茄果类蔬菜单性结实研究进展
2022-05-05姜政杜丽慧张宇申磊杨旭
姜政 杜丽慧 张宇 申磊 杨旭
摘 要:茄果类蔬菜单性结实可以改善果实品质、增强抗性、提高产量等。研究茄果类蔬菜单性结实对改良品种和提高产品品质有重要意义。对茄果类蔬菜(番茄 Lycopersicon esculentum、茄子 Solanum melongena L.、辣椒 Capsi cum annuum L.)单性结实的种质资源、生理指标、诱导因素、差异基因和遗传机制等方面的研究进行了综述,并对其今后的发展和研究方向进行了展望。
关键词:茄果类蔬菜;单性结实;种质资源;内源激素;调控基因;遗传机制
中图分类号:S641 文献标志码:A 文章编号:1673-2871(2022)03-001-08
Abstract:Parthenocarpic fruit set can improve fruit quality, resistance and yield of solanaceous vegetable. Understandingof parthenocarpy of solanaceous vegetable is of great significance to improve varieties and product quality. This articlereviews the research on genetic resources, physiological indexes, influencing factors, regulatory genes and genetic mecha nism of parthenocarpic fruit set in solanaceous vegetables, Lycopersicon esculentum, Solanum melongena L. and Capsi cum annuum L. The research direction of solanaceous parthenocarpic fruit set is also outlined.
Key words:Solanaceous vegetable; Parthenocarpy; Germplasm resource; Endogenous hormone; Regulatory gene; Geneticmechanism
茄果类蔬菜是指茄科以浆果作为食用部分的蔬菜作物,一般包括番茄、茄子、辣椒,是我国蔬菜栽培生产中最重要的种类之一[1]。茄果类蔬菜喜温暖,在低温(<15 ℃)和高温(>28 ℃)环境下均会致其坐果不牢,易发生落花落果[2]。茄果类蔬菜对光周期要求不严,根系较为发达,具有一定的抗旱能力,需水需肥量大[3]。单性结实是指植物不经过受精就可以结出正常果实的现象,一般是植物在自然演化中发生变异而来[4]。单性结实根据其发生条件一般可分为兼性单性结实和专性单性结实[5],兼性单性结实是指其单性结实性会受环境等多方面因素的共同作用,无法稳定遗传。目前,在茄果类蔬菜单性结实的研究中使用的大都是兼性单性结实材料。专性单性结实是指其单性结实性不受环境的影响,仅由基因所控制,且可以稳定遗传。这种材料仅能靠无性繁殖的方法培育,目前尚未发掘,是今后茄果类蔬菜育种的重要方向之一。单性结实的植物因没有经过受精作用,其产生的果实为无籽果实。诸多研究表明,茄果类蔬菜单性结实可显著改善果实品质,克服落花落果障碍[6]。单性结实的番茄无籽果实能提升品质,改善口感[7],并且可以减少畸形果,提高果实耐贮性[8]。单性结实品系的茄子可以提高抗寒性[9],减少由低温造成的落花落果问题[2],提升果实品质,对早期增产[10]和提高经济效益具有重要意义。辣椒单性结实可显著提升低温下的坐果率,增加早期产量等[11]。可见,研究茄果类蔬菜单性结实的相关特性对改良现有品种、提升产品的商品价值具有重要意义。笔者对近年来国内外有关茄果类蔬菜单性结实的资源、影响因素、分子机制、遗传机制进行综述,并提出展望,以期为今后单性结实的进一步研究和茄果类蔬菜的品种改良提供参考。
1 茄果类蔬菜单性结实的种质资源
有关番茄的单性结实资源大部分是通过远缘杂交的手段得到,少部分是经过自然突变而来[12]。有研究指出,目前被广泛研究的番茄单性结实材料主要有 3 个,即含有 pat 基因的 Montfavet 191、含有pat2 基因的 Severianin 和含有 pat3/pat4 基因的RP75/79[13]。其中,含有 pat 基因的材料為雌性不育型,含有 pat3/pat4 基因的材料本身存在一定的缺陷,二者在生产上难以应用[14],因此在实际研究中多用含有 pat2 基因的材料。
有关茄子单性结实资源的研究较早,1998 年中国农业科学院蔬菜花卉研究所以单性结实的圆茄为基础材料,经过多代筛选后得到多个稳定的种质资源[15]。Restaino 等[16]使用激素处理的方法,获得具有抗低温胁迫的单性结实材料。目前,用于茄子单性结实研究较多的资源有单性结实品系 QZ/4[17],圆茄自交系中选育得到的 D-10[18]、D-11[19]、D-21[20]品系等。
有关辣椒单性结实的研究尚少,所得资源也较为匮乏。日本学者 Ichiro Honda 等[21]在研究辣椒果实大小和单性结实时使用了单性结实性品系CNPH 2622,并发现了具有高度单性结实性的新品种 Shishitoh。张金凤[11]使用高代自交系辣椒 07-33作为单性结实材料进行坐果习性、生理特征和遗传规律的研究。
茄果类蔬菜单性结实的相关研究虽已进行多年,但种质资源仍不丰富。发掘更丰富的株系材料是进行单性结实遗传机制、分子机制研究的基础,有利于进一步开发单性结实的分子标记,从而利用单性结实的调控基因选育新品种。
2 茄果类蔬菜单性结实的生理指标
诸多研究已经证实,茄果类蔬菜单性结实能获得有利于提升其商品性的优良性状。王静等[22]对单性结实茄子品系 D2 进行研究,观察其果实可以正常生长发育,果实可溶性糖、可溶性蛋白质含量测定的结果表明,单性结实品系与非单性结实品系的果实在生长发育过程中可溶性糖和可溶性蛋白质含量的变化表现出高度的一致性。这说明单性结实品系的果实不会发生性状退化,可大面积栽培,具有应用价值。潘秀清等[8]对茄子单性结实品系D-11 的研究发现,单性结实的果实具有更好的坐果习性,且果实质量增加速度更快,平均单果质量更大。另有其他学者的研究亦得出此结论[23],推测原因是单性结实品系在开花之前已开始积累有机物,而并非常规品种授粉后快速积累,这一特点可使其早期产量提高,可在冬季或早春的设施栽培中得以应用。成玉富等[17]对茄子单性结实品系 QZ/4 的研究发现,在相同的栽培條件下,单性结实品系的平均坐果率比非单性结实品系高 6.37%;在不适于茄子生长的高温及低温胁迫下,单性结实品系的花粉萌发率更高,可见单性结实品系 QZ/4 的花粉粒中贮存了更多的酶和代谢物质,在对抗温度胁迫时具有更好的耐受性。在其他茄果类蔬菜单性结实品系的生理指标研究中也得出了相似的结论。Mao等[24]在对番茄单性结实的研究中发现,单性结实品系的坐果率更高,耐低温性和耐贮性更好。王利英等[25]研究番茄果实性状时发现,单性结实品系的畸形果率低于非单性结实品系,可得到商品性更为优良的产品。张金凤[11]对辣椒单性结实的研究中发现,单性结实株系 07-33 具有更好的耐低温性,营养积累更快,果实内 IAA 和 ABA 含量有明显的升高。可见茄果类蔬菜的单性结实品系更耐低温,且前期生长发育更快,积累的营养物质更多,坐果率更高,在生产上应用单性结实品系可提高产品的经济效益。故培育单性结实品系是茄果类蔬菜的重要育种方向,具有很高的研究价值。
3 茄果类蔬菜单性结实的诱导因素
3.1 内源激素
3.1.1 生长素(IAA) 生长素可以调节植物生长速率,促进种子萌发,在植物生长发育的不同阶段均起到一定的调控作用。研究发现,茄子单性结实品系的子房的生长依赖于高内源生长素水平的维持,这说明内源生长素可能在茄子单性结实过程中起着重要作用[26]。武彦荣等[27]的试验得出单性结实品系的子房内源生长素含量在开花时比非单性结实品系高出 4 倍,也印证了这一点。李艳玮等[28]的研究发现,在低温环境下,非单性结实品系的茄子子房中生长素含量低于单性结实品系,推测是由于低温不利于茄子生长素的合成,单性结实品系由于在生长早期就已积累了大量的生长素,所以受低温的影响较小。另外,张伟春等[29]的研究也得到了相同的结论。在番茄单性结实研究中,Mapelli 等[30]发 现,单性结实品系的番茄子房中开花前的生长素含量比非单性结实品系约高 3 倍,且生长素积累速度也高于非单性结实品系,Hazra 等[31]对番茄单性结实品系 pat2 的研究也得到了同样的结论。此外,张金凤[11]在对辣椒单性结实的研究中同样发现其子房内源性生长素的含量显著高于非单性结实品系。由此可见,生长素在不同的茄果类蔬菜单性结实中所发生的含量变化基本一致,可以推测影响生长素合成、运输和发挥作用的相关基因和化学物质都有可能影响茄果类蔬菜的单性结实性。此外,单性结实品系出现生长素水平较高的原因可能是其子房没有受精作用的刺激,会在前期自发的大量合成生长素,以保证果实的正常生长发育。
3.1.2 赤霉素(GA3) 赤霉素可刺激植物生长,具有促进植物器官发育、促进开花结果的作用。有研究报道称赤霉素可刺激子房发育,促进单性结实果实的膨大生长。Fos 等[32]的研究发现,番茄雌蕊柱头经过赤霉素处理后,其子房细胞的生长和分裂速度加快,更容易诱导出单性结实。Francesco 等[33]研究发现,兼性单性结实番茄的子房中赤霉素含量高于野生种。张金凤[11]在对辣椒的研究中也得到了相似的结论。但是在茄子单性结实的研究中关于赤霉素的作用尚存争议。张伟春等[29]研究发现,单性结实品系茄子的子房赤霉素含量变化与生长素类似,在开花前后保持较高的水平,并在同一发育时间下高于非单性结实品系。但武彦荣等[27]研究认为,内源性赤霉素对茄子单性结实的影响不大,不同品系间激素含量的差异可能是源于基因型的差异。此外,李艳玮等[20]的研究发现,赤霉素的同系物GA4在茄子果实发育中主要对温度响应,且会受到基因型的控制,与单性结实果实的发育关系不大。可见,有关赤霉素对茄果类蔬菜单性结实的诱导情况尚未研究清楚,其作用机制和规律尚不明确,今后可作为进一步研究的方向之一。
3.1.3 其他激素 大多数学者认为,IAA 和 GA3是调控园艺植物单性结实的主要内源激素,但仍有报道指出单性结实受 ZR、细胞分裂素、脱落酸等其他激素的影响[34]。张伟春等[29]研究发现,单性结实品系茄子的子房内 ZR 含量在开花后先急剧增加,在开花后 4 d 时有所降低,随后又发生回升。此外,该研究还发现,在授粉期单性结实品系的子房内 ZR含量比同期非单性结实品系的低 4.5 倍。因此认为,ZR 对茄子的单性结实性发挥重要的调控作用。同样的结论在张金凤[11]对辣椒的研究中也得以显示。但李艳玮等[28]研究发现,茄子子房内 ZR 激素水平与其授粉方式有关,人工授粉会显著提高单性结实品系茄子的 ZR 激素水平,而同等处理下单性结实品系和非单性结实品系的 ZR 含量变化趋于一致,因此认为,茄子单性结实果实的生长发育与ZR 无直接关系。武彦荣等[27]也同样认为,ZR 水平对诱导茄子单性结实的作用不明显,单性结实性主要受基因的控制。另外,Yoshihito 等[35]的研究发现,使用对乙烯不敏感的 Sletr1-1 突变体或 1-甲基环丙烯(1-MCP)阻断去雄花中的乙烯感知,增加了单性结实果实的长度,而同时用 GA 生物合成抑制剂多效唑(PAC)处理则会抑制单性结实;而乙烯前体1-氨基环丙烷-1-羧酸(ACC)应用于授粉子房会减少坐果,但 Sletr1-1 突变体单性结实果实并没有表现出生长素积累的增加,而是具有生物活性的 GAs水平升高。这表明乙烯可通过抑制 GA 代谢来促进番茄单性结实。Laura 等[36]的研究发现,番茄的单性结实还受脱落酸、细胞分裂素等激素的调控。王孝萱等[12]发现细胞激动素对番茄单性结实果实的形成起到调控作用。在对茄子单性结实激素诱导的研究中,吕忠恕等[37]发现,内源激素的平衡状态对诱导园艺植物单性结实有明显的作用。李艳玮等[28]的试验结果发现,高水平的 ABA 有利于诱导茄子单性结实,推测 ABA 在诱导茄子单性结实中作为一种促进激素来发挥作用。张金凤[11]在对辣椒的研究中也发现,单性结实品系辣椒的子房脱落酸含量明显高于非单性结实品系。另外,毛自朝等[38]的研究还发现,将 ipt 基因(异戊烯基转移酶基因)通过农杆菌转化到番茄基因组中,可使果实内细胞分裂素水平上升,诱导出单性结实的果实,可以推测细胞分裂素对番茄的单性结实诱导也起到一定作用。此 外,马凯等[39]在文章中提到黄瓜单性结实果实的形成受外源植物激素的诱导,胡仲远等[40]的研究还发现,使用软 X 射线辐射西瓜花粉可以提高果实内源激素水平,形成单性结实的高品质无籽果实。虽然有关诱导单性结实激素的研究较多,但关于各激素间比例对诱导单性结实的影响及同种激素对不同植物单性结实的诱导作用尚缺乏清晰的结论,可作为今后的研究方向之一。
3.2 环境
一些研究认为,茄果类蔬菜的单性结实性主要受温度的影响。刘富中[18]等经过研究得出,茄子的单性结实性在低温下更容易表现。李冰等[41]研究了单性结实品系茄子的最适生长温度,发现低温可以启动单性结实相关基因的表达,且最适温度为 7~15 ℃。张映等[42]研究表明,最低温度是诱导茄子单性结实的关键因素,可以启动相关基因的表达,并且相同的现象也出现在番茄和辣椒中。此外,孙春明等[43]发现,光周期是影响黄瓜单性结实的主要因素,认为短日照对单性结实起到一种类似生长素的效应。但光照对茄果类蔬菜的单性结实诱导作用尚未发现,李冰等[41]在对茄子单性结实率和日照时长的研究中得出二者无明显相关性。有关光周期对番茄和辣椒的单性结实性的影响目前还未有定论。
4 茄果类蔬菜单性结实的调控基因
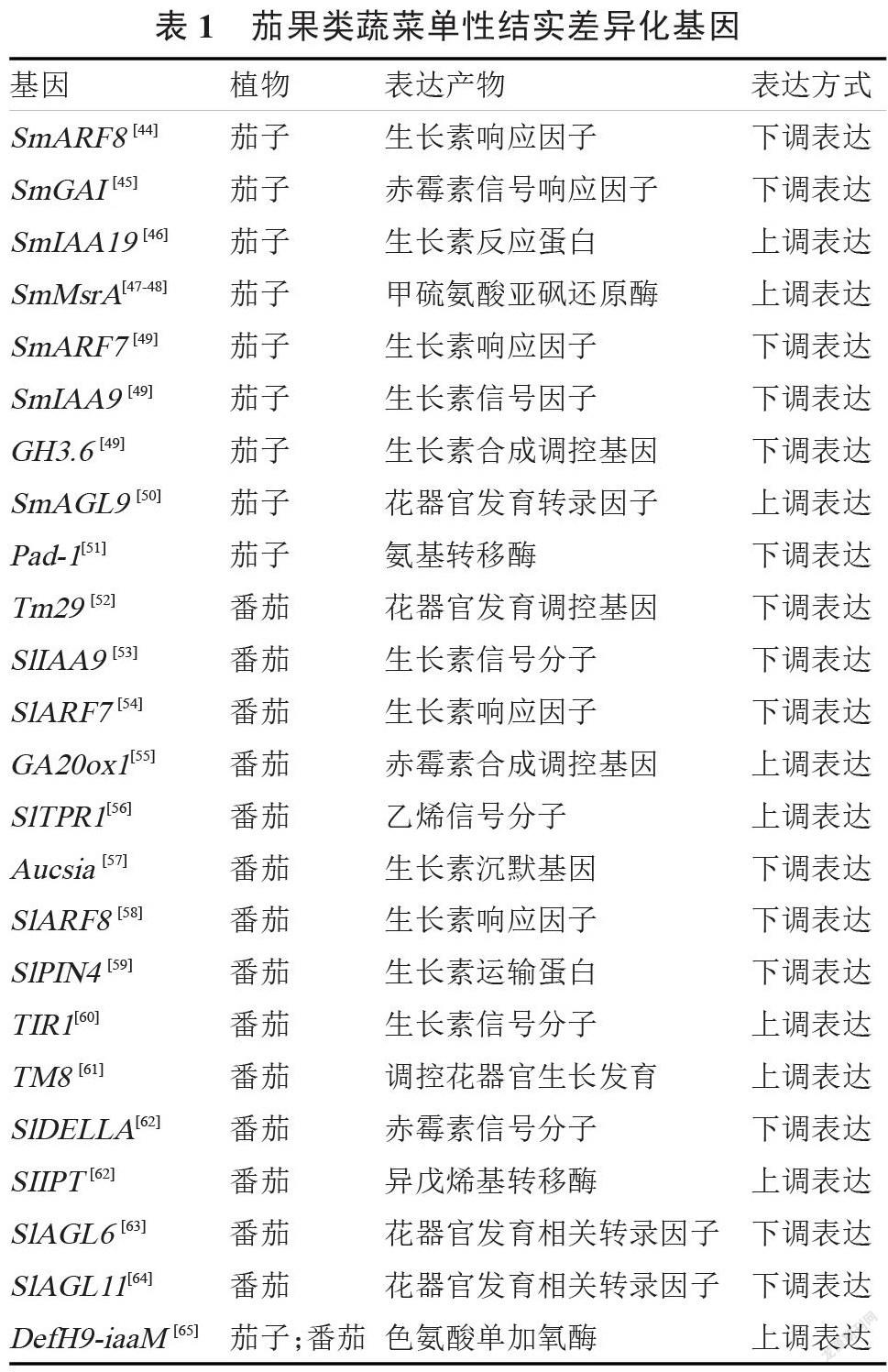
已有大量研究表明,园艺植物的单性结实性受内源激素和酶的调控,茄果类蔬菜单性结实相关基因的挖掘也多从激素调控基因和酶调控基因入手。笔者将目前已有报道的有关茄果类蔬菜单性结实的差异化基因进行了归纳整理(表 1),以供其他学者研究参考。
4.1 生长素诱导单性结实的分子机制
生长素是植物生长发育过程中的关键激素之一。研究发现,将 DefH9-iaaM 基因导入茄子、番 茄、烟草等植物中可获得单性结实果实,对其生长素含量的测定发现,DefH9-iaaM 可显著提高生长素在子房中的积累量,促使其子房在未受精的条件下膨大,产生单性结实果实[66]。DefH9-iaaM 基因编码色氨酸单加氧酶,可在生长素合成过程中促进吲哚乙酰胺的产生,大幅提高生长素在茄子子房中的合成量[67]。此外 ,Giuseppe 等[68]通过改良得到 De fH9-RI-iaaM 基因,在保证高单性结实性的同时,有效降低了茄子畸形果的产生。在生长素极性运输过程中,PIN(生长素输出载体)蛋白可通过其在膜上的不均匀分布调控生长素的运输方向[6],生长素运输蛋白基因(如番茄 SlPIN4)的表达可提高 PIN蛋白活性,该基因沉默将影响植物体内生长素的极性运输,导致生长素在子房积累,产生单性结实果实[59]。在生长素信号转导过程中,ARFs 和 AUX/IAA基因家族发挥着重要作用[69]。研究表明,生长素信号转导调控基因 ARFs[44]、AUX/IAA[53]在下调表达或沉默后可引起茄子单性结实。ARFs 基因可与AuxRE 发生特异性反应,二者在共同作用下可控制生长素响应分子相关基因的表达,从而调控生长素的信号转导过程。此外,F-box 基因家族的 TIR1 翻译为生长素受体蛋白,该基因过表达后可使黄瓜产生单性结实[60]。编码氨基转移酶的 Pad-1 基因通过催化吲哚-3-丙酮酸(IPyA)转化为色氨酸(Trp)来负向调控茄子的单性结实[51]。
4.2 赤霉素诱导单性结实的分子机制
赤霉素调控植物细胞的分裂分化,并参与果实的 形 成 过 程 。 有 研 究 发 现 ,赤 霉 素 的 合 成 受GA20oxl 基因的调控,该基因编码 GA20 氧化酶,是内源赤霉素合成的关键酶[6]。GA20oxl 基因过表达可提高赤霉素生物合成量,形成单性结实果实[70]。在赤霉素信号转导过程中,DELLA 蛋白会抑制赤霉素响应因子[6],从而抑制植物子房的发育,因此,DELLA 基因是单性结实的负调控因子。此外,GAI基因编码赤霉素信号响应因子来调控赤霉素信号转导过程。在茄子中,SmGAI 编码的氨基酸序列与番茄 SlDELLA 存在较高的同源性,因此推测其也有调控单性结实的功能[44]。
4.3 其他激素诱导单性结实的分子机制
ipt 基因编码异戊烯基转移酶,该酶可用于合成细胞分裂素前体 iPRPs(异戊烯基腺苷–5'–磷酸),进而用于细胞分裂素的内源合成[6]。有关 ipt 基因调控单性结实的报道多见于黄瓜的研究中。在对番茄的研究中,涂冬萍等[62]报道 SIIPT 基因参与细胞分裂素的生物合成,且正向调控单性结实性。此 外,Shinozaki 等[71]研究发现,番茄的单性结实性与乙烯有一定关系,TPR1 基因编码乙烯信号分子,其与乙烯受体 NR 和 LeETR1 发生相互作用,调节乙烯的体内合成运输过程[56]。
4.4 MADS-box 转录因子诱导单性结实的分子机制
MADS-box 基因家族为植物花器官发育过程中的关键转录因子,可协调花器官分生组织相关基因的表达来控制植物花器官发育和开花过程[72]。有关MADS-box 转录因子在单性结实的研究表明,控制雄蕊发育与形成的基因 TM29 在单性结实的番茄植株中下调表达,引起子房不育[52]。心皮发育调控基因 TM8 在单性结实番茄中上调表达,促进花器官和子房发育 ,形成单性结实果实[61]。 D 类和 E 类MADS-box 基因调控花器官分化[6],其中,AGL11 被发现有响应赤霉素信号的功能,在单性结实葡萄[73]和番茄[64]中下调表达。此外,AGL6 也被报道在梨[74]和番茄[63]的单性结实品系中下调表达,且发现该基因与赤霉素信号响应也有一定联系。
5 茄果类蔬菜单性结实的遗传机制
茄果类蔬菜单性结实的遗传机制尚未研究清楚,但诸多研究表明其属于多基因控制的数量性状。田时炳等[15]认为,茄子的单性结实属于隐形遗传,且非等位基因之间存在上位作用。刘富中等[75]用单性结实和非单性结实株系进行杂交,分析杂交后代单性结实性状分离的情况,结果发现,供试材料的单性结实是受细胞核内的单显性基因控制。另外,张映[42]等的研究也得到了同样的结论。薛 萍[76]的研究发现,茄子单性结实为隐性遗传,在遗传特征上属于加性-显性-上位性遗传模型。李冰等[41]报道,通过单性结实品系和非单性结实品系的杂交得到 F1代,超亲优势为负值。此外还发现,茄子的单性结实性状受基因的加性效应影响较大,达到极显著水平,广义遗传率和狭义遗传率均比较高。有关番茄和辣椒的研究也得到类似的结论。朱为民等[77]的试验得出,单性结实番茄材料 OS 和 SN 的单性结实是隐性性状,受一对隐性基因控制。张金凤[11]在对辣椒的研究中得出,辣椒的单性结实为负显性-加性-上位性遗传模型,属于隐性遗传。在其他植物的单性结实研究中,华南型旱黄瓜的单性结实性符合加性-显性-上位性遗传模型,属于由 1 对不完全显性基因控制的数量性状[78]。虞夏清等[79]对广西地方黄瓜等 11 个不同的黄瓜品种进行杂交,统计了不同组合的子代单性结实能力,结果发现,供试材料单性结实能力的广义遗传率和狭义遗传率均偏低,且变异系数高,这表明其单性结实性状通过杂交、改善种植技术等方法来获得變异材料的概率较大。由于供试材料不同,茄果类蔬菜的遗传机制不尽相同。可见单性结实是由多基因控制的,除了存在显性效应和加性效应的作用,还可能存在上位性等其他遗传效应的作用,遗传机制较为复杂,需要今后进一步研究来确定。
6 问题与展望
6.1 茄果类蔬菜单性结实的种质资源较少
近年来,随着设施栽培技术的不断完善,农业上对无籽果实的需求日益增加。虽然茄果类蔬菜有关单性结实的资源发现得较早,但仍缺乏更为广泛和深入的研究,在实际的农业生产中缺少可应用的单性结实品系。茄子和番茄在试验中已有不少可供研究的单性结实品系,但由于其机制尚未明确,产品性状不稳定,仍无法推广应用于实际生产中。有关辣椒的单性结实研究起步较晚,可用于试验的单性结实品系较少,后续的研究难以进行,目前辣椒多采用单倍体育种技术获得具有优良性状的试验材料[80]。另外,当前用于单性结实研究的资源大多属于环境敏感型,其单性结实性易受温度等环境因素的影响,在研究过程中难以精确地判断其发生条件。因此,相关研究应对已获得的单性结实资源进行改良,研究其遗传规律和分子机制,为单性结实品系的发掘和选育奠定理论基础。
6.2 茄果类蔬菜单性结实的诱导因素尚待研究
诸多研究发现,植物体内的关键性内源激素(生长素、赤霉素等)都有调控茄果类蔬菜单性结实性的作用。但不同激素的调控水平和调控方式各不相同,作用机制复杂多样,当前研究仍不是很深入,具体的调控过程仍不明确。各激素的具体作用、不同激素之间比例和平衡性对单性结实性的诱导也有待进一步的研究。此外,当前各学者对影响茄果类蔬菜单性结实的环境因素的研究集中在温度上,缺乏对光强、光质、光周期等其他环境因子诱导作用的研究,且关于土壤中的无机元素对单性结实性的诱导作用也鲜有报道。明确影响单性结实的因素,可以更好地进行遗传育种和分子育种试验,在今后的应用和推广中充分发挥单性结实品系的潜能。
6.3 茄果类蔬菜单性结实的遗传方式尚不明确
截至目前,茄果类蔬菜单性结实相关基因的研究主要为内源激素和酶的调控基因,相关基因的挖掘已有不少的报道,但仍未得到与单性结实性状距离较近的分子标记。进一步开发分子标记将有利于茄果类蔬菜单性结实的鉴定评价和品种选育。此外,茄果类蔬菜单性结实的分子机制研究起步较晚,关于遗传机制的研究较少,现有的研究结论并不统一,遗传方式尚未有定论。因此,挖掘茄果类蔬菜单性结实相关的关键基因,分析表达模式,探究单性结实具体的遗传机制,并利用其遗传特性和分子标记选育新品种是下一步研究的重点方向。
参考文献
[1]刘正鲁.茄果类蔬菜植物工厂[J].生命世界,2019(10):41.
[2]李香景,刘富中,张映,等.茄子单性结实研究进展[J].中国蔬菜,2012(6):8-14.
[3]刘继辉,邹春华,徐勋志,等.茄果类蔬菜轻简化栽培技术[J].辣椒杂志,2019,17(3):26-31.
[4]南思楠,邹薛,闫道良.植物单性结实的发生诱因及其机制研究[J] .安徽林业科技,2021,47(5):11-15.
[5]陈学好,陶俊,曹碚生.园艺作物单性结实的类型[J].生物学通报,2001,35(9):6-7.
[6]刘剑峰,朱娅玫,陈学好,等.园艺作物单性结实功能基因研究进展[J].分子植物育种,2020,18(18):6007-6015.
[7]VASILIOS R,KONSTANTINOS S,GEORGIOS T,et al.Seeded and parthenocarpic cherry tomato fruits exhibit similarsucrose,glucose,and fructose levels,despite dissimilarities in UGPase and SPS gene expression and enzyme activity[J].Journal of Plant Growth Regulation,2015,34(1):47-56.
[8]潘秀清,高秀瑞,武彦荣,等.茄子单性结实果实发育规律与营养物质的关系[J].华北农学报,2007,22(2):50-52.
[9]吕玲玲,李威,肖熙鸥.具有单性结实能力茄子的耐冷性研究[C]//中国作物学会.2015 年学术年会论文摘要集.黑龙江哈尔滨,2015:128.
[10] 刘绍银.单性结实机理概述[J].生物学教学,2016,41(6):72-73.
[11] 张金凤.辣椒单性结实习性、生理特性及遗传规律的研究[D].江苏扬州:扬州大学,2009.
[12] 王孝宣,杜永臣,李树德,等.番茄中的单性结实[J].中国蔬菜,1999(5):44-48.
[13] 张祥胜,陈日远,刘厚诚.蔬菜单性结实研究进展[C]//中国园艺学会第五届青年学术讨论会论文集.广州:广州出版社,2002:396-404.
[14] 张少丽,邵景成,胡志峰,等.番茄单性结实研究进展[J].园艺学报,2016,43(1):183-192.
[15] 田时炳,刘富中,王永清,等.茄子单性结实性的遗传分析[J].园艺学报,2003,30(4):413-416.
[16] RESTAINO F,PERRONE D F,CORREALE A.New parthenocarpic geno-types of eggplant suitable for greenhouse cultivation[C]//PALLOIX A,DANNAYMC.Xth meeting on genetics and breeding of capsicum and eggplant.Paris:INRAParis,1998:273.
[17] 成玉富,薛萍,張金凤.茄子单性结实习性的研究[C]//中国茄子大会暨学术研讨会论文集.武汉:《中国蔬菜》编辑部,2008:29-31.
[18] 刘富中,连勇,陈钰辉,等.温度和蕾期去雄及去柱头处理对茄子单性结实性的影响[J].园艺学报,2005,32(6):1021-1025.
[19] 潘秀清,武彦荣,高秀瑞.茄子单性结实材料 D-11 的发现[J].华北农学报,2005,20(6):33.
[20] 李艳玮,刘富中,陈钰辉,等.温度对茄子单性结实子房(果实)发育过程中内源激素含量的影响[J].中国蔬菜,2013(22):32-38.
[21] ICHIRO H,HIROSHI M,KAORI K,et al.Identification of pepper(Capsicum annuum L.)accessions with large or small fruit that have a high degree of parthenocarpy[J].Scientia Horticulturae,2012,135:68-70.
[22] 王静,张伟春,魏毓棠,等.茄子单性结实的果实内可溶性糖、蛋白质含量变化的研究[J].辽宁农业科学,2005(1):38-39.
[23] 陈学好,曾广文,曹碚生.园艺作物的单性结实及应用[J].植物生理学通讯,2001,37(6):570-575.
[24] MAO Z C,YU Q J,ZHEN W,et al.Expression of ipt genedriven by tomato fruit specific promoter and its effects on fruitdevelopment of tomato[J].Chinese Science Bulletin,2002,47(11):928-933.
[25] 王利英,石瑶,刘文明,等.番茄单性结实遗传效应分析和新材料的选育[J].天津农业科学,2008,14(2):38-40.
[26]TAKASHI I,HIROSHI Y,MASAYUKI O A,et al.Growthdependence of ovaries of facultatively parthenocarpic eggplant in vitro on indole- 3- acetic acid content[J].Scientia Horticulturae,1999,79(3/4):143-150.
[27] 武彥荣,郭秀林,高秀瑞,等.茄子单性结实系开花期内源激素含量的变化[J].河北农业科学,2009,13(9):14-16.
[28] 李艳玮,张映,杨锦坤,等.茄子单性结实发育过程中内源激素含量的变化[J].北方园艺,2019(10):9-16.
[29] 张伟春,魏毓棠,王静,等.茄子单性结实与非单性结实品系胚胎发育及果实解剖结构的观察[J].沈阳农业大学学报,2008,39(5):534-537.
[30] MAPELLI S,FROVA C,TORTI G,et al.Relationship betweenset,development and activities of growth regulators in tomatofruits[J].Plant and Cell Physiology,1978,19(7):1281-1288.
[31] HAZRA P,K D A,CHATTERJEE P.Altered gibberellin and auxin levels in the ovaries in the manifestation of genetic parthenocarpy in tomato(Solanum lycopersicum)[J].Current Science,2010,99(10):1439-1443.
[32] FOS M,NUEZ F,GARCı́A-MARTı́NEZ J L.The Gene pat-2,which induces natural parthenocarpy,alters the gibberellin content in unpollinated tomato ovaries[J].Plant Physiology,2000,122(2):471-479.
[33] FRANCESCO M,MARÍA L V,LORENZO M,et al.Induction of gibberellin 20-oxidases and repression of gibberellin 2β-oxidases in unfertilized ovaries of entire tomato mutant,leads to accumulation of active gibberellins and parthenocarpic fruit formation[J].Plant Growth Regulation,2015,75(2):415-425.
[34] DING J G,CHEN B W,XIA X J,et al.Cytokinin-induced parthenocarpic fruit development in tomato is partly dependent on enhanced gibberellin and auxin biosynthesis[J].Plos One,2013,8(7):e70080.
[35] YOSHIHITO S ,SHUHEI H ,MIKIKO K ,et al .Ethylene suppresses tomato( Solanum lycopersicum )fruit set through modification of gibberellin metabolism[J].The Plant Journal,2015,83(2):237-251.
[36] LAURA P,JOSE M B,JOAQUIN C,et al.Transcriptomic analysis of tomato carpel development reveals alterations in ethylene and gibberellin synthesis during pat3/pat4 parthenocarpic fruit set[J].BMC Plant Biology,2009,9(1):67-84.
[37] 呂忠恕,王保民,张承烈,等.开花前后子房中生长调节物质的变化及其与结果及单性结果的关系[J].植物生理学报,1979,5(3):253-260.
[38] 毛自朝,于秋菊,甄伟,等.果实专一性启动子驱动 ipt 基因在番茄中的表达及其对番茄果实发育的影响[J].科学通报,2002,47(6):444-448.
[39] 马凯,牛莉莉,唐艳领,等.黄瓜果实发育相关性状的分子调控研究进展[J].中国瓜菜,2019,32(10):1-5.
[40] 胡仲远,张明方,蓝善荣,等.软 X 射线辐照花粉通过影响西瓜中激素信号诱导单性结实[J].中国瓜菜 ,2019,32(8):253-254.
[41] 李冰,武彦荣,高秀瑞,等.茄子单性结实性遗传效应研究[J].河北农业科学,2010,14(12):50-51.
[42] 张映,陈钰辉,连勇,等.园艺植物单性结实的分子研究进展[J].园艺学报,2018,45(7):1402-1414.
[43] 孙春明.黄瓜单性结实研究概述[J].上海农业科技,2004(3):69-70.
[44] 杜黎明,毛伟海,包崇来,等.茄子生长素响应因子 SmARF8 的克隆与分析[J].中国农业科学,2009,42(7):2434-2441.
[45] 杜黎明,包崇来,胡天华,等.茄子 GA 响应因子 SmGAI 的克隆与分析[J].中国蔬菜,2009(16):26-30.
[46] 张伟伟,刘富中,张映,等.茄子生长素诱导基因 SmIAA19 的克隆和分析[J].园艺学报,2014,41(11):2231-2240.
[47] 赵祯.茄子单性结实差异表达序列鉴定及 SmMsrA 基因功能初步分析[D].北京:中国农业科学院,2015.
[48] 郭亚鹤,赵祯,张映,等.茄子单性结实差异表达序列鉴定及分析[J].园艺学报,2017,44(10):1914-1924.
[49] 张立慧.茄子单性结实相关基因的克隆及表达分析[D].重庆:西南大学,2016.
[50] 陈露露.茄子 SmMsrA 基因启动子互作蛋白的鉴定及单性结实转录组研究[D].北京:中国农业科学院,2021.
[51] MATSUO S,MIYATAKE K,ENDO M,et al.Loss of function of the Pad-1 aminotransferase gene,which is involved in auxin homeostasis,induces parthenocarpy in Solanaceae plants[J].Proceedings of the National Academy of Sciences of the United States of America,2020,117(23):12784-12790.
[52] AMPOMAH- DWAMENA C,MORRIS B A,SUTHERLAND P,et al.Down-regulation of TM29,a tomato SEPALLATA homolog,causes parthenocarpic fruit development and floral reversion[J].Plant Physiology,2002,130(2):605-617.
[53] WANG H,JONES B,LI Z G,et al.The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis[J].The Plant cell,2005,17(10):2676-2692.
[54] GOETZ M,HOOPER L C,JOHNSON S D,et al.Expression of aberrant forms of AUXIN RESPONSE FACTOR8 stimulates parthenocarpy in arabidopsis and tomato[J].Plant Physiology,2007,145(2):351-366.
[55] IRENE O,FRANCESCA S,RICCARDO C,et al.Tomato fruit set driven by pollination or by the parthenocarpic fruit allele are mediated by transcriptionally regulated gibberellin biosynthesis[J].Planta,2007,226(4):877-888.
[56] LIN Z F,ARCIGA-REYES L,ZHONG S L,et al.SlTPR1,atomato tetratricopeptide repeat protein,interacts with the ethylene receptors NR and LeETR1,modulating ethylene and auxinresponses and development[J].Journal of Experimental Botany,2008,59(15):4271-4287.
[57] MOLESINI B,PANDOLFINI T,ROTINO G L,et al.Aucsiagene silencing causes parthenocarpic fruit development in tomato[J].Plant Physiology,2009,149(1):534-548.
[58] 王飞燕,吴健,张佳景,等.番茄生长素应答因子 SlARF8-1 的分离与 SlARFs 表达特征分析[J].浙江大学学报(农业与生命科学版),2011,37(3):237-244.
[59] MOUNET F,MOING A,KOWALCZYK M,et al.Down-regulation of a single auxin efflux transport protein in tomato induces precocious fruit development[J].Journal of Experimental Botany,2012,63(13):4901-4917.
[60] 崔利.基于转录组学的黄瓜单性结实分子调控机制研究[D].南京:南京农业大学,2013.
[61] MARGHERITA D,SIMONA M,FRANCESCA R,et al.Characterization of TM8,a MADS- box gene expressed in tomato flowers[J].BMC Plant Biology,2014,14(1):319-333.
[62] 涂冬萍.罗汉果刺激性单性结实果实的生物学特性研究[D].北京:北京协和医学院,2015.
[63] CHEN K,ESTER Y,ANTHONY M B,et al.Tomato facultative parthenocarpy results from SlAGAMOUS- LIKE 6 loss of function[J].Plant Biotechnology Journal,2017,15(5):634-647.
[64] HUANG B,ROUTABOUL J,LIU M,et al.Overexpression of the class D MADS- box gene Sl-AGL11 impacts fleshy tissue differentiation and structure in tomato fruits[J] .Journal of Experimental Botany,2017,68(17):4869-4884.
[65] GIUSEPPE L R,NAZARENO A,EMIDIO S,et al.Open fieldtrial of genetically modified parthenocarpic tomato:seedlessness and fruit quality[J].BMC Biotechnology,2005,5(1):32-39.
[66] PANDOLFINI T,ROTINO G L,CAMERINI S,et al.Optimisation of transgene action at the post- transcriptional level:high quality parthenocarpic fruits in industrial tomatoes[J].BMCBiotechnology,2002,2(1):1-11.
[67] 涂冬萍,马小军,莫长明,等.单性结实的分子研究进展[J].中草药,2014,45(20):3034-3040.
[68] GIUSEPPE L R,DARASINH S,FULVIA R,et al.Current status in production and utilization of dihaploids from somatic hybrids between eggplant( Solanum melongena L.)and its wild rela-tives[J].Acta Physiologiae Plantarum,2005,27(4):723-733.
[69] 张佳景.番茄 LeARF8 基因克隆及其时空表达特性的研究[D].杭州:浙江大学,2008.
[70] GARCIA-HURTADO N,CARRERA E,RUIZ-RIVERO O,et al.The characterization of transgenic tomato overexpressing gib berellin 20- oxidase reveals induction of parthenocarpic fruit growth,higher yield,and alteration of the gibberellin biosynthet ic pathway[J].Journal of Experimental Botany,2012,63(16):5803-5813.
[71] SHINOZAKI Y,HAO S H,KOJIMA M,et al.Ethylene suppresses tomato(Solanum lycopersicum)fruit set through modification of gibberellin metabolism[J].Plant Journal,2015,83(2):237-251.
[72] 王瑩,穆艳霞,王锦.MADS-box 基因家族调控植物花器官发育研究进展[J].浙江农业学报,2021,33(6):1149-1158.
[73] 崔梦杰.葡萄 miR156s 及其靶基因 VvSBPs 在果实发育与成熟中的作用分析[D].南京:南京农业大学,2018.
[74] 刘璐璐.GA4+7诱导砀山酥梨单性结实的机制研究[D].陕西杨凌:西北农林科技大学,2017.
[75] 刘富中,周亚君,张映,等.茄子单性结实特性及相关基因的分离[C]//《园艺学报》编辑部:中国园艺学会 2010 年学术年会论文摘要集(辽宁沈阳),2010:107.
[76] 薛萍.茄子单性结实习性、生理机制及遗传规律的研究[D].江苏扬州:扬州大学,2006.
[77] 朱为民,朱龙英,薛林宝.番茄单性结实性的遗传效应和转育[J].上海农业学报,2003,19(1):23-26.
[78] 牛志红,李晓丽,宋晓飞,等.旱黄瓜单性结实性遗传分析[J].河北科技师范学院学报,2019,33(4):24-28.
[79] 虞夏清,任晓政,陈劲枫.新型光皮长黄瓜重要性状杂种优势和配合力分析[J].中国瓜菜,2020,33(12):22-28.
[80] 姚秋菊,常晓轲,程志芳,等.辣椒单倍体育种技术研究进展[J].中国瓜菜,2021,34(7):1-6.
