(Ⅵ)的能力,但是每个组分的单独还原能力完全不同。现已知的Cr(Ⅵ)去除机制有生物吸附、细胞内积累和生物还原,其中由还原酶在细胞壁膜上介导的生物还原是sp.T124去除Cr(Ⅵ)的主要途径,sp. T124 分泌的还原酶大多数在细胞外实现对Cr(Ⅵ)的还原,可能是Cr(Ⅵ)的毒性限制了细胞内还原的过程。SILVER等发现在利用硫酸盐的生物细胞中,Cr(Ⅵ)是通过硫酸盐转运通道的膜被吸收。另外有研究表明,细胞外还原的Cr 主要是水溶性有机Cr(Ⅲ),这些Cr(Ⅲ)无法渗透进入细胞内,而部分在细胞内被储存于细胞质中的Cr(Ⅲ)是由Cr(Ⅵ)还原而来。AHEMAD等也发现Cr通过质粒转运至细胞质,慢慢地在细胞内积累,细胞内的个别成分(有机酸、核酸和氨基酸等)可作为电子供体,而Cr(Ⅵ)作为电子受体接受电子后还原为Cr(Ⅲ)。总体来说,细胞表面还原的Cr(Ⅵ)通常有一部分会被释放到上清液中或者被官能团所吸附,因此细胞外Cr(Ⅵ)的还原对菌本身起到了间接的保护作用,并且在Cr(Ⅵ)的转移中不需要消耗额外的能量。
通常不同细菌对Cr(Ⅵ)的还原机制有所差别,影响还原的因素也有所不同。例如,有些细菌会利用Cr(Ⅵ)作为最终的电子受体,有些细菌则会利用分泌的可溶性酶将Cr(Ⅵ)还原为Cr(Ⅲ)。此外,CHEN等指出蜡状芽孢杆菌对Cr(Ⅵ)的还原与细胞壁膜有关,在本研究中静止细胞较渗透细胞对Cr(Ⅵ)的还原量高出10.5%,也验证了这一结论,说明Cr(Ⅵ)的还原受限于细胞壁膜的通透性。肖伟等也发现细胞壁膜的通透性可以对Cr(Ⅵ)还原酶的作用产生影响。XRD 和拉曼分析进一步证明了在细胞膜上sp.T124将Cr(Ⅵ)还原为Cr(Ⅲ),且还原产物为CrO,由FTIR 的结果得出烷基、羧基以及多糖参与了Cr(Ⅵ)的还原,减轻了Cr(Ⅵ)对细菌细胞的毒性。XPS 分析为证明Cr(Ⅵ)的还原产物主要为Cr(Ⅲ)提供了更多的依据,并且说明sp.T124对Cr(Ⅵ)的去除途径是生物还原而非吸附,此结果和YAO等得到的反应机制一致。
3 结论
(1)sp.T124 能有效去除Cr(Ⅵ),在48 h内可去除43.2%的Cr(Ⅵ)。
(2)sp. T124 对Cr(Ⅵ)的还原效果主要受碳源和共存离子的影响。添加共存离子对Cr(Ⅵ)的还原产生了抑制作用,其中HCO的抑制效果最为明显。
(3)表征分析结果进一步证实了sp.T124 对Cr(Ⅵ)的去除主要是通过生物还原这一途径实现的,并且Cr的还原产物主要为Cr(Ⅲ)。
