竹节香附素A 通过PI3K/Akt/mTOR 信号通路对人宫颈癌HeLa 细胞增殖和凋亡的影响
2022-04-22张娟李雪蒋浩黄晓萍
张娟 李雪 蒋浩 黄晓萍
宫颈癌在全球女性常见恶性肿瘤中居第四位,在发展中国家常见恶性肿瘤中居第二位。当前的治疗策略包括手术切除、化学治疗和放射治疗,但疗效均有限。如何预防与治疗宫颈癌是全球医学工作者们共同关注的课题。竹节香附素A(RDA)是从常用的传统中草药海葵中提取的一种活跃三萜皂苷,通过抑制肿瘤细胞增殖和血管生成产生抗肿瘤活性,同时能诱导多种肿瘤细胞凋亡,如大肠癌细胞、胃癌细胞和肝癌细胞。磷脂酰肌醇3 激酶/蛋白激酶B/ 哺乳动物雷帕霉素靶蛋白(PI3K/Akt/mTOR)信号通路是一条常见的细胞信号通路,由于mTOR 能调节细胞代谢活动,因此被作为抑制肿瘤细胞生长的靶点。有研究者发现,RDA 可能通过激活p38MAPK 通路和抑制mTOR 活化诱导胃癌细胞凋亡。RDA 在宫颈癌中未有相关报道。因此,本研究探究RDA 对人宫颈癌HeLa 细胞增殖和凋亡的影响,并初步探讨其可能的作用机制。
材料与方法
一、主要材料
人宫颈癌HeLa 细胞,购自中国科学院典型培养物保藏委员会细胞库。RDA,纯度≥98%,购自上海源叶生物科技有限公司,批号20190203;胰岛素样生长因子-1(IGF-1),纯度≥98%,货号PHG0078,购自美国Gibco 公司。
二、主要试剂和仪器
高糖DMEM 培养基、胎牛血清,均购自美国Gibco 公司;B 细胞淋巴瘤2 家族蛋白(Bcl-2)抗体、β-actin 抗体、Bcl-2 相关X 蛋白(Bax)抗体、活化胱天蛋白酶-3(Cleaved-caspase-3)抗体、mTOR 抗体、PI3K 抗体、Akt 抗体、羊抗兔二抗均购自艾博抗(上海)贸易有限公司。
BA400Digital 数码三目摄像显微镜为麦克奥迪实业集团有限公司产品,Image-Pro Plus 6.0 图像分析软件为美国Media Cybernetics 公司产品,SpectraMAX Plus384 酶标仪为美谷分子仪器有限公司产品,Cytoflex 流式细胞仪为美国贝克曼库尔特公司产品。
三、方 法
1.细胞培养
将已经冻存HeLa 细胞株快速在37 ℃下解冻后,加入适量高糖DMEM 完全培养基(10%胎牛血清+0.5%青链霉素双抗),调整细胞密度为1×10/mL,在37 ℃、5%CO的培养箱中连续培养72 h,中途更换完全培养基2~3 次,然后进行传代备用。
2.实验分组
浓度筛选试验:将细胞分成5 组(0、5、10、20 和40 μmol/L),药物干预48 h。机制检测试验:将细胞分为空白对照组、IGF-1 组(100 ng/L)、RDA 组(20 μmol/L)与联合用药组(IGF-1 100 ng/L+RDA 20 μmol/L),各组药物均干预48 h。
3. CCK-8 法检测细胞增殖率
待HeLa 细胞培养至生长对数期,加入胰蛋白酶进行消化,使用完全培养基制成单细胞悬液,将细胞密度调整为5×10/L,每孔取100 μL 细胞悬液加入到96 孔细胞培养板中,每孔加入100 μL 完全培养基,置于37 ℃、5%CO的培养箱中培养24 h。待细胞完全生长贴壁后弃去培养基,使用磷酸盐缓冲液清洗2 次。将RDA 溶于完全培养基,调整终浓度为0、5、10、20 和40 μmol/L,每孔加入200 μL 含RDA 培养基,置于37 ℃、5%CO的培养箱中培养48 h。药物干预48 h 后,每孔加入10 μL CCK-8 溶液,继续在培养箱培养2 h,然后使用酶标仪测定在450 nm 波长下的吸光度值(A),以0 μmol/L 浓度的吸光度为对照值,根据公式计算培养48 h 的细胞增殖率,并计算半抑制浓度(IC),每组3 个复孔,结果取平均值。
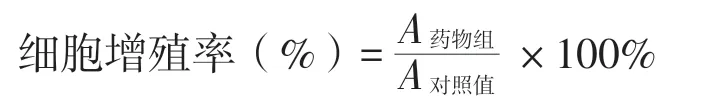
4.流式细胞术检测细胞凋亡
将处于生长对数期的HeLa 细胞用完全培养基制成1×10/mL 的细胞悬液,每孔取1 mL 细胞悬液加入6 孔板中,37 ℃、5%CO条件下培养24 h使细胞生长贴壁,分别按照前述步骤分组培养HeLa 细胞48 h,使用流式细胞仪检测细胞凋亡情况。每组3 个复孔,凋亡结果取平均值。
5.蛋白免疫印迹法检测各蛋白表达水平
分别按照前述步骤分组培养HeLa 细胞48 h,然后收集细胞行蛋白免疫印迹法检测细胞中Bcl-2、Bax、Cleaved-caspase-3、磷酸化-PI3K(p-PI3K)、p-Akt、p-mTOR 蛋白的表达量,并计算蛋白相对表达量。每组3 个复孔,蛋白表达结果取平均值。
四、统计学处理
使用SPSS 25.0 分析数据。统计数据使用表示,多组间均数比较采用单因素方差分析,多重比较采用Dunnett-t 检验;析因设计资料组间比较采用析因设计方差分析,交互效应有统计学意义时采用LSD-t 检验分析单独效应,P < 0.05 表示差异具有统计学意义。
结 果
一、不同浓度RDA 对HeLa 细胞凋亡的影响
CCK-8 结果显示,与0 μmol/L 组相比,5 μmol/L 浓度以上RDA 对HeLa 细胞有抑制作用(P < 0.05),抑制程度与RDA 浓度呈正相关,IC为20.90 μmol/L;细胞凋亡结果显示,5 μmol/L浓度以上RDA 对HeLa 有促进凋亡的作用(P <0.05),凋亡程度与RDA 浓度呈正相关,见图1、表1。

表1 不同浓度RDA 作用下HeLa 细胞的增殖率与凋亡率比较(,n=3)单位:%
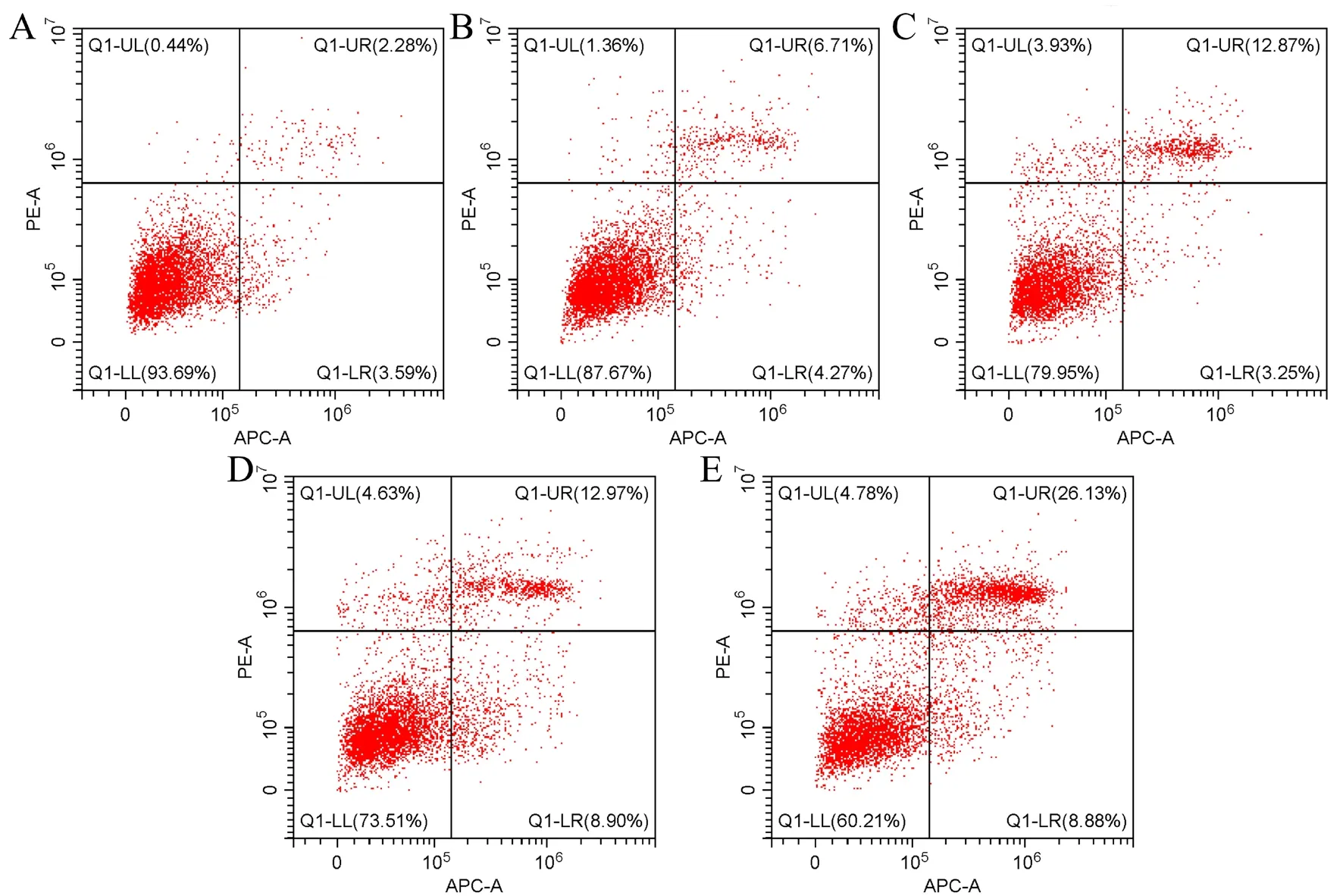
图1 不同浓度RDA 干预后HeLa 细胞的凋亡情况
二、不同浓度RDA 对HeLa 细胞凋亡相关蛋白的影响
蛋白免疫印迹检测显示,与0 μmol/L 组相比,20、40 μmol/L 组中Bax 和Cleaved-caspase-3 的表达量均升高,Bcl-2 的表达降低(P 均< 0.05),0、5 μmol/L RDA 对HeLa 中Bcl-2、Bax、Cleavedcaspase-3 凋亡蛋白表达无明显影响,10 μmol/L RDA 对Cleaved-caspase-3 蛋白表达无明显影响(P均> 0.05),见图2、表2。

表2 不同浓度RDA 作用下HeLa 细胞中Bcl-2、Bax 及Cleaved-caspase-3 的蛋白相对表达量比较(±s,n = 3)
三、RDA 对HeLa 细胞PI3K/Akt/mTOR 信号通路的影响
蛋白免疫印迹检测显示,与0 μmol/L 组相比,10、20 和40 μmol/L 组3 种蛋白磷酸化表达量均降低(P 均< 0.05)。5 μmol/L 组3 种蛋白磷酸化表达量有降低趋势,但p-Akt 和p-mTOR 与0 μmol/L 组比较差异无统计学意义(P 均> 0.05),见图2、表3。综合前述结果,采用40 μmol/L 的RDA 进行后续实验。
表3 不同浓度RDA 作用下HeLa 细胞中p-PI3K、p-Akt 及p-mTOR 的蛋白相对表达量比较(±s,n = 3)
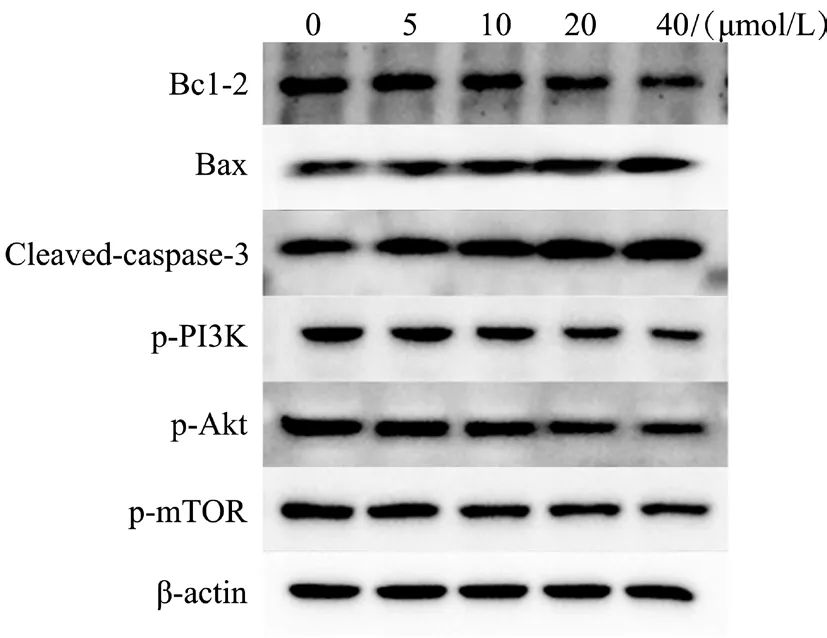
图2 不同浓度RDA 干预HeLa 后凋亡蛋白与PI3K/Akt/mTOR 信号通路蛋白的表达情况
四、激活PI3K/Akt/mTOR 通路后RDA 对HeLa 细胞的影响
检测与析因分析结果显示,IGF-1 和RDA 对细胞增殖率与凋亡率的主效应均有统计学意义,两者的交互作用对细胞增殖率(F = 11.028,P =0.006)与凋亡率(F = 48.494,P < 0.001)有影响;估算边际平均值后发现RDA 对细胞增殖率与凋亡率影响明显,见表4。
表4 IGF-1 与RDA 对HeLa 细胞增殖率与凋亡率的效应分析(±s,n = 3) 单位:%
与空白对照组相比,IGF-1 组细胞增殖率与凋亡率无明显变化(P > 0.05),RDA 组与联合用药组细胞的增殖率下降,凋亡率上升(P < 0.05);与IGF-1 组相比,RDA 组与联合用药组细胞的增殖率下降,凋亡率上升(P < 0.05),RDA 组抑制HeLa 细胞增殖与促进凋亡的效果比联合用药组更明显(P < 0.05),见表5、图3。

图3 激活PI3K/Akt/mTOR 信号通路后RDA 对HeLa 细胞凋亡的影响
表5 IGF-1 与RDA 对HeLa 细胞增殖率与凋亡率的影响(±s,n = 3) 单位:%
五、激活PI3K/Akt/mTOR 信号通路后RDA对HeLa 细胞中该信号通路蛋白表达的影响
析因分析结果显示,IGF-1 和RDA 对p-PI3K(F = 72.804,P < 0.001;F = 23.819,P = 0.001)、p-Akt(F = 36.510,P < 0.001;F = 5.317,P <0.001)、p-mTOR(F = 45.504,P < 0.001;F =11.982,P = 0.009)的主效应均有统计学意义,两者的交互效应对p-PI3K、p-Akt、p-mTOR 无影响(F = 0.272,P = 0.616;F = 1.291,P = 0.289;F =0.013,P = 0.912),估算平均边际值后发现RDA对p-PI3K、p-Akt、p-mTOR 影响明显,见表6。故主要解析各因素的主效应作用,其中IGF-1 与RDA对HeLa 细胞的PI3K/Akt/mTOR 信号通路影响均有统计学意义,IGF-1 对PI3K/Akt/mTOR 信号通路为激活效应,而RDA 对PI3K/Akt/mTOR 信号通路表现抑制效应,见表7、图4。

图4 激活PI3K/Akt/mTOR 信号通路后RDA 对HeLa 细胞p-PI3K、p-Akt 及p-mTOR 蛋白表达的影响
表6 IGF-1 与RDA 对HeLa 细胞中PI3K/Akt/mTOR 信号通路蛋白表达效应分析(±s,n = 3)
表7 激活PI3K/Akt/mTOR 信号通路后RDA 对HeLa 细胞p-PI3K、p-Akt 及p-mTOR 蛋白的影响(±s,n = 3)
讨 论
研究表明,RDA 可在体外抑制多种肿瘤细胞的生长,如肝癌、乳腺癌、胃癌及卵巢癌等,说明RDA 有着强大的抗肿瘤活性。由此推测RDA对宫颈癌可能有一定的抗癌作用。本研究采用人宫颈癌HeLa 细胞进行体外试验,从细胞凋亡率和PI3K/Akt/mTOR 信号通路等方面探讨RDA 对人宫颈癌细胞的抑制作用。研究结果显示,5~40 μmol/L 的RDA 对HeLa 细胞均有抑制作用,且抑制效果与RDA 浓度呈正相关,初步显示RDA 对HeLa 细胞具有明显的抑制效果。
Caspase 是一组在细胞凋亡过程起关键作用的酶,而Bcl 家族在抑制Bcl-2 或促进Bax 导致细胞死亡的途径中也起着关键作用。本研究显示,5~40 μmol/L 的RDA 能对HeLa 细胞产生促凋亡作用,且作用随RDA 浓度的升高而增强,同时,20~40 μmol/L 的RDA 能使HeLa 细胞中的Bcl-2蛋白表达降低,Bax 和Cleaved-caspase-3 蛋白表达升高,这说明RDA 不仅对HeLa 细胞既有抑制增殖作用,也有促进凋亡作用。
PI3K/Akt/mTOR 信号通路广泛存在于多种细胞信号转导通路中,是目前恶性肿瘤研究的热点之一。该通路在肿瘤细胞的能量代谢、细胞增殖、侵袭能力、细胞凋亡和细胞周期等生理活动中发挥重要作用。PI3K/Akt/mTOR 信号通路在许多人类恶性肿瘤(包括宫颈癌)中异常激活。p-Akt和p-mTOR 蛋白在50%~53%的宫颈腺癌中均有表达,说明这2 种蛋白磷酸化过表达可能是导致宫颈癌的原因之一。本研究显示,使用RDA 干预HeLa 细胞后,p-PI3K、p-Akt 和p-mTOR 这3 种蛋白的相对表达量均降低,且抑制作用随RDA 浓度的升高而增强。由此说明,RDA 能通过抑制PI3K/AKT/mTOR 信号通路,降低HeLa 细胞增殖率。
IGF-1 通过活化Akt 激活下游信号通路,是胞外信号调节激酶1/2(ERK1/2)和PI3K/Akt 通路的强效刺激因子。有研究者发现,IGF-1 促进多种肿瘤细胞的有丝分裂、转移和抗凋亡,有助于肿瘤细胞的维持和促进肿瘤的进展。因此本研究探讨在PI3K/Akt/mTOR 信号通路激活的环境下,RDA 是否能发挥同样的抗癌效果。结果显示,IGF-1 和RDA 存在交互作用,RDA 的单独效应随IGF-1 变化,说明RDA 可能对IGF-1 诱导细胞增殖与抑制细胞凋亡的效果有抑制作用。与IGF-1 组相比,联合用药组能明显降低细胞增殖率且提高细胞凋亡率,但效果不如单用RDA,说明在IGF-1的干预下,RDA 仍然能发挥抑制肿瘤细胞生存的效果。进一步研究发现,IGF-1 和RDA 的交互作用并没有对p-PI3K、p-Akt 和p-mTOR 蛋白相对表达量产生影响,说明IGF-1 此时的作用只是单纯的PI3K/Akt/mTOR 信号通路激动剂,并未对RDA产生直接影响。与空白对照组相比,IGF-1 组的p-PI3K、p-Akt 和p-mTOR 蛋白相对表达量升高,说明IGF-1 激活了HeLa 细胞中的PI3K/Akt/mTOR信号通路。而RDA 能降低IGF-1 干预后p-PI3K、p-Akt 和p-mTOR 蛋白相对表达量,说明无论PI3K/Akt/mTOR 信号通路是处于正常状态还是异常激活状态,RDA 都能抑制HeLa 细胞中PI3K/Akt/mTOR信号通路中PI3K、Akt 和mTOR 蛋白的磷酸化,从而诱导HeLa 细胞死亡,阻止肿瘤细胞的增殖。
综上所述,RDA 能对人宫颈癌HeLa 细胞产生抑制增殖与促进凋亡的作用,其作用机制是通过抑制PI3K/Akt/mTOR 信号通路,降低p-PI3K、p-Akt 和p-mTOR 蛋白的表达,产生抗癌的效果。
