广东南岭植物群落物种多样性沿海拔梯度分布格局*
2022-04-25李泽华
徐 卫 杨 婷 李泽华 周 平
(广东省科学院 广州地理研究所,广东 广州510075)
山地具有复杂多变的生境条件,多种生物在此共存、繁衍,因此山地是生物多样性保护和自然资源可持续利用的重要场所[1-2]。在山地生态系统中,海拔综合了温度、水分等诸多环境因素的影响,有研究表明温度随海拔梯度变化要比随纬度梯度变化快1 000 倍,是影响生物多样性和群落结构的重要因素[3-4]。因此,山地植物群落研究中,生物多样性沿海拔梯度的变化规律一直是生态学家研究的重要内容,但并未达成普遍共识[5-6]。前人的研究中物种多样性与海拔梯度的相关关系归纳为4 种,即单调上升、单调下降、单峰曲线、无规律和先上升后平台,其中最常见的研究结果为单峰曲线和单调下降[6]。Lomolino[7]认为,多样性与海拔梯度格局的关系(正相关、负相关或单峰分布格局)在很大程度上依赖于环境变量之间的协变与互作。Rahbek[8]认为以往相关研究可能没有覆盖到完整海拔梯度,会造成物种多样性沿海拔梯度单调递增或递减的假象。因此在山地植被群落生物多样性分布格局的研究中,阐明生物多样性与海拔的相关性是重要前提。
国内很多关于山地植物群落物种多样性的垂直分布格局的研究,但因研究区域不同、山体坡向不同,得到的结果也各不相同[3-6,9-20]。其中,温带地区长白山北坡植物群落α 多样性随海拔增加呈线性下降趋势[19,21]。暖温带地区的太白山北坡和祁连山北坡植物群落生物多样性随海拔上升呈现先升后降的单峰变化曲线[18,22];福建武夷山黄岗山东南坡常绿阔叶林内木本植物多样性指数随海拔升高逐渐降低,西北坡的常绿阔叶林的多样性指数在海拔梯度上呈现先减小后增大的“V”型变化[5,9]。湖北神农架及云南紫溪山南坡等木本植物多样性的垂直格局为单峰曲线[15-16,23],鼎湖山植物群落及四川龙门山和卧龙保护区乔木多样性指数随海拔升高而呈下降趋势[13-14,24]。同时有研究表明,不同生活型植物多样性具有不同的海拔格局。赵淑清等[25]在长白山北坡发现木本植物多样性随海拔升高呈下降趋势,草本植物变化趋势不明显。喜马拉雅山地区的木本植物随海拔升高呈单峰格局,草本植物多样性变化不明显[26]。
国内亚热带山地植被研究多集中在西南地区[9,11-12,14,24],南部山地尤其是南岭的研究相对较少。南岭山地位于中亚热带,地质上属华南褶皱带,中国植被区划上属于中亚热带常绿阔叶林南部亚地带,作为国际意义上的陆地生物多样性关键地区之一,拥有同纬度带上保存最完整的亚热带植被[27-29]。南岭山地对于植被科学的研究已有了一定的积累,多集中在植被分布和植物区系方面,广东省北部南岭山地内群落结构、树种多样性的分布格局研究相对较少[20,30-32]。本研究依据广东南岭山地沿海拔梯度植被样方调查数据,分析南岭山地植物群落结构和物种多样性的垂直分布格局,对比分析不同海拔梯度植物群落的多样性和物种组成变化,为未来南岭地区生物多样性保护和自然保护区建设提供科学的参考依据。
1 研究方法
1.1 研究区概况
研究区处于广东省北部南岭国家自然保护区内,位于广东省北部南岭山脉中段,地跨韶关、清远两市的乳源、阳山和连州三县(市),由乳阳片区、称架片区、大东山片区和大顶山管 理 片 区 组 成。地 理 坐 标 为24°37′~24°57′N,112°30′~113°04′E,总面积达5.84 万hm2。最高峰石坑崆海拔1 902 m,相对高差1 489 m,亦是广东省的最高峰“广东第一峰”的主峰。南岭国家自然保护区属中亚与南亚热带过渡的地带湿润气候,多年平均气温17.7 ℃,极端最低气温-4.2 ℃,极端最高气温34.4 ℃;≥10℃年积温6 483 ℃。多年平均降水量为1 705 mm,降水多集中3—10月间,占全年降雨量的82%左右;多年平均相对湿度84%。年平均无霜期276 天,平均霜期89天。历年平均日照时数为1 234 h,年平均日照百分率为40%。成土母质为花岗岩,土壤类型为山地黄壤和黑钙土。
1.2 研究方法
1.2.1 样地设置 本研究依托于南岭森林生态系统国家野外科学观测研究站的研究样地,海拔跨度从413 m 到1 698 m,植被类型随海拔升高变化趋势为“沟谷常绿阔叶林-山地常绿阔叶林—针阔叶混交林—山地矮林—高山草甸”,共24 个森林样地(图1)。其中每个样点包括1 个乔木层样方,面积为40 m× 40 m,由16 个10 m×10 m 的小样方组成。每个乔木层样方内设置3 个灌木样方,面积为2 m× 2 m,3 个草本植物样方,面积为1 m× 1 m。高山草甸内设置草本样方3 个,面积为20 m× 20 m。
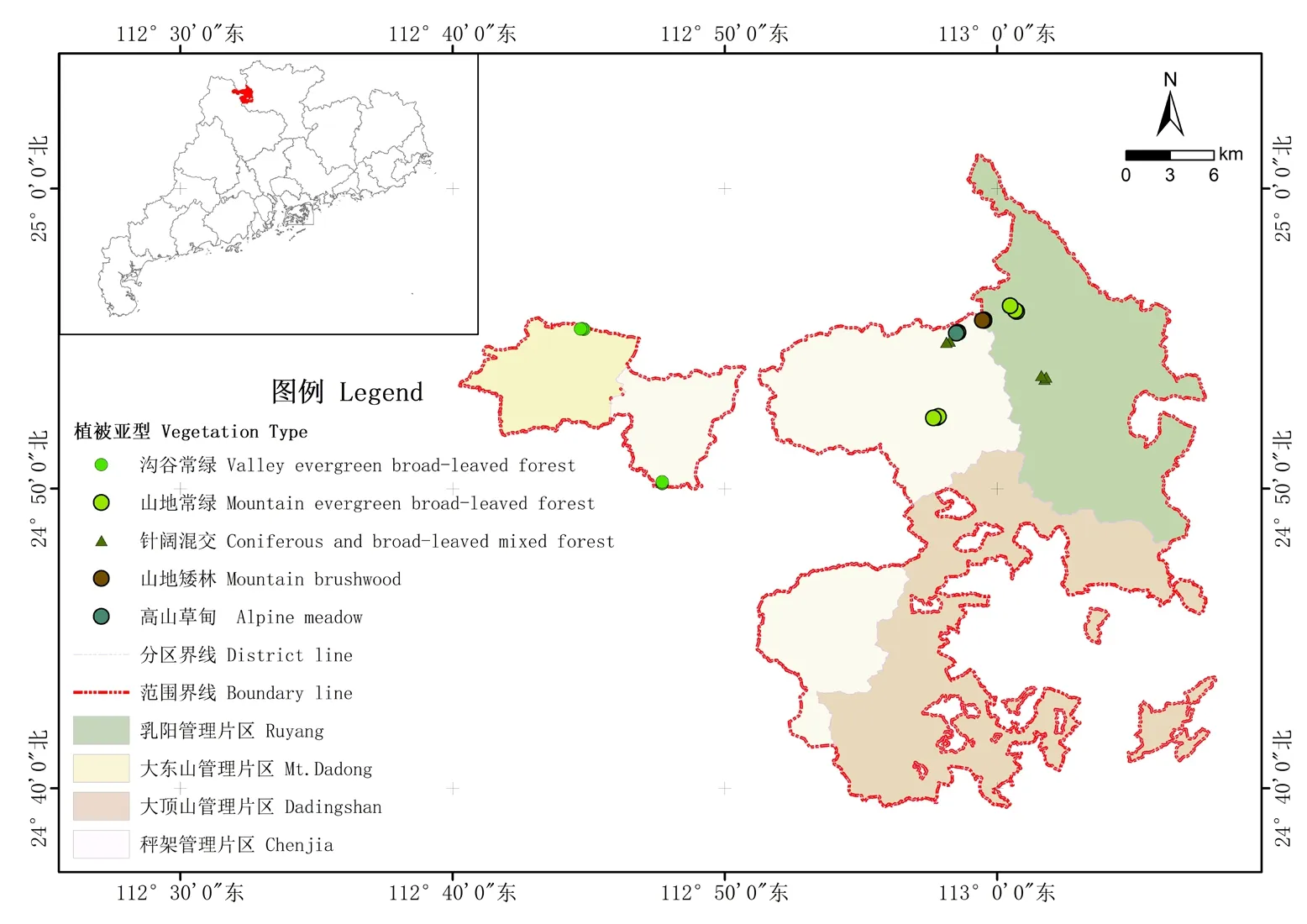
图1 南岭调查样点分布Figure 1 Distribution of sample plots in Nanling Mountains
记载样地的植被类型、经纬度、海拔、方位角、坡度、坡向。2018 年对样方中所有胸径(DBH)≥ 1 cm 的木本植物个体进行定位、鉴定和挂牌标记,并测量其胸径、树高、冠幅;调查灌木的物种名、基径、株高和丛幅;调查草本植物的物种名称、群落盖度、株数、高度等。
1.2.2 α 生物多样性指数计算 选取物种丰富度、Shannon-Wiener 指数、Simpson 指数和Pielou 均匀度指数,计算方式如下:
物种丰富度:S=样方内出现的物种数。


其中,Pi为第i个物种占总数的比例[31]。
1.2.3 气候指标估算 从南岭地区11 个气象监测站点获取的2003—2017 年日尺度气象资料,以年平均降水量(Mean annual precipitation, MAP)和年平均气温(Mean annual temperature, MAT)作为水分和热量指标,通过建立气象因子与气象站点经纬度和海拔的关系,估算各样地的气象指标[32]。
2 结果与分析
2.1 物种组成
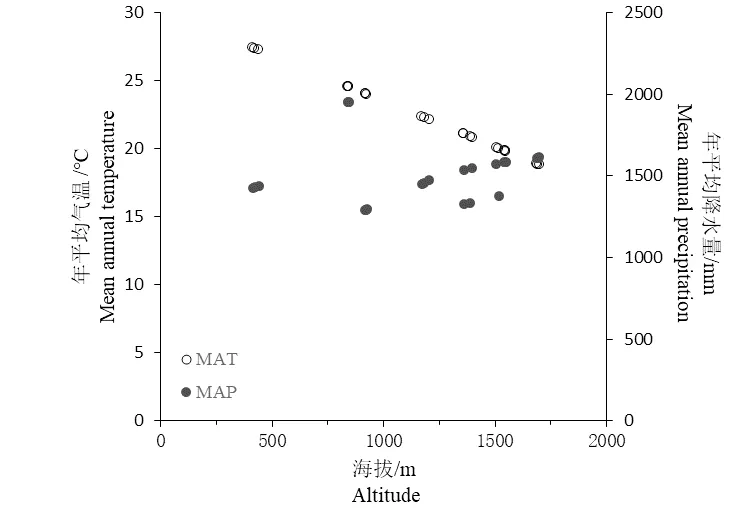
图2 南岭山地气象因子与海拔的关系Figure 2 Relationship between altitude and meteorological factors in Nanling Mountains
南岭山地属中亚热带常绿阔叶林南部亚地带,地带性植被为常绿阔叶林,随海拔增加的植被类型变化依次为常绿阔叶林、针阔混交林、山地矮林和高山草地。作为南岭山地的地带性植被,研究区内的常绿阔叶林分为河谷常绿阔叶林和山地常绿阔叶林。沟谷常绿阔叶林的物种丰富度(58 ±12.48)略低于山地常绿阔叶林(61.67 ± 17.39),而Shannon-Wiener 指数(3.31 ± 0.42)略高于山地常绿阔叶林(3.29 ± 0.39)。沟谷常绿阔叶林主要分布在海拔900 m 以下的沟谷内,以广东润楠Machilus kwangtungensis、华润楠M. chinensis等樟科润楠属乔木和壳斗科的鹿角锥Castanopsis lamontii占优势。而山地常绿阔叶林主要分布在海拔900 ~1 360 m 的山体,主要以甜槠Castanopsis eyrei、水青冈Fagus longipetiolata、鹿角锥、罗浮锥C. faberi、米槠C.carlesii等壳斗科树种为优势树种。针阔混交林和山地矮林的物种丰富度和多样性指数均低于常绿阔叶林,其中针阔混交林内针叶树种主要以广东松Pinus kwangtungensis和马尾松P. massoniana为主,阔叶树种以荷木类Schimaspp、青冈Cyclobalanopsis glauca和甜槠等为主。山地矮林主要分布在海拔1 600 m以上的山地,是常绿落叶阔叶混交林适应南岭山顶风大的特殊环境条件的一种植被变型,平均树高在6 m 以下,优势树种主要有野茉莉Styrax japonicus、青冈和冬青Ilex chinensis等(表1)。
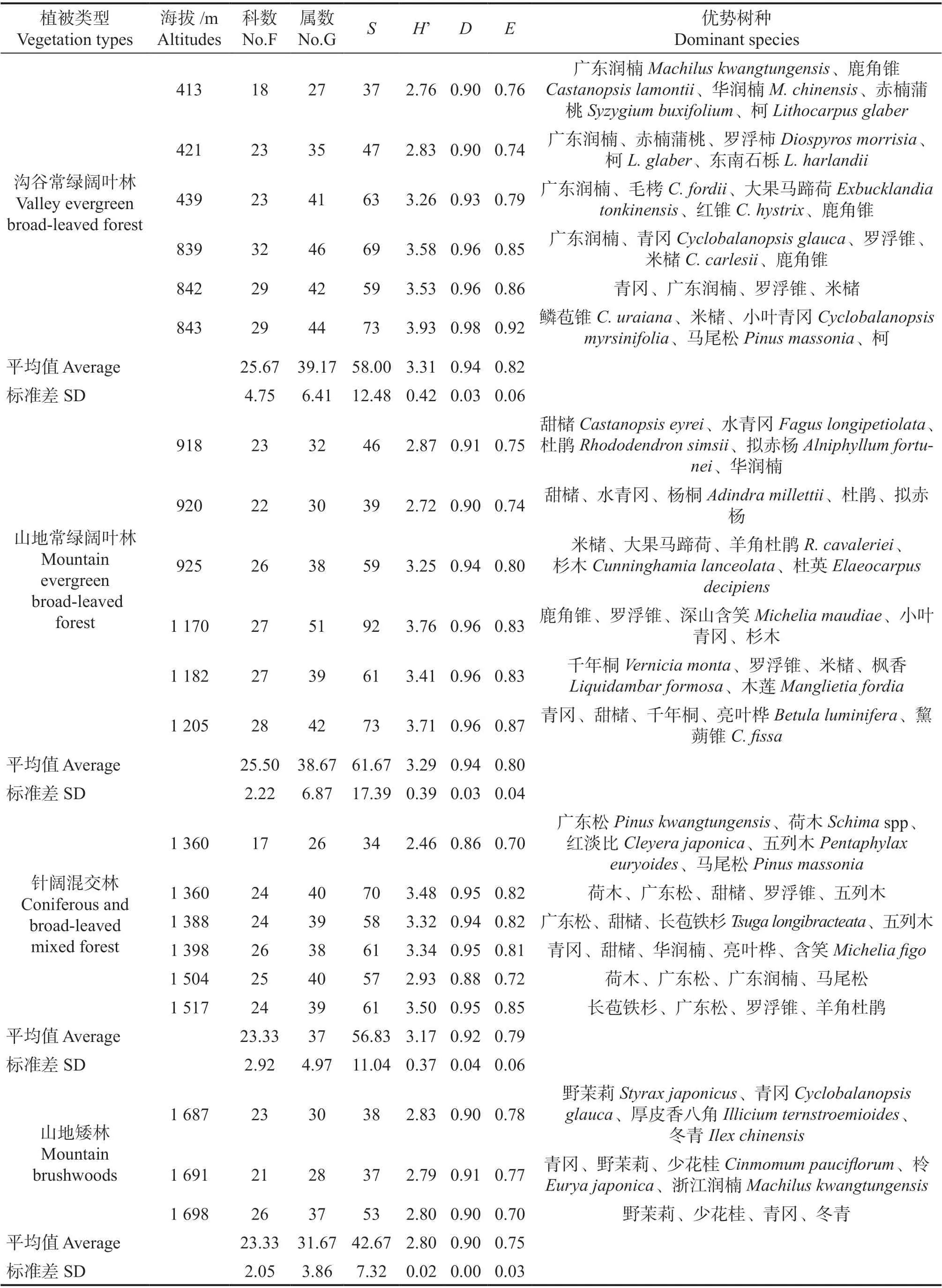
表1 南岭山地沿海拔梯度乔木物种组成变化Table 1 Comparison of tree species composition along altitude on Nanling Mountains
2.2 群落结构
随着海拔的增加,乔木层的最大树高(Hmax)呈现单峰变化趋势。乔木的最大树高从低海拔常绿阔叶林的20.83 m 增加到针阔混交林内的26 m,后减少到亚热带山顶常绿阔叶矮林的10.17 m 左右。其中小黄山内海拔1 388 m 样方内最大树高26 m。最大树高的变化反映了随着海拔的变化,树木生长潜力发生改变。与最大树高的变化相似,乔木层的最大胸径(DBHmax)随海拔增加的变化曲线单峰分布格局,表现出先增加后减少的趋势。从低海拔(413 m)的沟谷常绿阔叶林内乔木的最大胸径39.3~51.5 cm 增加到中海拔(1 170 m)的山地常绿阔叶林内乔木最大胸径72 cm,中高海拔的针阔混交林内乔木最大胸径降低到58.4 cm,海拔1 600 m 以上的山地常绿矮林内乔木最大胸径降低到42 cm。乔木物种的胸高断面积和(BA)随海拔梯度增加的变化趋势与乔木最大胸径相似,即随着海拔增加,胸高断面积和为单峰曲线变化格局。其中位于海拔1 170 m 的八宝山处的山地常绿阔叶林内乔木层的胸高断面积和为最大值。立木密度(N)随海拔上升表现出不明显的下降趋势。群落内立木密度对海拔的变化响应并不明显,不同植被类型森林群落的立木密度有所差别,海拔1 170 m 的八宝山处的山地常绿阔叶林样方内乔木层立木密度值最高,但针阔混交林内乔木层平均立木密度高于其他3 种植被类型(图3)。
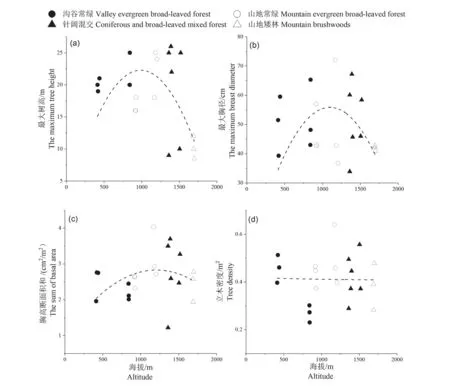
图3 南岭山地乔木层的最大树高(a)、最大胸径(b)、胸高断面积和(c)和立木密度(d)随海拔梯度的变化Figure 3 Changes of maximum tree height (Hmax), maximum diameter at breast height (DBHmax), total basal area (BA) and tree density (N) along the altitude gradient on Nanling Mountains
南岭山地中随着海拔的增加,森林群落内乔木个体的生活型组成也发生变化(图4)。常绿阔叶树种在研究区域的群落内均占据优势地位,这符合南岭地带性植被研究的结论。常绿阔叶树种的比例随着海拔的增加呈现降低的趋势,这与落叶阔叶树种和针叶树种的比例增加有关。落叶阔叶植物占比随着海拔缓慢增加,主要为安息香属、槭属和柿属的一些物种。在南岭山地调查区域内,针叶树种的分布主要在海拔1 300 m 以上,海拔1 000 m 以下分布的针叶树种主要是马尾松和杉木,分布在大东山海拔800~900 m 的沟谷常绿阔叶林内。在海拔1 600 m 以上的山地常绿矮林中分布的针叶树为白豆杉Pseudotaxus chienii,均为DBH 小于10 cm 的个体。分布在1 000 ~1 500 m中海拔的山地的针叶树种多为广东松和马尾松,与常绿阔叶树种形成比较明显的混交状态。
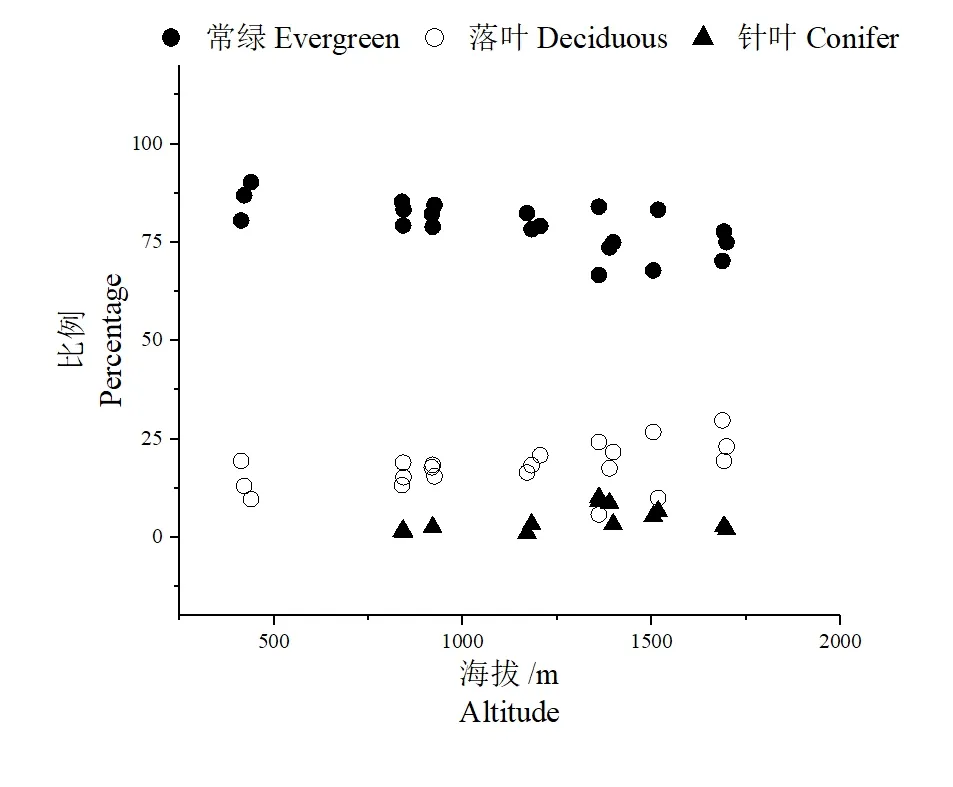
图4 南岭山地乔木生活型组成比例随海拔的变化Figure 4 Changes in the proportion of tree life forms along altitude of Nanling Mountains
2.3 物种多样性
乔木物种数随海拔变化趋势呈单峰曲线分布格局,灌木和草本的物种数海拔格局为线性单调递减。乔木树种的Shannon-Wiener 多样性指数随海拔升高呈现单峰曲线变化模式,而Simpson 多样性指数随海拔升高的变化为不明显的下降趋势。灌木和草本植物的Shannon-Wiener 多样性指数和Simpson 多样性指数的海拔分布格局为下降趋势。但由于山地矮林的样点数较少,一定程度上造成Simpson 多样性指数的单峰曲线格局并不明显。乔木层Pielou 均匀度指数的海拔分布格局为单峰分布,灌木和草本植物的Pielou 均匀度指数随海拔升高逐渐降低,如图5。
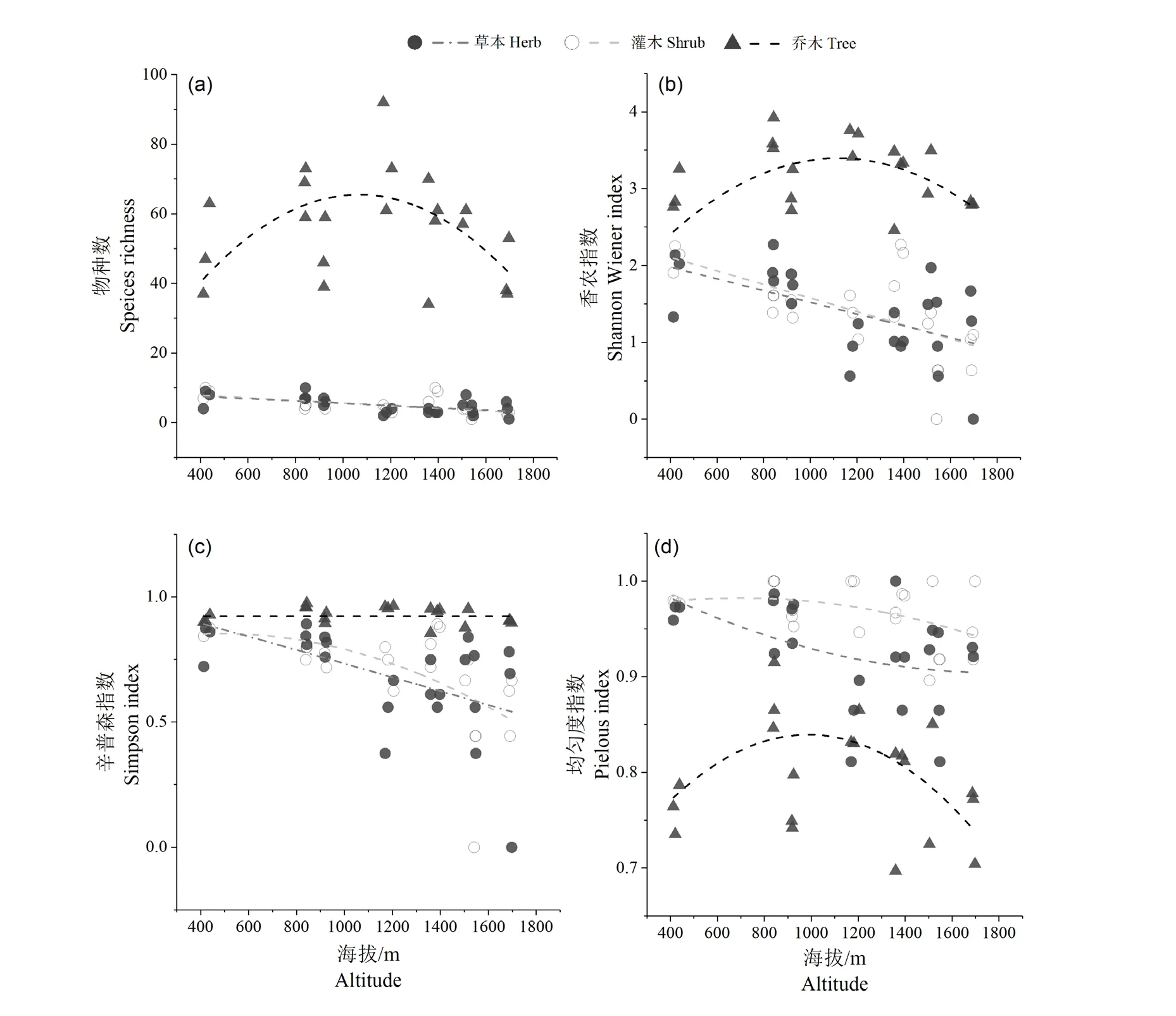
图5 南岭山地乔灌草物种多样性随海拔的变化Figure 5 Changes in species diversity of tree, shrub and grass along altitude at Nanling Mountains
3 讨论与结论
山地生态系统因具有独特的植被、土壤、水文等变化趋势,是研究植物群落生态规律和生态过程的理想生态系统。随海拔梯度变化,区域内环境因子发生剧烈变化,进而影响植物群落的物种组成和空间分布格局[3]。南岭国家级自然保护区内不同海拔梯度生长着不同植被类型。本文研究中植物群落内乔木、灌木和草本植物物种多样性指数随海拔梯度的升高表现不同的变化趋势。植物群落间结构和物种组成存在的差异性,间接可以反映出群落或生境的多样性[2,34]。
3.1 群落内物种组成是区分不同群落的重要特征。本研究发现随着海拔梯度的上升,南岭山地内植被类型变化依次为沟谷常绿阔叶林、山地常绿阔叶林、针阔混交林和山地常绿阔叶矮林。常绿阔叶树种在群落内占的比例随着海拔的增加呈现降低的趋势,而落叶阔叶树种的占比逐渐升高,符合植被的垂直地带性变化规律。海拔900 m 以下的沟谷内,乔木优势树种以广东润楠、华润楠等樟科润楠属为主。海拔900~1 360 m 的山体生长的常绿阔叶林,主要以甜槠、水青冈、鹿角锥、罗浮锥、米槠等壳斗科树种为优势树种。而针阔混交林内针叶树种主要以广东松和马尾松为主,阔叶树种以青冈和甜槠等为主。朱彪等人[20]的研究表明南岭山地一般山体较小,海拔不够高,难以形成云冷杉为代表的高山暗针叶林,因此山顶多为山地矮林,以野茉莉、青冈和冬青等常绿树种为优势种。从物种组成随海拔梯度的变化,侧面体现了以气温为主的环境因子随海拔升高的变化特征[35]。
3.2 随着海拔的增加,乔木层的最大树高(Hmax)、最大胸径(DBHmax)和胸高断面积和(BA)的变化曲线为单峰分布格局。立木密度(N)随海拔变化表现不明显的下降变化趋势,这与朱彪等人[20]在猫儿山的研究结果相似。谭一波等人[36]对猫儿山3 个海拔梯度的植物群落树木研究发现,树高随着海拔梯度的变化为单峰变化模式,峰值为海拔1 500 m 的常绿落叶阔叶混交林的乔木层,与本研究结果相似。最大树高和最大胸径峰值均在针阔混交林群落内的广东松植株,这可能与针阔混交林内植株个体密度较中低海拔群落小,以及针叶树对恶劣环境条件的抵抗力更强有关[37]。山地矮林群落个体为了减少风害造成的枝干机械损伤,表现出增加胸径和冠幅、降低株高与冠层厚度的自我保护的策略[38-39]。
3.3 南岭国家级自然保护区内的乔木层物种多样性随海拔的升高呈现“单峰曲线”变化趋势,灌木和草本层物种多样性随海拔升高分布格局为下降趋势,这与许多学者的研究结果相似[12,15,40-44]。乔木物种多样性的单峰分布格局与年平均降水的海拔分布格局类似,随海拔升高会先增加后降低,峰值位于八宝山内海拔1 170 m 的山地常绿阔叶林内,温度对南岭森林的乔木多样性格局的影响不大。低海拔的山地未表现很高的乔木物种多样性原因可能在于低海拔地区接近保护区内居民住所,受人为干扰影响相对较大。高海拔地区水热条件降低、风速较大,生境条件严酷,降低了部分物种的生存机会。
灌木和草本植物的物种多样性格局为随海拔升高递减,与年平均温度的海拔分布格局相似,说明灌木和草本植物分布更受温度限制。与秦浩[45]在山西关帝山发现的草本植物随海拔上升递增的格局并不一致,关帝山的草本植物多样性格局主要限制因子是降水。而与古兜山和莲花山上的灌木和草本海拔分布格局相似[47-48]。南岭山区降水丰富,水分并不是灌木和草本植物生长的限制因子。森林生态系统中,灌木和草本层位于森林垂直结构的下层,因此灌木和草本的多样性形成和分布格局受林冠植物的种类、结构和环境因子的综合作用,海拔分布格局多样[48-49]。
Liu 等[40]研究表明植物丰富度的分布受到水热条件的综合影响,温度、水分条件最理想的生境内可容纳最多的物种数。Pausas 和Austin[35]认为局域尺度上决定物种丰富度模式的主要因素是资源的可利用性和对水热条件等环境变量变化的响应,水热条件等环境变量对植物生长或资源可利用性有直接作用。在南岭山地观察到的物种丰富度分布格局也表明这可能是由于人为干扰加上温度和降水随海拔梯度升高而展现的非随机模式变化[50]。南岭地区生物多样性格局可为山地植物多样性垂直分布格局提供资料积累,为当地生物多样性保护和森林群落经营提供切实的科学依据。
致谢: 野外调查得到广东省科学院广州地理研究所黄明敏和南岭国家级自然保护区管理局的支持,在此致谢!
