紫云英与油菜混播对稻田土壤N2O排放及相关功能基因丰度的影响
2022-04-21杨滨娟李萍胡启良黄国勤
杨滨娟,李萍,胡启良,黄国勤
紫云英与油菜混播对稻田土壤N2O排放及相关功能基因丰度的影响
杨滨娟,李萍,胡启良,黄国勤
江西农业大学农学院/江西农业大学生态科学研究中心,南昌 330045
【】探究绿肥混播对稻田土壤N2O的减排效果和机制,筛选既高产稳产,又减排的适合南方双季稻区发展应用的绿肥混播种植模式。比较不同紫云英与油菜混播比例(单播紫云英(CK1)、单播油菜(CK2))、3/4紫云英+1/4油菜(3/4M + 1/4R)、1/2紫云英+1/2油菜(1/2M + 1/2R)、1/4紫云英+3/4油菜(1/4M + 3/4R))对稻田N2O排放规律及相关功能基因的影响。(1)作物种类影响N2O排放。各处理的N2O排放规律基本一致,其中冬作季N2O排放主要集中在冬季绿肥的盛花期,早晚稻季主要集中在水稻的分蘖、灌浆和成熟期。总体来看,稻田N2O排放主要集中在水稻季,早稻季的N2O排放量低于晚稻季;2018年稻田N2O排放通量总体比2019年高。(2)各处理的早稻N2O累积排放量差异均不显著。连续两年单播处理的N2O累积排放量低于混播各处理,其中2019年处理CK1最低,处理3/4M+1/4R的N2O周年累积排放量在各混播处理中最低。(3)整体来看,晚稻土壤的-AOA和-AOB基因拷贝数比早稻要高,-AOA基因拷贝数比-AOB高1—2个数量级。晚稻的土壤、、基因拷贝数比早稻要高,其中基因拷贝数比和高1—2个数量级。紫云英与油菜混播提高了土壤-AOA和-AOB基因丰度,-AOA基因为优势功能基因,而且提高了土壤、、基因丰度,基因占主导地位。反硝化基因拷贝数比硝化基因拷贝数高1—3个数量级,反硝化作用对N2O排放贡献更大。“3/4紫云英+1/4油菜-早稻-晚稻”种植模式在保证高产和高肥力的情况下,N2O排放量相对较低,是较理想的种植模式。
紫云英;油菜;绿肥混播;氧化亚氮;功能基因;稻田
0 引言
【研究意义】双季稻田冬种绿肥的稻田三熟制是我国南方一种传统的种植模式[1-2]。但目前普遍存在绿肥种子价格高、成苗率低、鲜草量少等问题,制约了绿肥种植技术的直接经济效益,导致农民种植积极性不高。冬季绿肥紫云英(含氮量较高)和油菜(含磷钾较高)混播不但能满足水稻不同生长时期对养分的需求,而且能充分利用各自生育上的时间差和生长上的空间差,从而提高光能和土地的利用率[3-4],其可持续能值指数为0.0315,环境负载率较低[5]。因此,在南方双季稻区合理开发紫云英、油菜混播模式,对发展冬季绿肥、高效利用农业资源、改善土壤性状、提高经济和生态效益具有重要意义。【前人研究进展】大气中N2O对温室效应的贡献率仅为5%[6],但其增温效应约为CO2的296—310倍,在大气中的存留时间长达150年左右[7]。水稻生长季排放的N2O占稻田总排放量的7%—11%[8]。有报道指出在淹水稻田等厌氧环境下,土壤硝化作用受到限制,N2O主要来源于土壤反硝化作用。土壤农田管理措施和土壤性质都会对、、型反硝化细菌微生物丰度和多样性产生一定影响[9],尤其是、、群落对于土壤pH和土壤速效氮较为敏感[10]。冬季绿肥在生长发育过程中会产生一系列复杂的生理生化作用,改变了以土壤微生物群落结构和数量及其活性等微生态环境为主的根际土壤环境[11],直接或间接影响了土壤中硝化反硝化过程,从而影响土壤中N2O的排放和各形态氮库容量及其相互转化与平衡。另外,农田作物多样性改变,增加了农田生物多样性[12]。紫云英、油菜混播相比常规冬种绿肥模式,由于根系分布的范围不同,根系分泌物、根系微生物区系、生长特性等诸方面不同,可能会对稻田土壤氮循环微生物群落产生不同的影响。王艳秋等[13]研究表明,在江西黄泥田中,紫云英翻压还田能提高土壤硝化强度及硝化势,进而显著提高-AOA和-AOB的基因丰度。高嵩涓[14]研究结果显示,冬种紫云英翻压还田后增加了酸性红壤中的-AOA-AOB和的基因丰度,紫云英与化肥配施能提高土壤中的基因丰度。【本研究切入点】但绿肥混播模式下稻田N2O排放及土壤相关硝化与反硝化基因丰度的研究较少。【拟解决的关键问题】因此,开展紫云英、油菜混播对稻田土壤N2O相关功能基因丰度的影响,为减少稻田氧化亚氮排放和构建氧化亚氮等温室气体减排的稻作制度提供重要理论指导与技术支撑,也为紫云英、油菜混播技术的推广应用以及南方稻区生态系统可持续发展提供理论依据。
1 材料与方法
1.1 试验地概况
试验从2017年9月开始,在江西农业大学科技园水稻试验田(28°46′N,115°55′E)进行。试验地属于亚热带季风性湿润气候,年均太阳总辐射量为4.79×1013J·hm-2,年均日照时数为1 852 h。光照分布与水稻生长旺季基本同步,对水稻生产有利。≥0℃的年积温达6 450℃,年降水量1 624 mm,年平均气温17.1—17.8℃。供试土壤为发育于第四纪的红黏土,为亚热带典型红壤分布区。试验前表层(0—15 cm)土壤pH 5.29,有机质38.87 g·kg-1,全氮2.90 g·kg-1,碱解氮137.38 mg·kg-1,全磷0.56 g·kg-1,全钾21.24 g·kg-1,有效磷10.68 mg·kg-1,速效钾36.22 mg·kg-1。
1.2 试验设计
试验设5个处理,3次重复,随机区组排列,共15个小区,小区面积16.5 m2(5.5 m×3 m),小区之间用水泥埂隔开,试验前各小区土壤肥力均匀一致。两边设有保护行,保护行宽度为30 cm。具体试验处理为:单播紫云英(CK1);单播油菜(CK2);3/4紫云英+ 1/4油菜(3/4M + 1/4R);1/2紫云英+ 1/2油菜(1/2M + 1/2R);1/4紫云英+ 3/4油菜(1/4M + 3/4R),其他季节的种植模式均为双季稻。
紫云英品种为余江大叶籽、油菜为德油5号、早稻为中嘉早17、晚稻为天优华占。
田间管理措施:(1)冬季绿肥。2017—2018年:紫云英2017年10月2日撒播,不施肥;油菜2017年11月12日撒播,不施肥;2018年4月5日进行冬季绿肥测产。2018—2019年:紫云英2018年10月1日撒播,不施肥;油菜2018年11月11日撒播,不施肥;2019年4月7日进行冬季绿肥测产。单播紫云英播种量均为22.5 kg·hm-2,单播油菜播种量均为7.5 kg·hm-2,紫云英与油菜混播处理按照试验设计要求的比例换算相应的播种量。所有冬季绿肥在早稻移栽前15 d全部收割翻压还田,沤8—10 d即可进行常规耙田、施肥和早稻移栽。在翻压时先适度截断油菜茎秆,然后将紫云英和油菜同向翻压,使土壤完全覆盖紫云英和油菜植株,然后封住稻田排水口沤制,沤制时保持田面有水且最高水位不高于1.25 cm。(2)双季稻:2018年早稻于2018年3月28日播种,4月26日施基肥,4月27日以行距为0.2 m,株距为0.2 m移栽,5月5日追加分蘖肥,5月25日开始排水晒田,6月8日追加穗肥,7月16日收获;晚稻于2018年6月23日播种,7月24日施基肥,7月25日以行距为0.2 m,株距为0.2 m移栽,8月5日追加分蘖肥,8月25日开始排水晒田,9月4日追加穗肥,10月27日收获。2019年早稻于2019年3月27日播种,4月22日施基肥,4月26日以行距为0.2 m,株距为0.2 m移栽,5月8日追加分蘖肥,5月30日开始排水晒田,6月14日追加穗肥,7月24日收获;晚稻于2019年7月1日播种,8月2日施基肥,8月3日以行距为0.2 m,株距为0.2 m移栽,8月11日追加分蘖肥,8月30日开始排水晒田,9月14日追加穗肥,11月1日收获。
化肥种类及用量参照当地常规施肥:所有作物施用化肥均为尿素(N 46%),钙镁磷肥(P2O512%),氯化钾(K2O 60%)。水稻当地施肥常规用量为180 kg N·hm-2、90 kg P2O5·hm-2、120 kg K2O·hm-2。氮肥按基肥﹕分蘖肥﹕穗肥= 3﹕4﹕3施用;磷肥全部作基肥,一次性施入;钾肥按分蘖肥﹕穗肥=7﹕3施用。早晚稻收获后所有处理秸秆均不还田。
1.3 测定项目与方法
1.3.1 土壤N2O 采用静态暗箱-气相色谱法测定田间小区温室气体排放。静态箱长、宽、高均为0.5 m,采样箱体由304不锈钢材质制成,箱体高度随水稻高度而增加(拔节期前放置一层0.5 m,拔节期后放置两层共1 m)。采样箱内顶部装有12V小风扇,防止箱内气体不均匀。箱子顶部设有3个小孔,分别为风扇电池接口、温度计插口、采气孔,采气孔插入硅胶管然后用注射器连接,用三通阀控制开关。外部包有白色海绵罩,防止太阳照射导致箱内温度变化过大。绿肥生长季10—15 d采样1次,水稻生长季一般7 d采样1次。采样时间为上午9:00—11:00,采样前将采样箱体置于事先埋好的底座上,采样时用50 mL注射器来回抽5—10次以混匀气体,随后抽取气体样品到气袋中,4个连续样品分别在箱体密闭后按0、10、20和30 min的时间间隔用50 mL的注射器采集气体,3 d内利用A7890b安捷伦气相色谱(美国安捷伦科技有限公司)完成N2O浓度测定,其中,N2O检测器ECD检测温度为300℃,柱温60℃,载气为99.99%高纯氩/甲烷气(95%氩气+5%甲烷),流速40 mL·min-1。
(1)气体排放通量计算公式为:
F=ρ×h×dc/dt×273/(273+T)
式中,F为气体排放通量(mg·m-2·h-1或μg·m-2·h-1),ρ为标准状态下气体的密度(kg·m-3),h是采样箱的净高度(m),dc/dt为单位时间内采样箱内气体的浓度变化率,T为采样过程中采样箱内的平均温度(℃),273为气态方程常数[15]。
(2)N2O累积排放量(EN2O,kg N·hm-2)的计算公式[16]如下:
EN2O=∑(fi+fi+1)/2×(ti+1-ti)×24×10-2
式中,f代表N2O气体通量(mg N2O-N·m-2·h-1),i代表测量次数,(ti+1-ti)代表两次邻近测量天数。
1.3.2 土壤功能基因丰度 分别于2019年5月12日(早稻分蘖期)、7月24日(早稻成熟期)、8月14日(晚稻分蘖期)、11月1日(晚稻成熟期)随机在各小区选取5个点,采集0—15 cm土层土样进行混匀,手工挑去砾石和植物残体等,然后用干冰冷冻保存,样品委托武汉天一辉远生物科技有限公司测定。
本试验以土壤中氨氧化古菌基因、氨氧化细菌基因、亚硝酸盐还原酶()基因和一氧化二氮还原酶()基因的拷贝数分别表示AOA、AOB 、和基因型丰度。利用荧光定量PCR技术依次检测AOAAOB和基因拷贝数,即用SYBR® Select Master Mix(2X)试剂盒,在荧光定量PCR仪(Applied Biosystems StepOnePlus™ Real-Time System)上进行绝对定量PCR的分析,具体PCR引物如表1。定量 PCR 标准曲线采用含有AOAAOB和基因的克隆进行制备。首先采用特定引物分别扩增目的基因,构建克隆文库后,将含有目的基因的克隆在培养基中过夜培养,提取质粒纯化并测定质粒浓度,根据摩尔常数计算标准品基因的拷贝数,取标准品质粒稀释至109,然后取10 µL加入到90 µL无菌水中,依次10倍梯度稀释得到108、107、106、105、104、103,6个浓度梯度按照定量PCR中引物信息实验条件制作标准曲线。取qPCR96Well,配制反应体系,每个反转录产物配制3管。定量PCR 扩增反应体系为20 µL,包括2xqPCRmix10 µL、上下游引物(5 pmol·µL-1)各1 µL、DAN模板2 µL、灭菌水6 µL。试验控制扩增率在83.0%—105.0%。将得到的结果以标准质粒拷贝数的对数值为纵坐标,以不同浓度质粒的Cp值为横坐标建立出标准曲线,并根据标准曲线计算基因丰度。

表1 实时荧光定量PCR扩增引物
1.4 数据分析
采用 Microsoft Excel 2010 处理数据;StepOne Software v2.3软件分析功能基因丰度数据;SPSS19.0软件进行统计分析。
2 结果
2.1 不同混播比例对稻田土壤N2O排放的影响
2.1.1 稻田N2O排放通量的动态变化 2018年N2O的总体排放量比较少,各处理的N2O排放规律基本一致。冬作季出现了2个排放高峰(2017年11月26日和2018年4月1日),峰值分别为处理1/2M+1/2R的48.49 µg·m-2·h-1和处理CK2的30.09 µg·m-2·h-1,而两个时期排放最低的是处理3/4M+ 1/4R和1/2M+1/2R,分别低于峰值67.5%和66.5%。在早稻季和晚稻季均出现了3个排放高峰,而全年N2O的排放最高峰出现在晚稻季的10月14日,峰值为处理1/4M+3/4R的111 µg·m-2·h-1,排放最低的为CK2的43.37 µg·m-2·h-1。总体来看,单播处理的N2O排放量要低于混播各处理,混播处理中3/4M+ 1/4R排放量较低。
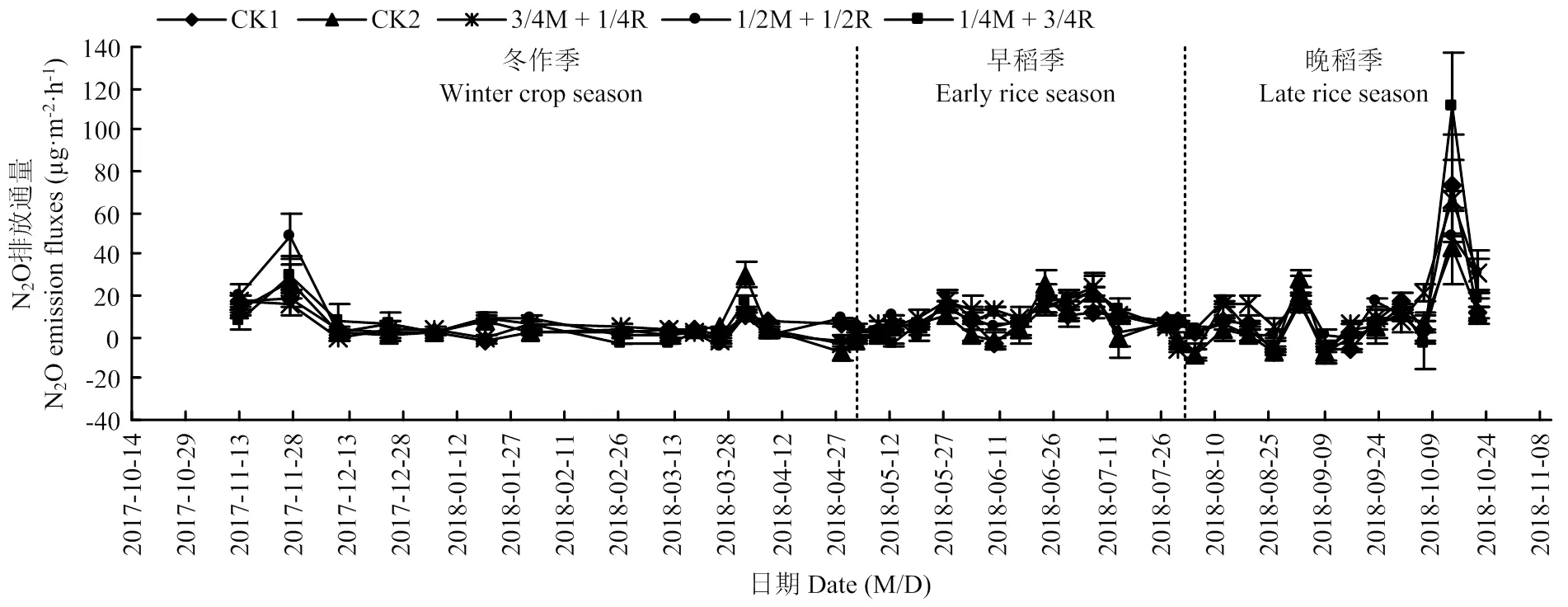
数据为2017年冬作起至2018年晚稻收获后 The data is between winter crop in 2017 and harvest of the late rice in 2018
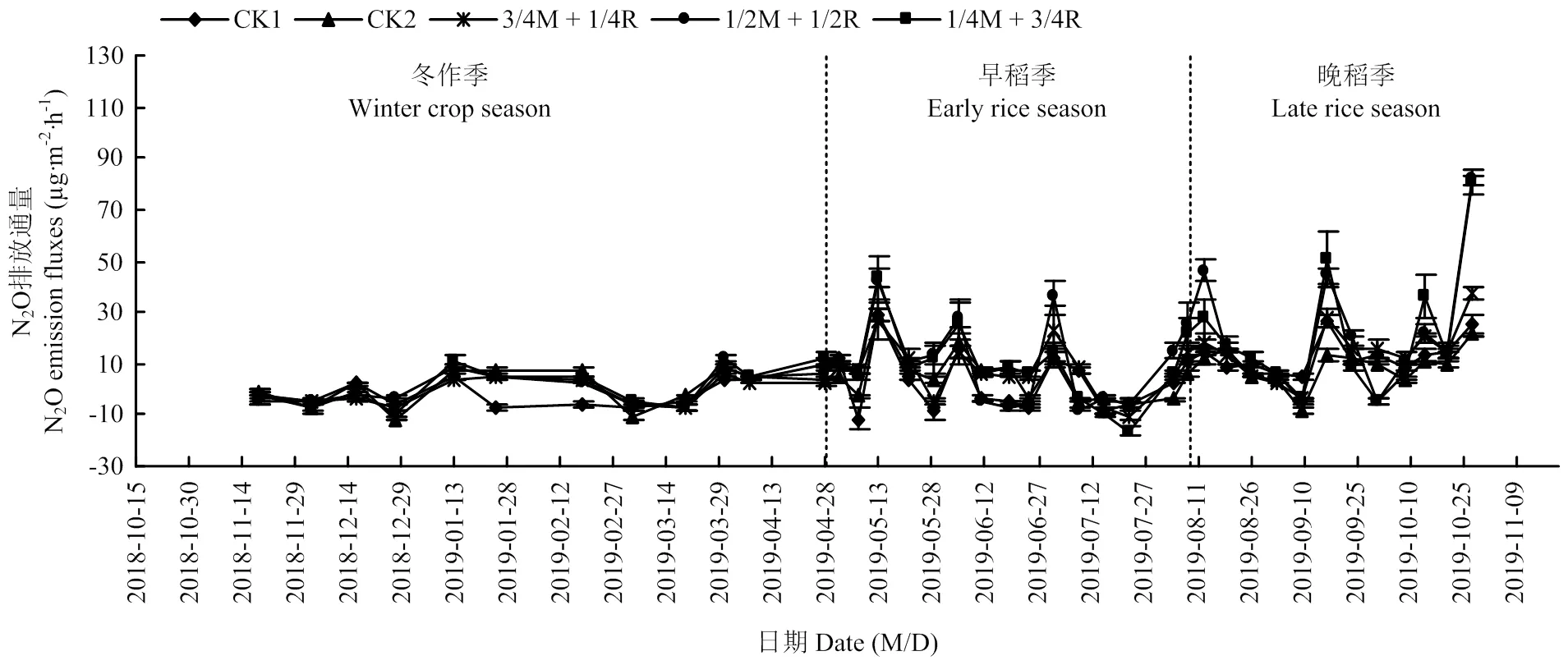
数据为2018年冬作起至2019年晚稻收获后 The data is between winter crop in 2018 and harvest of the late rice in 2019
2019年稻田N2O排放通量总体低于2018年。冬作季各处理N2O排放通量很低,排放范围为-11.46—12.52 µg·m-2·h-1。冬作季也出现了2个排放高峰(1月12日和3月30日),峰值分别为处理1/4M+3/4R的11.24 µg·m-2·h-1和处理1/2M+1/2R的12.52 µg·m-2·h-1。早稻季出现了3个排放高峰,晚稻季比2018年多了1个排放小高峰,出现了4个排放高峰,但2019年晚稻季排放通量总体要低于2018年。总体来看,与2018年趋势一致,单播处理的N2O排放量要低于混播各处理,混播处理中的3/4M+1/4R排放量较低。
2.1.2 稻田N2O累积排放量 由表2可知,2018年冬作季,混播各处理N2O累积排放量均高于处理CK1,处理1/2M+1/2R最高,处理CK1累积排放量最低,且除了处理3/4M+1/4R外,与其他处理间差异显著(<0.05);但早晚稻各处理间差异不显著(>0.05)。总的来看,两个对照的N2O累积排放量要低于混播各处理,其中处理CK2的累积排放量最低,比其他处理分别低1.8%、20.6%、27.0%、22.9%。2019年,冬作季各处理累积排放量很低,其中CK1最低为-0.11 kg·hm-2,且与其他处理存在显著差异(<0.05);晚稻季,处理CK2最低,处理1/2M+1/2R最高,除了处理1/4M+3/4R外,与其他处理间差异显著(<0.05),但早稻季各处理间差异不显著(>0.05)。总的来看,单播N2O累积排放量要低于混播各处理,处理CK1低于其他处理14.8%—61.0%,处理CK1、CK2、3/4M +1/4R与处理1/2M+1/2R、1/4M + 3/4R间差异显著(<0.05)。综合两年数据可以看出,2019年的累积排放量低于2018年,作物种类会影响N2O的排放。冬种不同作物还田对早稻N2O排放没有显著影响。单播处理的N2O累积排放量低于混播各处理,其中2018年处理CK2的周年N2O的累积排放量最低,2019年处理CK1最低,处理3/4M+1/4R的N2O周年累积排放量在各混播处理中最低。
2.2 不同混播比例对土壤硝化和反硝化功能基因丰度的影响
2.2.1基因丰度 由图3可知,土壤-AOA基因各处理各时期间差异均不显著(>0.05)。由图4可知,处理CK2的土壤AOB基因拷贝数在早稻分蘖期(5月12日)、成熟期(7月24日)和晚稻成熟期(11月1日)均达到最大,但晚稻成熟期各处理间差异不显著(>0.05)。处理CK2在早稻分蘖期,除了处理1/2M+1/2R,显著高出其他处理182.1% —530.0%(<0.05);在成熟期达到7.37×105copies·g-1,显著高于其他处理(<0.05)。晚稻分蘖期(8月14日)的土壤AOB基因拷贝数较早稻分蘖期有所增加,处理1/2M + 1/2R最高,显著高出两个对照处理137.1%、216.8%(<0.05)。整体来看,晚稻土壤的AOA和AOB基因拷贝数比早稻要高,AOA基因拷贝数比AOB高1—2个数量级。
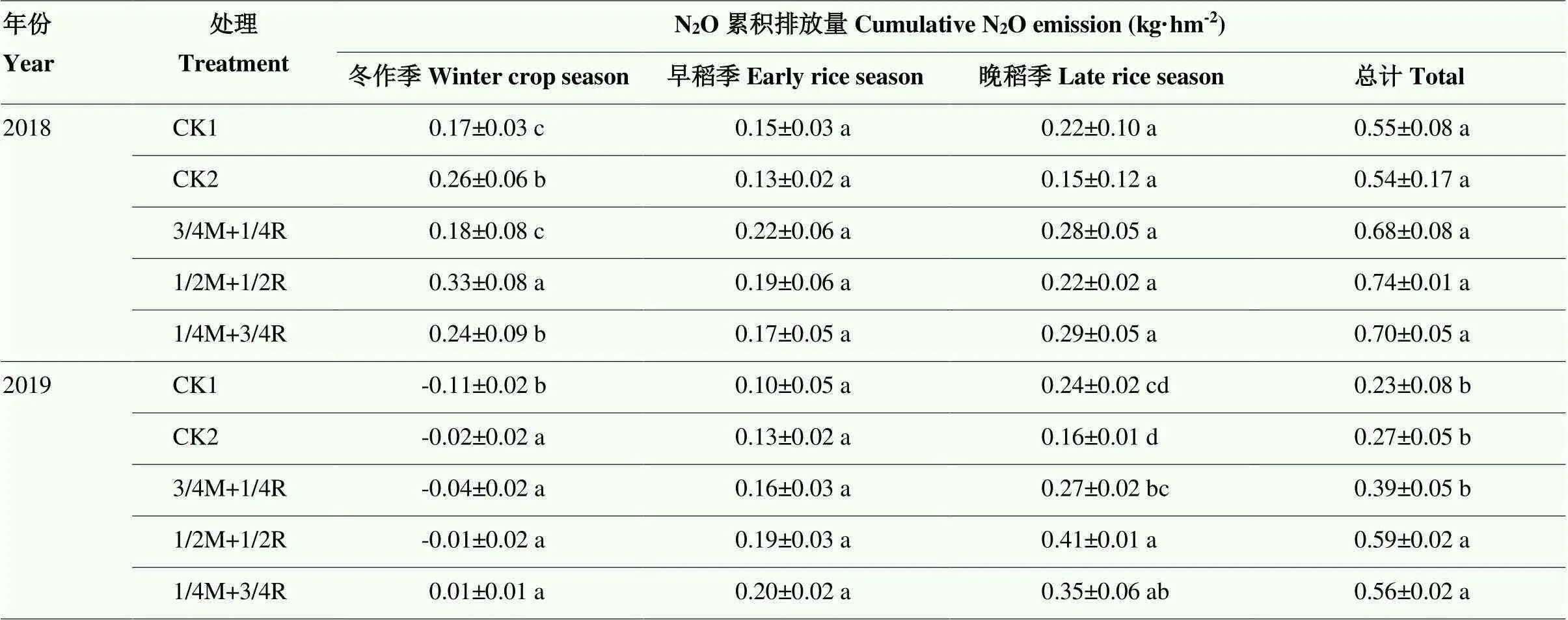
表2 各处理N2O累积排放量
同列不同的字母分别表示差异达5%显著水平。下同 The different letter in same column indicated significantly at 5% levels. The same as below
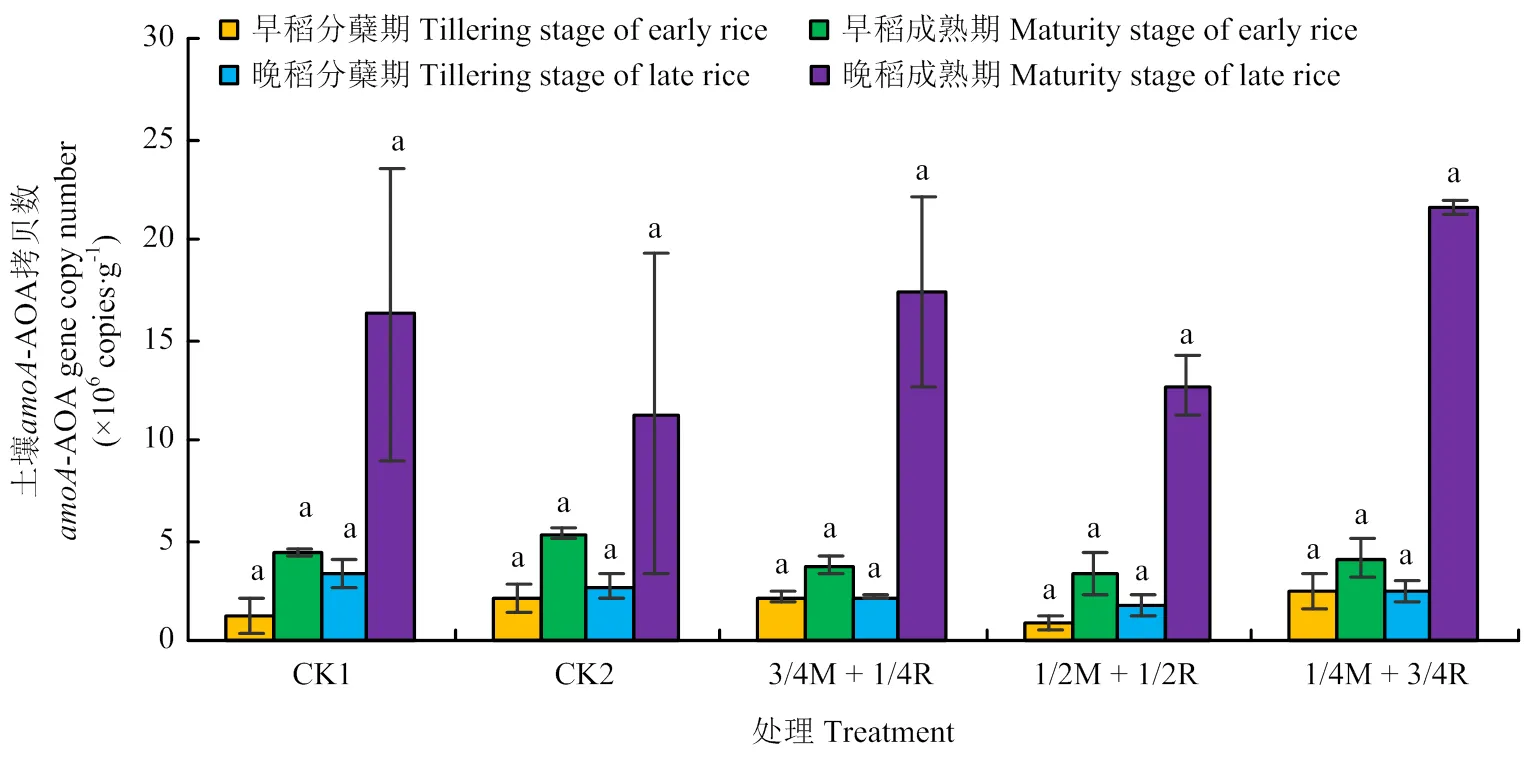
图3 土壤amoA-AOA基因丰度变化
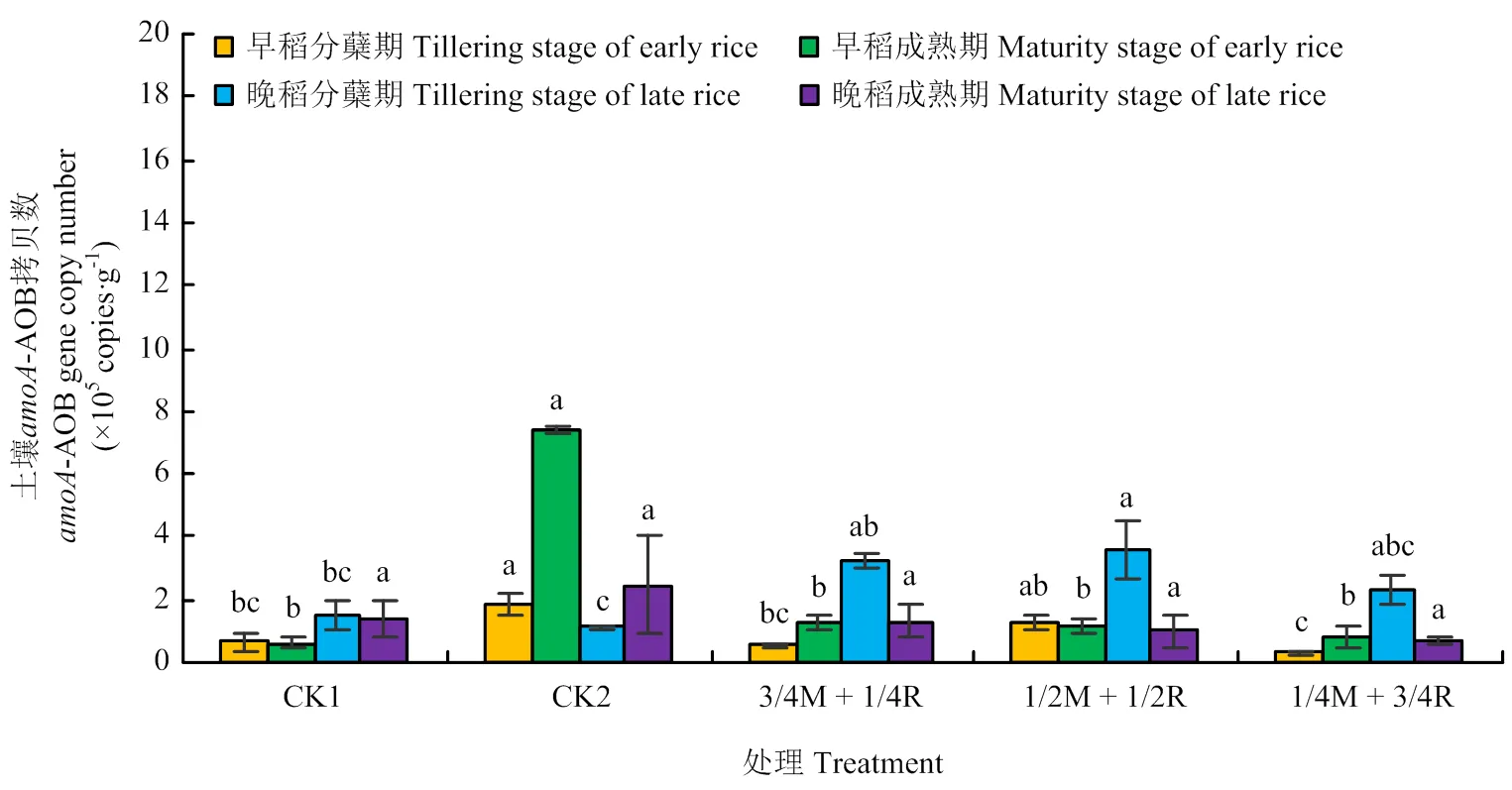
图4 amoA-AOB基因丰度变化
2.2.2、、基因丰度 由图5可知,早稻分蘖期和晚稻分蘖、成熟期的土壤基因拷贝数均是处理1/4M+3/4R达到最大,但早稻分蘖期和晚稻成熟期各处理间差异不显著(>0.05),在晚稻分蘖期处理1/4M+3/4R与处理1/2M+1/2R达到显著差异(<0.05)。早稻成熟期的土壤基因的拷贝数处理CK2最高,但仅与处理3/4M+1/4R间差异显著(<0.05)。土壤基因的拷贝数总体表现为晚稻高于早稻,处理1/4M+3/4R表现较好。由图6可知,早稻分蘖期、晚稻分蘖期和晚稻成熟期的土壤基因拷贝数各处理间差异不显著(>0.05);早稻成熟期是CK2达到最大,且与其他处理间差异显著(<0.05)。由图7可知,早稻分蘖期和晚稻分蘖、成熟期的土壤基因拷贝数各处理间均差异不显著(>0.05)。早稻成熟期CK2处理达到最大,为7.06×107copies·g-1,且显著高出其他处理43.5%—106.4%(<0.05)。总体来看,晚稻的土壤基因拷贝数比早稻要高,其中基因拷贝数比和高1—2个数量级。反硝化基因拷贝数比硝化基因拷贝数高1—3个数量级。其中处理1/4M+3/4R和CK2表现较好。
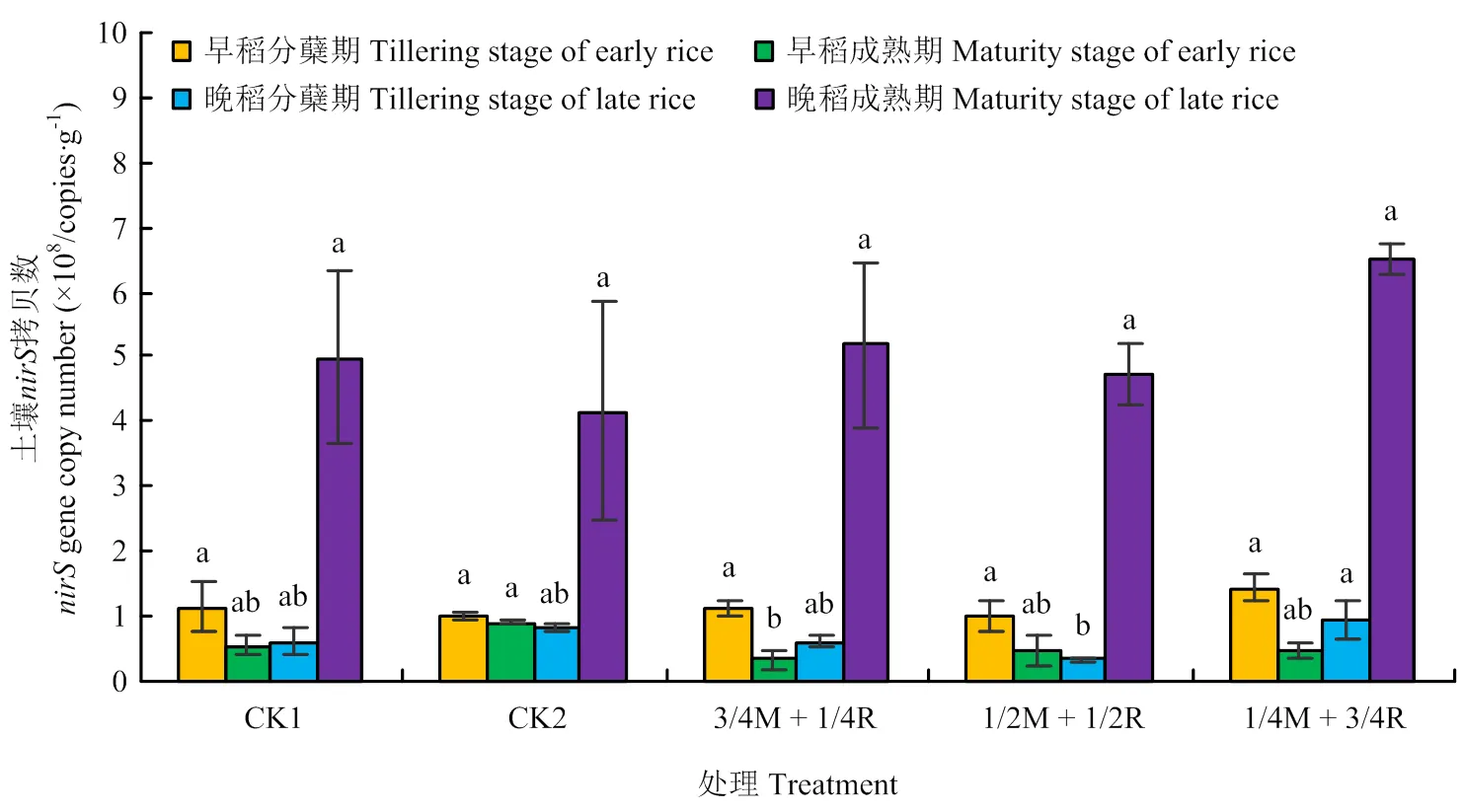
图5 土壤nirS基因丰度变化
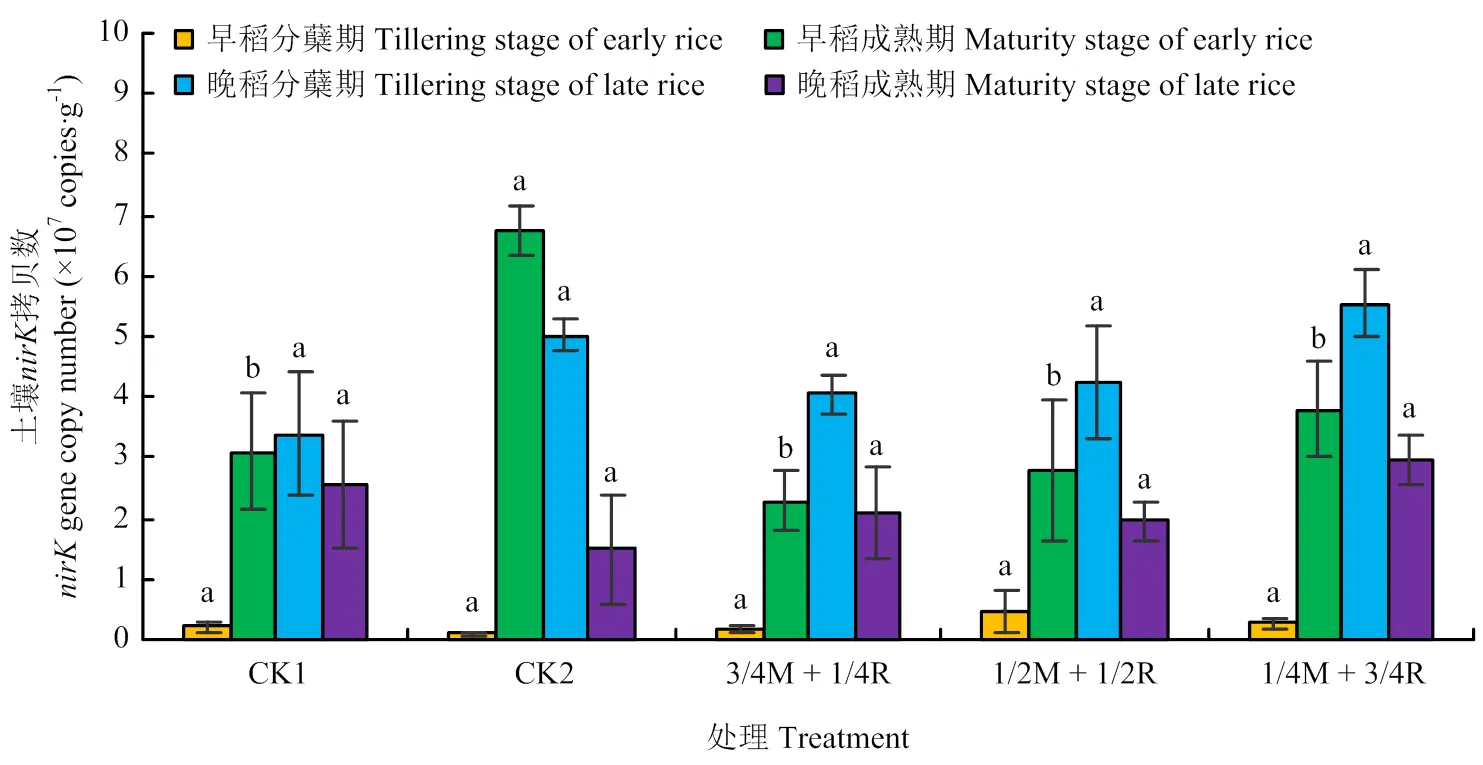
图6 土壤nirK基因丰度变化
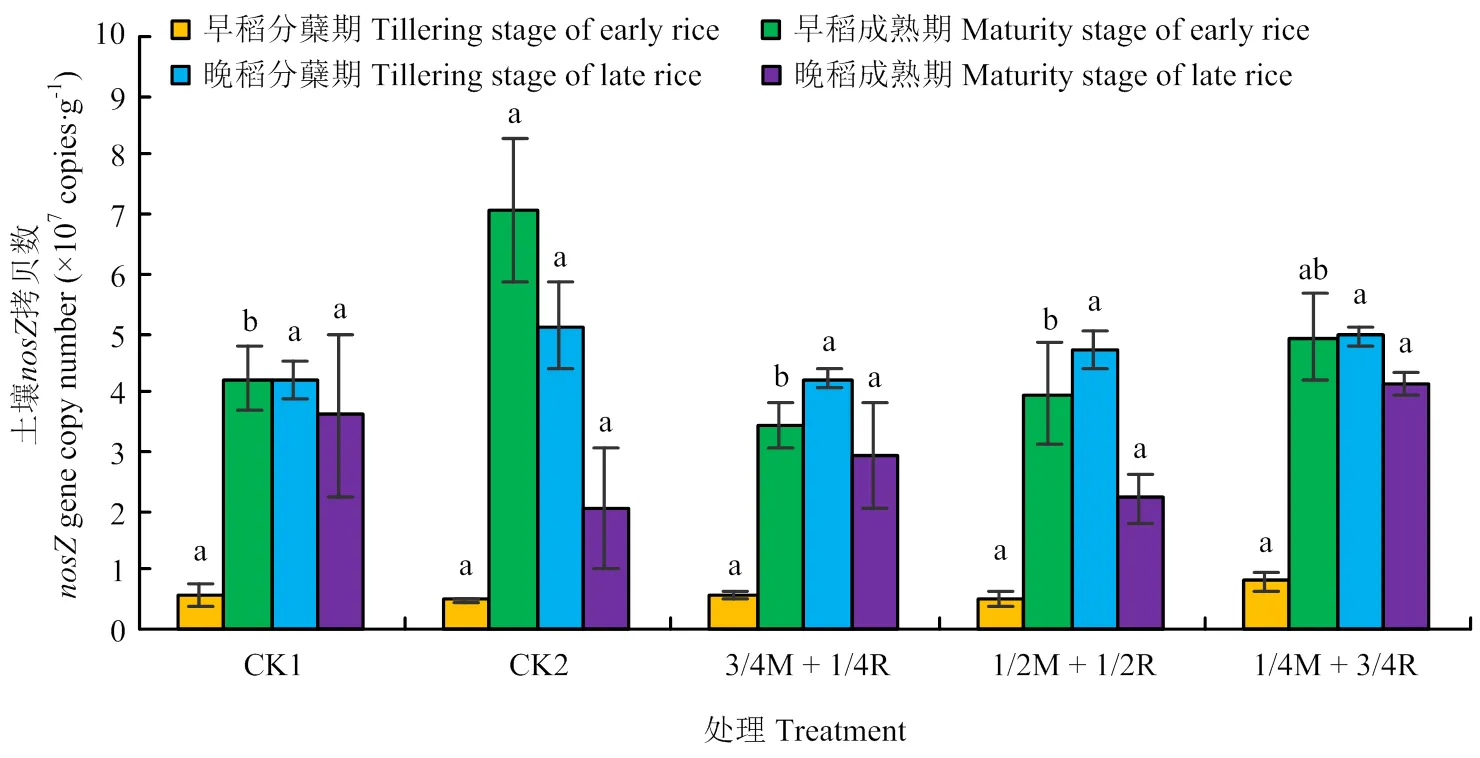
图7 土壤nosZ基因丰度变化
3 讨论
3.1 稻田N2O排放主要影响因素
稻田N2O排放主要受田间水分管理、作物种类及肥料施用等的影响[22-23]。冬作季N2O的排放主要集中在冬季绿肥的盛花期,因为当温度达到土壤微生物的最适温度时,能够刺激微生物活动,从而促进N2O的排放,这与唐海明等[24]的研究结果一致。而水稻季N2O的排放主要集中在水稻的分蘖、灌浆和成熟期,主要是因为稻田灌溉-排水晒田、下雨等造成的干湿交替环境及施肥增加了N2O的排放[25-26],这与张岳芳等[27]、田伟等[28]的研究结果一致。GOLDBERG等[29]研究发现,当土壤含水量较低时会出现N2O负排放。本研究中,N2O排放量为负值多集中在冬作季,因为与水稻季相比,冬季土壤多为干旱环境并且冬季温度较低,导致出现净吸收。本试验结果还表明,2019年各处理的周年累积排放量比2018年降低了20.0%—58.2%,说明冬季绿肥翻压还田可以降低N2O的排放,这与常单娜等[30]、聂江文等[31]研究结果相似,但还需要长期的监测。唐海明等[32]研究表明,不同冬季作物种类会影响稻田N2O的排放。本研究中,绿肥混播各处理的N2O排放总量比单播要高,一方面是因为绿肥翻压还田后,混播的腐解速率要高于单播[33],释放更多的养分,另一方面是由于混播能够改善土壤化学性状,提高土壤肥力,为土壤提供充足的营养条件,从而使混播的碳氮比更能满足土壤微生物的需求,为硝化与反硝化作用过程提供了丰富的基质,从而增加了N2O的排放。但在各混播处理中3/4M+1/4R的N2O周年累积排放量最低。另外,本研究N2O的累积排放量高于朱波等[34]、纪钦阳等[35]的研究结果,低于钟川等[36]的研究结果,这可能与作物种类、种植模式、施肥方式有关。
3.2 稻田N2O相关功能基因丰度
微生物转化过程主要是含有氧化亚氮还原酶(NOS)基因的反硝化微生物将N2O还原为N2的过程。氧化亚氮还原酶(NOS)在反硝化过程中主导N2O的还原,因此以氧化亚氮还原酶基因()为靶基因揭示微生物种群的N2O消耗还原能力是较直观的[37]。本研究也得出类似结果,晚稻土壤基因拷贝数比早稻要高,其中基因拷贝数比和高1—2个数量级。本试验土壤为酸性土壤,pH在4.85—5.55之间,而AOA的丰度比AOB高1—2个数量级,说明在酸性环境中AOA基因丰度高于AOB,且在硝化作用中AOA占比更高,这与李双双等[38]研究结果类似。土壤NO3-含量的增加可以提高反硝化速率[39],土壤有机质可以为反硝化微生物提供能量和良好的反应条件,进而有利于反硝化过程的发生[40-41]。本研究结果亦表明,土壤反硝化基因拷贝数方面是处理1/4M+3/4R和CK2表现较好,而通过土壤理化性质分析[42],处理1/4M+3/4R能提高土壤全氮、碱解氮和有机质含量,进一步影响了土壤反硝化基因进程,与前人结果一致。同时,本研究中土壤基因丰度远远高于AOAAOB基因丰度,说明反硝化作用对N2O的贡献更大。另外,本研究发现,晚稻土壤中的AOA、AOB、基因丰度比早稻要高,说明紫云英与油菜混播能提高土壤硝化和反硝化基因丰度。由于本研究试验时间较短,导致有不少指标差异不显著,需继续定位试验加以探究。在今后的试验中可增加与N2O排放相关的影响指标如土壤含水量、Eh等,进行更深入研究。
4 结论
冬作季N2O排放主要集中在冬季绿肥的盛花期,早晚稻季N2O排放主要集中在水稻的分蘖、灌浆和成熟期。总体来看,稻田N2O排放主要集中在水稻季,早稻季的N2O排放量要低于晚稻季。连续两年的N2O周年累积排放量均是单播处理比混播处理要低,而混播处理中3/4M+1/4R的N2O周年累积排放量最低。反硝化作用对N2O排放贡献更大。紫云英与油菜混播提高了土壤AOA和AOB基因丰度,AOA基因为优势功能基因,而且提高了土壤基因丰度,基因占主导地位。综合来看,“3/4紫云英+1/4油菜-早稻-晚稻”种植模式在保证高产和高肥力的情况下,N2O排放量相对较低,是较理想的种植模式。
[1] 黄国勤. 长江经济带稻田耕作制度绿色发展探讨. 中国生态农业学报(中英文), 2020, 28(1): 1-7. doi:10.13930/j.cnki.cjea.190500.
HUANG G Q. Green development of paddy field farming systems in the Yangtze River Economic Belt. Chinese Journal of Eco-Agriculture, 2020, 28(1): 1-7. doi:10.13930/j.cnki.cjea.190500. (in Chinese)
[2] 杨滨娟, 黄国勤. 双季稻田冬种紫云英“双减双增”绿色高效循环农业模式. 江苏农业科学, 2018, 46(16): 51-56.
YANG B J, HUANG G Q. A green and efficiency recycling agriculture mode of milk vetch with “double-reduction and double- increment”. Jiangsu Agricultural Sciences, 2018, 46(16): 51-56. (in Chinese)
[3] 张永亮, 潘东, 吴明浩. 苜蓿+禾草混播方式对二龄混播草地牧草产量的影响. 草业科学, 2018, 35(9): 2210-2219.
ZHANG Y L, PAN D, WU M H. Effect of mixed alfalfa and grass cultivation on the forage yield of grasslands after two-year mixed sowing. Pratacultural Science, 2018, 35(9): 2210-2219. (in Chinese)
[4] 王家琴, 王仕玥, 陆家环, 杨绍华. 非豆科绿肥与豆科绿肥混播对绿肥鲜草生长的影响. 现代农业科技, 2010(12): 250-253.
WANG J Q, WANG S Y, LU J H, YANG S H. Effects of mixed planting of non-leguminous green manure and leguminous green manure on the growth of green manure. Modern Agricultural Science and Technology, 2010(12): 250-253. (in Chinese)
[5] 杨滨娟, 孙松, 陈洪俊, 黄国勤. 稻田水旱轮作系统的能值分析和可持续评价. 生态科学, 2017, 36(1): 123-131.
YANG B J, SUN S, CHEN H J, HUANG G Q. Research on emergy analysis and sustainability evaluation under paddy-upland rotation systems. Ecological Science, 2017, 36(1): 123-131. (in Chinese)
[6] 张玉铭, 胡春胜, 张佳宝, 董文旭, 王玉英, 宋利娜. 农田土壤主要温室气体(CO2、CH4、N2O)的源/汇强度及其温室效应研究进展. 中国生态农业学报, 2011, 19(4): 966-975.
ZHANG Y M, HU C S, ZHANG J B, DONG W X, WANG Y Y, SONG L N. Research advances on source/sink intensities and greenhouse effects of CO2, CH4and N2O in agricultural soils. Chinese Journal of Eco-Agriculture, 2011, 19(4): 966-975. (in Chinese)
[7] IPCC. Intergovernmental Panel on Climate Change 2007: The Physical Science Basis. Cambridge University Press, 2007.
[8] ZOU J W, HUANG Y, ZHENG X H, WANG Y S. Quantifying direct N2O emissions in paddy fields during rice growing season in mainland China: Dependence on water regime. Atmospheric Environment, 2007, 41: 8030-8042.
[9] YOSHIDA M, ISHII S, FUJII D, OTSUKA S, SENOO K. Identification of active denitrifiers in rice paddy soil by DNA- and RNA-based analyses. Microbes and Environments, 2012, 27: 456-461.
[10] OLSSON P A, HANSSON M C, BURLEIGH S H. Effect of P availability on temporal dynamics of carbon allocation and glomus intraradices high-affinity P transporter gene induction in arbuscular mycorrhiza. Applied and Environmental Microbiology, 2006, 72: 4115-4120.
[11] TEJADA M, GONZALEZ J L, GARCIA-MARTINEZ A M, PARRADO J. Effects of different green manures on soil biological properties and maize yield. Bioresource Technology, 2008, 99(6): 1758-1767.
[12] 李隆. 间套作强化农田生态系统服务功能的研究进展与应用展望. 中国生态农业学报, 2016, 24(4): 403-415. doi:10.13930/j.cnki.cjea. 160061.
LI L. Intercropping enhances agroecosystem services and functioning: current knowledge and perspectives. Chinese Journal of Eco- Agriculture, 2016, 24(4): 403-415. doi:10.13930/j.cnki.cjea.160061. (in Chinese)
[13] 王艳秋, 高嵩涓, 曹卫东, 李景环, 聂军, 徐昌旭, 白金顺, 曾闹华, 周国朋. 多年冬种紫云英对两种典型双季稻田土壤肥力及硝化特征的影响. 草业学报, 2017, 26(2): 180-189. doi:10.11686/cyxb2016281.
WANG Y Q, GAO S J, CAO W D, LI J H, NIE J, XU C X, BAI J S, ZENG N H, ZHOU G P. Fertility and nitrification characteristics of two typical paddy soils after application of milk vetch () for 8 years. Acta Prataculturae Sinica, 2017, 26(2): 180-189. doi:10.11686/cyxb2016281. (in Chinese)
[14] 高嵩涓. 冬绿肥—水稻模式下的土壤微生物特征及硝化作用调控机制[D]. 北京: 中国农业大学, 2018.
GAO S J. Soil microbial community and the regulation mechanisms of nitrification in winter green manure-rice cropping system[D]. Beijing: China Agricultural University, 2018. (in Chinese)
[15] LIU G, MA J, YANG Y T, YU H Y, ZHANG G B, XU H. Effects of straw incorporation methods on nitrous oxide and methane emissions from a wheat-rice rotation system. Pedosphere, 2019, 29(2): 204-215. doi:10.1016/S1002-0160(17)60410-7.
[16] CHEN Z M, DING W X, XU Y H, MÜLLER C, YU H Y, FAN J L. Increased N2O emissions during soil drying after waterlogging and spring thaw in a record wet year. Soil Biology and Biochemistry, 2016, 101: 152-164. doi:10.1016/j.soilbio.2016.07.016.
[17] FRANCIS C A, ROBERTS K J, BEMAN J M, SANTORO A E, OAKLEY B B. Ubiquity and diversity of ammonia-oxidizing Archaea in water columns and sediments of the ocean. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(41): 14683-14688. doi:10.1073/pnas.0506625102.
[18] ROTTHAUWE J H, WITZEL K P, LIESACK W. The ammonia monooxygenase structural geneas a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Applied and Environmental Microbiology, 1997, 63(12): 4704-4712. doi:10.1128/aem.63.12.4704-4712.1997.
[19] THROBÄCK I N, ENWALL K, JARVIS Å, HALLIN S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiology Ecology, 2004, 49(3): 401-417. doi:10.1016/j.femsec.2004.04.011.
[20] BRAKER G, FESEFELDT A, WITZEL K P. Development of PCR primer systems for amplification of nitrite reductase genes (nirK and nirS) to detect denitrifying bacteria in environmental samples. Applied and Environmental Microbiology, 1998, 64(10): 3769-3775. doi:10. 1128/AEM.64.10.3769-3775.1998.
[21] HENRY S, BRU D, STRES B, HALLET S, PHILIPPOT L. Quantitative detection of the nosZ gene, encoding nitrous oxide reductase, and comparison of the abundances of 16S rRNA, narG, nirK, and nosZ genes in soils. Applied and Environmental Microbiology, 2006, 72(8): 5181-5189. doi:10.1128/AEM.00231-06.
[22] 王晓萌, 孙羽, 王麒, 宋秋来, 曾宪楠, 冯延江. 稻田温室气体排放与减排研究进展. 黑龙江农业科学, 2018(7): 149-154. doi:10.11942/j.issn1002-2767.2018.07.0149.
WANG X M, SUN Y, WANG Q, SONG Q L, ZENG X N, FENG Y J. Research progress on greenhouse gas emission and emission reduction in rice fields. Heilongjiang Agricultural Sciences, 2018(7): 149-154. doi:10.11942/j.issn1002-2767.2018.07.0149. (in Chinese)
[23] 李虎, 邱建军, 王立刚, 任天志. 中国农田主要温室气体排放特征与控制技术. 生态环境学报, 2012, 21(1): 159-165. doi:10.16258/ j.cnki.1674-5906.2012.01.025.
LI H, QIU J J, WANG L G, REN T Z. The characterization of greenhouse gases fluxes from croplands of China and mitigation technologies. Ecology and Environmental Sciences, 2012, 21(1): 159-165. doi:10.16258/j.cnki.1674-5906.2012.01.025. (in Chinese)
[24] 唐海明, 肖小平, 孙继民, 汤文光, 汪柯, 李微艳, 杨光立. 种植不同冬季作物对稻田甲烷、氧化亚氮排放和土壤微生物的影响. 生态环境学报, 2014, 23(5): 736-742. doi:10.16258/j.cnki.1674-5906.2014. 05.022.
TANG H M, XIAO X P, SUN J M, TANG W G, WANG K, LI W Y, YANG G L. Effects of different winter covering crops cultivation on methane and nitrous oxide emission fluxes and soil microorganism in double-cropping paddy field. Ecology and Environmental Sciences, 2014, 23(5): 736-742. doi:10.16258/j.cnki.1674-5906.2014.05.022. (in Chinese)
[25] 胡玉麟, 汤水荣, 陶凯, 何秋香, 田伟, 秦兴华, 伍延正, 孟磊. 优化施肥模式对我国热带地区水稻-豇豆轮作系统N2O和CH4排放的影响. 环境科学, 2019, 40(11): 5182-5190. doi:10.13227/j.hjkx. 201905095.
HU Y L, TANG S R, TAO K, HE Q X, TIAN W, QIN X H, WU Y Z, MENG L. Effects of optimizing fertilization on N2O and CH4emissions in a paddy-cowpea rotation system in the tropical region of China. Environmental Science, 2019, 40(11): 5182-5190. doi:10. 13227/j.hjkx.201905095. (in Chinese)
[26] 王肖娟, 王永强, 赵双玲, 李丽, 苏天潮, 刘小武. 不同灌溉方式及施肥量对稻田土壤N2O排放的影响. 大麦与谷类科学, 2018, 35(3): 1-4, 21. doi:10.14069/j.cnki.32-1769/s.2018.03.001.
WANG X J, WANG Y Q, ZHAO S L, LI L, SU T C, LIU X W. Effects of drip irrigation and flood irrigation under different application rates of nitrogen fertilizer on N2O emission in rice field. Barley and Cereal Sciences, 2018, 35(3): 1-4, 21. doi:10.14069/j.cnki.32-1769/s.2018. 03.001. (in Chinese)
[27] 张岳芳, 周炜, 王子臣, 陈留根, 朱普平, 盛婧, 郑建初. 氮肥施用方式对油菜生长季氧化亚氮排放的影响. 农业环境科学学报, 2013, 32(8): 1690-1696.
ZHANG Y F, ZHOU W, WANG Z C, CHEN L G, ZHU P P, SHENG J, ZHENG J C. Effects of nitrogen fertilizer application modes on nitrous oxide emissions during growing season of oilseed rape(). Journal of Agro-Environment Science, 2013, 32(8): 1690-1696. (in Chinese)
[28] 田伟, 伍延正, 汤水荣, 胡玉麟, 赖倩倩, 文冬妮, 孟磊, 吴川德. 不同施肥模式对热区晚稻水田CH4和N2O排放的影响. 环境科学, 2019, 40(5): 2426-2434. doi:10.13227/j.hjkx.201808221.
TIAN W, WU Y Z, TANG S R, HU Y L, LAI Q Q, WEN D N, MENG L, WU C D. Effects of different fertilization modes on greenhouse gas emission characteristics of paddy fields in hot areas. Environmental Science, 2019, 40(5): 2426-2434. doi:10.13227/j.hjkx.201808221. (in Chinese)
[29] GOLDBERG S D, GEBAUER G. N2O and NO fluxes between a Norway spruce forest soil and atmosphere as affected by prolonged summer drought. Soil Biology and Biochemistry, 2009, 41(9): 1986-1995. doi:10.1016/j.soilbio.2009.07.001.
[30] 常单娜, 刘春增, 李本银, 吕玉虎, 潘兹亮, 高嵩涓, 曹卫东. 翻压紫云英对稻田土壤还原物质变化特征及温室气体排放的影响. 草业学报, 2018, 27(12): 133-144. doi:10.11686/cyxb2018029.
CHANG D N, LIU C Z, LI B Y, LÜ Y H, PAN Z L, GAO S J, CAO W D. Effects of incorporating Chinese milk vetch on reductive material characteristics and greenhouse gas emissions in paddy soil. Acta Prataculturae Sinica, 2018, 27(12): 133-144. doi:10.11686/cyxb2018029. (in Chinese)
[31] 聂江文, 王幼娟, 吴邦魁, 刘章勇, 朱波. 紫云英还田对早稻直播稻田温室气体排放的影响. 农业环境科学学报, 2018, 37(10): 2334-2341. doi:10.11654/jaes.2018-0195.
NIE J W, WANG Y J, WU B K, LIU Z Y, ZHU B. Effect of Chinese milk vetch incorporation on greenhouse gas emissions from early-rice direct-seeding paddy fields. Journal of Agro-Environment Science, 2018, 37(10): 2334-2341. doi:10.11654/jaes.2018-0195. (in Chinese)
[32] 唐海明, 肖小平, 帅细强, 汤文光, 林叶春, 汤海涛, 杨光立. 双季稻田种植不同冬季作物对甲烷和氧化亚氮排放的影响. 生态学报, 2012, 32(5): 1481-1489. doi:10.5846/stxb201101230115.
TANG H M, XIAO X P, SHUAI X Q, TANG W G, LIN Y C, TANG H T, YANG G L. Effects of different winter covering crops cultivation on methane(CH4)and nitrous oxide(N2O)emission fluxes from double-cropping paddy field. Acta Ecologica Sinica, 2012, 32(5): 1481-1489. doi:10.5846/stxb201101230115. (in Chinese)
[33] 魏云霞. 紫云英与油菜、黑麦草混播种植和利用效应研究[D]. 武汉: 华中农业大学, 2013.
WEI Y X. Study on cultivation and utilization of the mixture cropping of Chinese milk vetch with rape and ryegrass[D]. Wuhan: Huazhong Agricultural University, 2013. (in Chinese)
[34] 朱波, 易丽霞, 胡跃高, 曾昭海, 唐海明, 肖小平, 杨光立. 黑麦草鲜草翻压还田对双季稻CH4与N2O排放的影响. 农业工程学报, 2011, 27(12): 241-245. doi:10.3969/j.issn.1002-6819.2011.12.045.
ZHU B, YI L X, HU Y G, ZENG Z H, TANG H M, XIAO X P, YANG G L. Effects of ryegrass incorporation on CH4and N2O emission from double rice paddy soil. Transactions of the Chinese Society of Agricultural Engineering, 2011, 27(12): 241-245. doi:10.3969/j.issn. 1002-6819.2011.12.045. (in Chinese)
[35] 纪钦阳, 张璟钰, 王维奇. 施肥量对福州平原稻田CH4和N2O通量的影响. 亚热带农业研究, 2015, 11(4): 246-253. doi:10.13321/j.cnki. subtrop.agric.res.2015.04.006.
JI Q Y, ZHANG J Y, WANG W Q. Effect of fertilization rate on CH4and N2O emission in paddy fields in Fuzhou plain. Subtropical Agriculture Research, 2015, 11(4): 246-253. doi:10.13321/j.cnki. subtrop.agric.res.2015.04.006. (in Chinese)
[36] 钟川, 杨滨娟, 张鹏, 李萍, 黄国勤. 基于冬种不同作物的水旱轮作模式对水稻产量及稻田CH4、N2O排放的影响. 核农学报, 2019, 33(2): 379-388. doi:10.11869/j.issn.100-8551.2019.02.0379.
ZHONG C, YANG B J, ZHANG P, LI P, HUANG G Q. Effect of paddy-upland rotation with different winter corps on rice yield and CH4and N2O emissions in paddy fields. Journal of Nuclear Agricultural Sciences, 2019, 33(2): 379-388. doi:10.11869/j.issn. 100-8551.2019.02.0379. (in Chinese)
[37] 秦红灵, 陈安磊, 盛荣, 张文钊, 邢肖毅, 魏文学. 稻田生态系统氧化亚氮(N2O)排放微生物调控机制研究进展及展望. 农业现代化研究, 2018, 39(6): 922-929. doi:10.13872/j.1000-0275.2018.0092.
QIN H L, CHEN A L, SHENG R, ZHANG W Z, XING X Y, WEI W X. A review on the microbial regulation mechanism of N2O production and emission of rice paddy ecosystems. Research of Agricultural Modernization, 2018, 39(6): 922-929. doi:10.13872/j. 1000-0275.2018.0092. (in Chinese)
[38] 李双双, 陈晨, 段鹏鹏, 许欣, 熊正琴. 生物质炭对酸性菜地土壤N2O排放及相关功能基因丰度的影响. 植物营养与肥料学报, 2018, 24(2): 414-423. doi:10.11674/zwyf.17272.
LI S S, CHEN C, DUAN P P, XU X, XIONG Z Q. Effects of biochar application on N2O emissions and abundance of nitrogen related functional genes in an acidic vegetable soil. Plant Nutrition and Fertilizer Science, 2018, 24(2): 414-423. doi:10.11674/zwyf.17272. (in Chinese)
[39] 谭立山. 农业土壤N2O产生途径及其影响因素研究进展. 亚热带农业研究, 2017, 13(3): 196-204. doi:10.13321/j.cnki.subtrop.agric.res. 2017.03.011.
TAN L S. Advances in N2O generation pathway in agricultural soils and major influencing factors. Subtropical Agriculture Research, 2017, 13(3): 196-204. doi:10.13321/j.cnki.subtrop.agric.res.2017.03.011. (in Chinese)
[40] LIMMER A W, STEELE K W. Denitrification potentials: measurement of seasonal variation using a short-term anaerobic incubation technique. Soil Biology and Biochemistry, 1982, 14(3): 179-184. doi:10.1016/ 0038-0717(82)90020-7.
[41] 李萍. 紫云英与油菜混播对稻田土壤肥力、N2O排放及相关功能基因丰度的影响[D]. 南昌: 江西农业大学, 2020.
LI P. Effects of the mixture of Chinese milk vetch and rape on soil fertility, N2O emission and abundance of related functional genes in paddy fields[D]. Nanchang: Jiangxi Agricultural University, 2020. (in Chinese)
[42] 李勇先. 稻田土壤中氧化亚氮的释放机制及控制[D]. 杭州: 浙江大学, 2003.
LI Y X. Mechanism and control of N2O emission from paddy soils[D]. Hangzhou: Zhejiang University, 2003. (in Chinese)
Effects of the Mixted-cropping of Chinese Milk Vetch and Rape on Soil Nitrous Oxide Emission and Abundance of Related Functional Genes in Paddy Fields
YANG BinJuan, LI Ping, HU QiLiang, HUANG GuoQin
College of Agronomy, Jiangxi Agricultural University/Research Center on Ecological Science Jiangxi Agricultural University, Nanchang 330045
【】The aim of this study was to determine how a green manure mixted-cropping affected nitrous oxide (N2O) emissions from soil and the abundance of related functional genes in paddy field soil. The ultimate aim of this research was to identify a green manure mixted-cropping that had high and stable yields and reduces emissions, and was suitable for cultivation in the double-rice areas of southern China.【】The effects of mixted-cropping of Chinese milk vetch and rape at different ratios (single Chinese milk vetch, single rape, 3/4 Chinese milk vetch +1/4 rape, 1/2 Chinese milk vetch +1/2 rape, and 1/4 Chinese milk vetch +3/4 rape) on N2O emissions and the abundance of related functional genes in paddy field soil were determined and compared.【】(1) The crop species affected N2O emissions. The N2O emissions were similar among the different treatments. Among them the N2O emissions from winter crop soil were mainly concentrated at its flowering stage, and those during the rice growing period were concentrated at the tillering, filling, and maturity stages of early and late rice crops. On the whole, the N2O emissions from paddy fields were lower N2O in the early rice season than that in the late rice season. The overall N2O emissions flux in the paddy fields was higher in 2018 than that in 2019. (2) There was no significant difference in N2O cumulative emission of early rice under different treatments. The cumulative N2O emissions from a single seeding treatment were lower than those from mixed treatments over two consecutive years. The annual cumulative N2O emissions were lowest under CK1 in 2019. Among all the mixed treatments, the 3/4 Chinese milk vetch +1/4 rape treatment had the lowest annual cumulative N2O emissions. (3) In general, the copy number of-AOA and-AOB genes was higher in the soil of late rice than that in the soil of early rice, and the copy number of-AOA was up to two orders of magnitude higher in late rice soil than in early rice soil. The copy numbers of,andwere higher in the soil of late rice crops than that in the soil of early rice crops, and the copy number ofwas up to two orders of magnitude higher than those ofandin late rice soil. The mixted-cropping of Chinese milk vetch and rape increased the abundance-AOA and-AOB genes in soil, and-AOA was the dominant functional gene. It also increased the abundance of,andgenes in soil, andgene was the dominant gene. The copy number of genes related to denitrification was one to three orders of magnitude higher than the copy number of genes related to nitrification, indicating that denitrification made a larger contribution to N2O emissions.【】Overall, the 3/4 Chinese milk vetch + 1/4 rape-early rice-late rice was the best mixed planting system, because it had the best combination of low N2O emissions as well as high yield and high fertility.
Chinese milk vetch; rape; green manure mixted-cropping; nitrous oxide; functional gene; paddy fields

10.3864/j.issn.0578-1752.2022.04.010
2021-01-21;
2021-05-14
国家自然科学基金项目(32160528)、国家重点研发计划课题(2016YFD0300208)、国家自然科学基金(41661070)
杨滨娟,Tel:0791-83828143;E-mail:yangbinjuan@jxau.edu.cn。通信作者黄国勤,Tel:0791-83828143;E-mail: hgqjxes@sina.com
(责任编辑 李云霞)
