鸟类物种组合模式的稳定性
——以珠海淇澳自然保护区红树林鸟类群落变化为例
2022-04-16叶锦玉虞皓琦廖宝文游奕来刘新科林寿明杨道德胡慧建
叶锦玉 ,虞皓琦,廖宝文,游奕来,刘新科,林寿明,杨道德,胡慧建*
1. 中南林业科技大学野生动植物保护研究所,湖南 长沙 410004;
2. 广东省科学院动物研究所/广东省动物保护与资源利用重点实验室/广东省野生动物保护与利用公共实验室,广东 广州 510260;3. 中国林业科学研究院热带林业研究所,广东 广州 510520;4. 广东珠海淇澳-担杆岛省级自然保护区管理处,广东 珠海 519000;5 广东省林业调查规划院,广东 广州 510520
群落构建规则(community assembly rules)是生态学研究的核心和基础问题,也是恢复生态学的重要研究领域(Brown et al.,1991;Temperton et al.,2004;Götzenberger et al.,2012)。群落的物种组合模式从不同侧面反映了群落构建的特点,是其重要研究内容之一(Brown et al.,1991)。许多研究表明,动物群落的物种组合具有多种相对固定的模式,如种面积效应(Brown et al.,1991;Connor et al.,2001)、物种个体大小频次偏正态分布(Brown et al.,1991)、物种多度频次偏正态分布(Brown,1995),以及种-属数量间强相关关系(Blackburn et al.,1999;Gaston,2000;胡慧建等,2003)和属含物种的频次分布可乘幂模型拟合(冯永军等,2006;郑孜文等,2008;连军豪等,2011)等。这些模式的存在可能意味着群落的构建具有稳定性。
植被群落是动物栖息地的重要组成部分,本地植被群落的改变会导致动物栖息地和食物变化,从而导致物种发生重组(Davidsdottir et al.,2016;Lindbladh et al.,2019)。若物种重组后其物种组合模式仍与重组前一致,则说明群落的物种组合模式具有可重复性,这对解释物种共存和物种多样性的维持有重要作用。目前植被变化对动物群落产生影响的相关研究主要集中在物种数量(江海声等,1998)、物种组成(Isacch et al.,2005;Fontaine et al.,2009;Nkwabi et al.,2010;张国钢等,2008)、多样性指数(Fontaine et al.,2009;Nkwabi et al.,2010;Curado et al.,2013)等,而对群落的种属关系、个体大小等物种组合模式变化的影响研究仍存在空白。研究植被变化前后动物群落是否具有相同或类似的物种组合模式,或存在其他明显的群落构建规则,是验证动物群落构建可重复性的重要依据,也可对动物群落的保护与修复提供有力的理论支撑。
广东珠海淇澳-担杆岛省级自然保护区的淇澳岛片区曾是原生红树林重要的分布地,但由于大量围海造田、围垦养殖以及桥梁、码头建设等,逐渐演变为以互花米草(Spartina alterniflora)为优势的植被群落。为解决红树林面积萎缩和互花米草入侵问题,自 2001开始在淇澳岛试种以无瓣海桑(Sonneratia apetala)为主的红树林植被,如今,红树林已替代互花米草成为该区域的优势植被(廖宝文等,2008)。本研究利用淇澳岛红树林恢复前后的2次鸟类调查数据(恢复前于2001年3月—12月,恢复后于2015年10月—2016年6月),分析区域植被发生变化后鸟类物种及其组合模式的变化,探讨鸟类群落构建中物种的组合模式,为动物群落的恢复提供新视角。
1 研究方法
1.1 研究地点
广东珠海淇澳-担杆岛省级自然保护区成立于2004年,由珠海市的淇澳岛和担杆岛两个片区组成,总面积7373.77 hm2。该区域位于珠江口,属海湾地貌,主要有丘陵、浅海沉积(滩涂)的地貌等类型;地处北回归线附近,南亚热带海洋性季风气候,年均气温22—23 ℃之间,年平均降雨量1975.1 mm。本研究于淇澳岛内,范围以植被覆盖最边缘处为界,主要位于海堤两侧中潮位以上,面积约 903 hm2, 四至边界为 113°36′40″— 113°39′15″E ,22°23′40″—22°27′38″N。
上世纪 80年代初,该区域内有红树林面积112.2 hm2,树种以秋茄(Kandelia obovata)和桐花树(Aegiceras carniculatum)为主。1990年以后,由于围海造地、人为砍伐和互花米草入侵等危害,红树林面积仅存32 hm2。保护区于1999年和2000年尝试小规模种植以无瓣海桑为主的红树林,面积约 1 hm2;2002年后开始大量种植(2001年无种植),最大面积达500 hm2,占淇澳岛片区总面积的9.80%,约占潮间带面积的50.00%(图1)。现红树林生态系统自 2008年开始趋于稳定,有红树植物32种,真红树20种,半红树12种,其中栽培种有21种(廖宝文等,2008)。

图1 珠海淇澳岛红树林及样线样点分布示意图Figure 1 Location of mangrove and transect in Qi’ao Island, Zhuhai, Guangdong
2 调查方法
第1次调查在红树林大面积恢复前(下简称恢复前),时间为2001年3月31日—4月1日、7月8—9日、10月12—13日和12月26—27日,此时调查范围内植被以互花米草为主。第2次调查在红树林恢复后(下简称恢复后),时间为2015年10月29—30日和12月24—25日、2016年3月29—30日和6月19—20日,此时的调查范围内已形成以红树林为主的植被群落(廖宝文等,2008)。
2次鸟类调查的样线和样点相同,调查选择天气良好情况下进行,每日调查时间为07:00—11:00、15:00—17:00,中午鸟类活跃性较低,不进行调查。分别在该片区西边海堤和东边栈道各有1条样线,长度分别为2.5 km和4.5 km,样线单侧宽度为100 m;东边海堤高处有1个样点,半径宽200 m(图1)。鸟类鉴定参照约翰·马敬能等(2000)所著《中国鸟类野外手册》,分类系统、区系和居留型等参照《中国鸟类分类与分布名录 (第2版)》(郑光美,2011)。
3 数据处理
3.1 物种累积曲线
采用物种累积曲线(species accumulative curve)判断2次鸟类调查的充分性(李巧,2011),使用R语言vegan程序包计算物种累计曲线。如果累积曲线为直线上升,表明抽样量不足,需要增加抽样量;如果曲线在急剧上升后变为渐近线或增速趋势放缓,则说明抽样充分。恢复后调查记录了物种个体数,因此直接计算物种累积曲线,并判断其抽样充分性。
由于恢复前调查记录的为数量等级,即“+”<5只,“++”为5—99只,“+++”为100—500只,“++++”为>500只,因此用数量等级的最小值计算物种累积曲线,即物种数量等级为“+”则该物种数量换算为1只,“++”换算为5只,“+++”换算为100只,“++++”换算为500只。换算后计算得出的物种累积曲线在急速上升后变为渐近线(图2),因此认为恢复后调查抽样充分。

图2 2次调查鸟类物种累积曲线Figure 2 Species accumulation curve of birds survey before and after mangrove restoration
3.2 物种组成
对2次调查的鸟类名录进行分析,对比物种丰度、区系、居留型、季节差异及保护物种等。统计2次调查全部、共有物种以及单次调查记录的区系(东洋界、古北界和广布种)、居留型(留鸟、夏候鸟和冬候鸟)、季节、生态类型(涉禽、游禽、猛禽、陆禽、攀禽和鸣禽)的物种数,用卡方检验判断其差异的显著性。本节及下节均采用统计软件IBM SPSS Statistics 25对数据进行分析。
3.3 种属结构关系
动物的属和种在数量上具有显著的相关性(Connor et al.,2001),属含物种数分布频次随物种数增加而减少,暨1种/属最多,2种/属次之,3种/属更少……(Brown,1995),且显示出明显的规律性,可用乘幂模型拟合(冯永军等,2006)。故本研究对比以上两种关系在2次调查中的差异,以观察其中分类结构的变化。
3.3.1 种-属数量关系
分别统计2次调查按春至冬季节顺序累计的物种数和属数,对种-属数量进行相关关系分析。采用K-S方法对数据进行分布型检验,凡符合正态分布的数据,采用Pearson线性相关分析,否则采用Spearman秩相关分析。采用协方差分析判断恢复前后种-属数量的回归直线是否具有一致性。
3.3.2 属含物种数的频次分布
分别统计2次调查具有相同物种数的属的频次,暨统计1种/属、2种/属、3种/属……出现的数量,并进行K-S方法对数据进行分布型检验,发现不符合正态分布。故根据其数据分布规律,采用乘幂模型进行拟合,再进行ln转换为线性模型,采用协方差分析该模型的一致性,以判断恢复前后属含物种数的乘幂模型的一致性。
3.4 体长结构
采用马敬能等著《中国鸟类野外手册》(约翰·马敬能等,2000)中鸟类物种体长数据对 2次调查名录中的鸟类体长进行统计,并进一步以10 cm为间隔分别统计2次调查名录中鸟类物种体长的频次分布。2次调查的鸟类全部和单次调查记录物种的体长频次分布的差异均采用卡方检验进行分析。
4 结果
4.1 物种累积曲线
恢复前调查的鸟类物种数量等级数据通过数量等级的最小值换算后,物种累积曲线在快速上升后变为一渐近线;恢复后调查的物种累积曲线亦为先快速上升后增速放缓,说明2次调查取样均较充分(图2)。
4.2 物种组成
2次调查共记录到鸟类99种,隶属于13目35科70属。其中,恢复前调查共记录鸟类11目29科58属78种;恢复后调查共记录鸟类12目33科57属80种,2次调查的鸟类物种丰度基本一致。2次调查共记录鸟类有东洋界 44种(占总数的44.44%),古北界31种(占31.31%),广布种24种(占24.24%);留鸟51种(占52.04%),冬候鸟37种(占37.76%),夏候鸟10种(占10.20%)(表1);共有鸟类物种有59种(表2),分别占2次调查的76.62%和73.75%,主要为涉禽类和鸣禽类(附表1)。

表1 红树林恢复前后鸟类区系和居留型Table 1 Fauna and residence types of birds survey before and after mangrove restoration

表2 红树林恢复前后各季节鸟类物种数Table 2 Number of bird species in different seasons before and after mangrove restoration
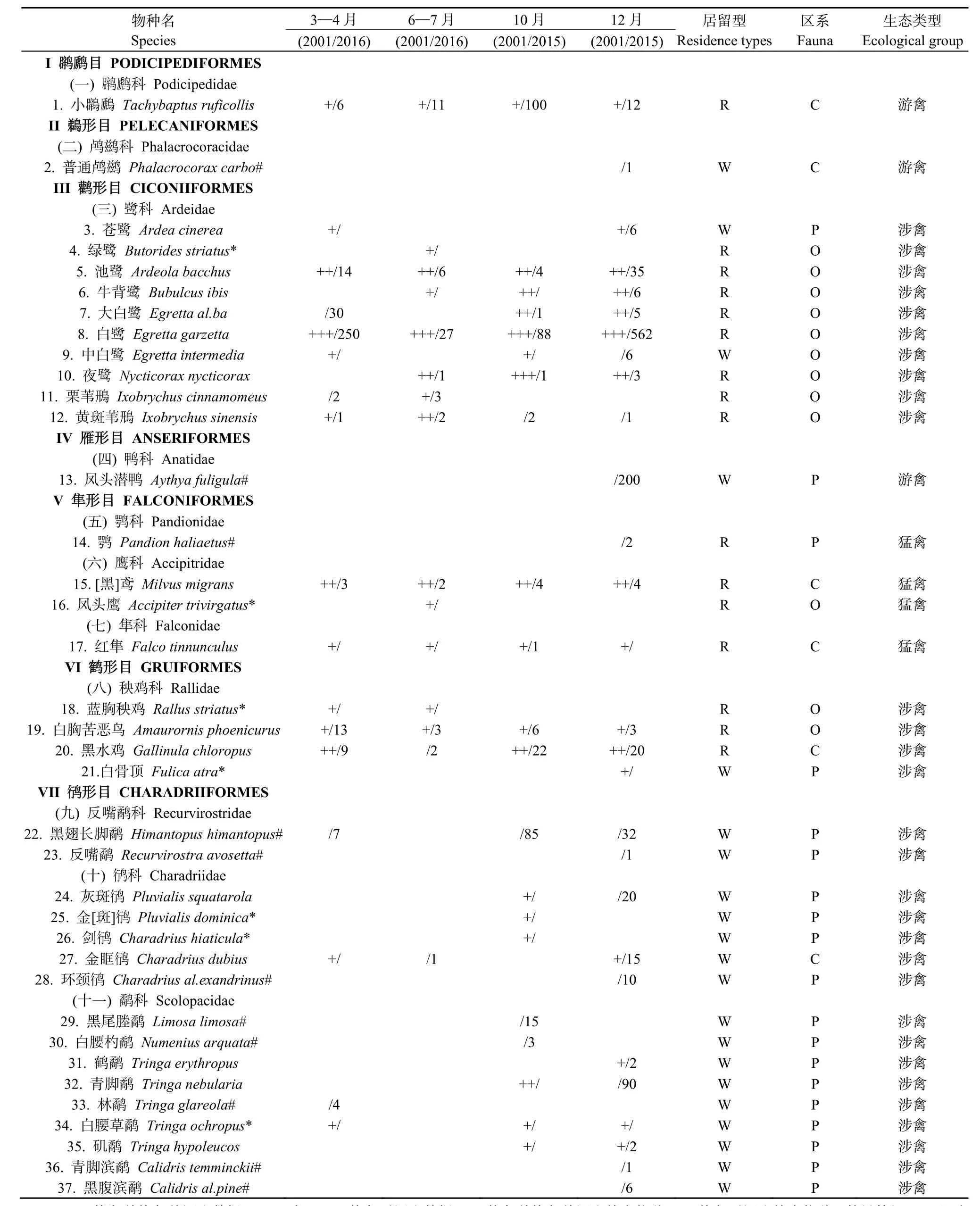
表1 红树林恢复前恢复前和恢复后不同季度鸟类种类与数量变化组成Appenddix 1 Variety and form of bird species and richness in each season before and after vegetation change

续附表1 红树林恢复前恢复前和恢复后不同季度鸟类种类与数量变化组成Continued appenddix 1 Variety and form of bird species and richness in each season before and after vegetation change

续附表1 红树林恢复前恢复前和恢复后不同季度鸟类种类与数量变化组成Continued appenddix 1 Variety and form of bird species and richness in each season before and after vegetation change
恢复前特有物种有5科17属18种,占其总数的23.38 %,主要生态类型为鸣禽、涉禽和猛禽;恢复后特有物种有 6科 12属 21种,占其总数的26.25 %,主要为涉禽和鸣禽(附表1)。恢复前与恢复后在区系(P=0.657)、居留型(P=0.638)和生态类型(P=0.151)上均无显著性差异。
4.3 季节变化
在4个季节中,恢复前后的鸟类物种组成发生了部分替代,约占总数的24%,以候鸟为主。2次调查在4个季节的物种丰富度、区系及居留型上均无显著差异(表3)。

表3 红树林恢复前后各季度鸟类的区系及居留型Table 3 Fauna and inhabitation pattern in different seasons of birds surveyed before and after mangrove restoration
4.4 分类阶元结构
4.4.1 种-属数量关系
2次调查的鸟类的物种-属数量均呈正相关关系,且能用线性方程进行拟合:恢复前为y=0.5596x+15.093(r2=0.990,P=0.001);恢复后为y=0.5215x+15.568(r2=0.990,P=0.002)(图3),协方差分析表明 2次调查的种-属间数量的回归直线平行,具有一致性(P=0.234)。

图3 红树林恢复前后鸟类属数量(y)与物种数(x)的线性模型Figure 3 Linear model of species-genera richness of birds surveyed before and after mangrove restoration
4.4.2 属含物种数的频次分布
2次调查属含物种数的频次分布均随着每属所含物种数增加呈递减关系,能用乘幂模型进行拟合:恢复前,y=51.65x−2.7(r2=0.984);恢复后,y=48.47x−2.49(r2=0.937)(图4a),协方差分析表明2次调查的属含物种数的频次分布的乘幂拟合曲线平行且具有一致性(P=0.688)。2次调查共有种的属含物种数频次分布亦可用乘幂模型进行拟合,单次调查记录物种的均为单种属占比较高(图4b)。

图4 红树林恢复前后鸟类属含物种数的频次分布Figure 4 Frequency distribution of number of species in genus before and after mangrove restoration
4.5 体长频次分布
2次调查的体长频次分布有相似,体长均以11—20 cm的最多,分别有27种和26种;体长为1—10 cm的鸟类物种数量均仅有4种;体长大于21 cm的体长组的物种数逐渐减少(图5a)。2次调查的体长分布均呈正偏态分布,卡方检验结果表明2次调查鸟类各体长组的频次分布无显著性差异(P=0.992)。共有种和特有种的体长频次分布皆的无显著差异(P=0.957)(图5b)。

图5 红树林恢复前恢复前后鸟类体长频次分布Figure 5 Frequency distribution of body size of species surveyed before and after mangrove restoration
5 讨论
5.1 植被变化对鸟类群落的影响
2次调查共发现鸟类99种,红树林恢复前记录到78种,恢复后调查到80种;恢复前特有的有19种,恢复后则有21种,约24%的物种发生变化。植物不仅是鸟类所有食物的最终来源,还为鸟类提供了巢址、巢材、栖木和隐蔽等各种必需的生存条件(郑光美,2012),某植被类型或某几种植被还可能与某些鸟类形成密切依存关系(Nkwabi et al.,2010;郑光美,2012)。红树林恢复后,棕扇尾莺(Cisticola juncidis)、红尾水鸲(Rhyacornis fuliginosus)、剑鸻(Charadrius hiaticula) 和金鸻 (Pluvialis dominica)等在水边灌丛或空地上觅食的物种消失,而蓝矶鸫(Monticola solitaries)、灰背鸫(Turdus hortulorum)和叉尾太阳鸟(Aethopyga christinae)等在林间活动的鸟类,及青脚滨鹬(Calidris temminckii)、黑翅长脚鹬(Himantopus himantopus)和黑尾塍鹬(Limosa limosa)等在较深水域觅食的物种出现。这与淇澳岛开始大面积人工种植无瓣海桑后,部分生境发生了变化,滩涂区面积减少,红树林区面积增加(廖宝文等,2008)有关。
5.2 鸟类群落物种组合模式的稳定性
动物群落的物种丰度具有种-面积效应,即物种丰度和区域面积呈正相关(Arrhenius,1921;唐志尧等,2009)。由此可推断当区域面积一定时,群落中的物种丰度可能保持相对稳定。但目前少有研究对比同区域下环境变化前后的动物群落变化,并对此推断进行验证。江海声等(1998)亦发现海南南湾半岛在植被破坏前(1950年)和恢复后(1990年)记录的鸟类物种丰度基本保持一致,分别为54种和53种,但未对此现象进行分析。本研究发现淇澳岛大面积种植红树林前后鸟类物种丰度分别为78种和80种,与南湾半岛的鸟类调查结果(江海声等,1998)类似,均符合种-面积效益的推断。但混合植被生境可能是此推断成立的条件,因为专性鸟类对植被的依赖性较强,在单一植被生境下差异较大,但在混合植被生境中专性鸟类丰度差异不显著(Ma et al.,2011)。本研究区域与海南南湾半岛(江海声等,1998)均为混合多种植被生境的区域,为鸟类提供了多样化的生境,弱化了一些专性鸟类对群落的影响。因此,研究认为在区域面积不变,且为非单一植被生境时物种丰度可能保持相对稳定。
研究发现红树林恢复前恢复前后2次调查的种和属在数量上呈正相关关系,属含物种数频次关系有1属1种频次最高,1属2种显著减少,以此类推的趋势并可用乘幂曲线拟合,均与冯永军等(2006)和郑孜文等(2008)分别对中国鸟兽的研究结果相似。说明种和属不仅在大尺度上有强相关性,在小区域尺度上也有此规律。胡军华等(2008)发现长江中游不同湖泊的鱼类在 1950s、1980s和2000s 3个时期的种-属数量及属含物种数频次分布也有此规律,但未检验其拟合曲线的一致性。本研究通过协方差分析发现,2次调查的种-属数量关系和属含物种数分布频次均无显著差异(P>0.05),且南湾半岛植被破坏前和恢复后鸟类物种(江海声等,1998)的属含物种数频次分布也有一致性(图6a),说明植被变化前后鸟类群落可在种-属结构上保持一致。

图6 南湾半岛植被变化前后鸟类属含物种数频次和体长频次分布Figure 6 Frequency distribution of number of species in genus and body size before and after vegetation change in Nanwan Peninsula
动物个体大小通过异速生长关系与物种的形态、生理和行为等密切相关(Blackburn et al.,1999)。有研究表明,个体大小相近的物种间的竞争也较强,物种个体大小分布频次分布具有规律性(Stanley,1973;Kozłowski et al.,2002;Allen et al.,2006),在大尺度上多呈正偏态分布(Blackburn et al.,1999;Clauset et al.,2009)。
本研究结果表明,红树林恢复前后的鸟类体长均呈正偏态分布,与前人的研究结果一致;且协方差分析表明,物种体长频次分布无显著性差异(P=0.893)。同样对南湾半岛的鸟类体长频次分布进行分析发现,植被破坏前和恢复后的鸟类的体长频次分布相似(图6b),且卡方检验结果表明其分布无显著差异(P=0.923)。说明鸟类群落的体长频次分布具有一定规律,并在植被变化前后可保持相对稳定的结构。2次调查记录的鸟类物种在区系、居留型、季节、生态类型和体长组上的频次分布,以及各季节的不同区系、居留型的物种数均无显著差异(表3),也进一步说明红树林恢复前后鸟类群落有相似的物种组合模式。
有研究表明,植被变化后部分新进入的鸟类与消失的鸟类存在替代关系。Nkwabi et al.(2010)发现当塞伦盖蒂平原的草原由长草变为短草时,2种云雀在功能上存在替代关系。从卡方检验结果来看,2次调查中单次调查记录鸟类在体长频次分布、生态类型、区系和居留型上均无显著差异,但除斑鱼狗(Ceryle rudis)与蓝翡翠(Halcyon pileate)外,其余物种在体长组、区系、居留型和生态类型无对应关系。研究还发现2次调查共有种的属含物种数与体长频次分布均与总体的有较高相似性,说明红树林恢复前恢复前后鸟类物种的组合模式可能受其共有物种的影响较大。
目前,尚未有研究报道环境变化后物种组合模式的可重复性及其形成机制。本研究发现2次调查的鸟类群落的属含物种数频次分布、体长频次分布均符合前人在大陆和局域尺度的研究,且在植被变化前后无显著差异。从群落构建的生态位理论出发,同属物种在形态和生态上均具有较高相似性(郑光美,2012),在环境筛选下进入同一区域,又因竞争排除法则(Gause,1934),即生态上相似的两个物种在同一条件下不能共存,使得过于相似的物种发生竞争排除,从而使各物种间的相似性受到限制,即群落共存物种间的极限相似性(limiting similarity)(Wilson et al.,1995)。这可能导致了群落中单种属最多,也是其体长分化的原因(Blackburn et al.,1999)。但同时一些物种由于对环境和资源的适合度不同而导致生态位的分化(Chesson,2000),令某些属或体长组内可共存多个物种。由此可推测物种经扩散到达某一区域后,在环境筛选和属内物种的极限相似这两个反作用力的共同推动下(Webb et al.,2002),形成了现有物种组合模式。调查区域植被变化未对这些模式造成显著性影响,可能说明其影响因子在于当地较为稳定的环境因子,如光照、温度和降水等。
5.3 物种组合模式的应用前景
目前生物群落构建的研究多集中群落构建的机制中的生态位理论(包含环境过滤和生物相互作用)(Clements,1916;Gleason,1926;Hubbell,2011)和中性理论(主要为扩散机制)(Hubbell,2011),且通常认为在不同尺度上起主要作用的构建机制不同(Götzenberger et al.,2012)。相比群落构建机制,群落的物种组合模式在不同尺度上均具有类似的规律,且其研究对象为物种的分类阶元、个体大小、生态类型和区系等变量,数据容易获取,且均与物种形态和觅食、繁殖行为等生态特征密切相关,能更高效直观地反映了动物群落的构建规则,因此在目前具有更高的应用前景。
本研究发现在植被发生变化后,鸟类群落仍依照变化前的属含物种数频次分布、体长频次分布和生态类型频次分布等物种组合模式进行构建。这些物种组合模式有望作为判断鸟类群落完整性的重要依据,如通过鸟类物种数量(Fang et al.,2020;徐正春等,2019)、属含物种数频次分布及体长分布等数据的变化,判断鸟类群落的稳定性和完整性。其中属含物种数的频次分布模型在动物群落恢复上有较高的应用价值:一方面可用于评价恢复效果,可模型的符合程度来评价群落是否已稳定;一方面在群落未达到稳定的情况下,可计算未稳定群落与稳定群落模型的差值来确定缺失的相关物种。
6 结论
本文研究结果验证了:鸟类群落在当地植被变化后其物种组成发生变化,但在物种丰富度和群落物种分类结构、体长频次分布、居留型、区系组成和生态类型上均保留了原有特征。故本研究认为区域内动物群落可依照固定的物种组合模式进行构建,具有可重复性。
致谢:感谢珠海市林业局、广东珠海淇澳-担杆岛省级自然保护区巡护队给予的支持和帮助;感谢广东省科学院动物研究所的曹宏芬、刘金成等参与调查,以及胡一鸣博士对本文摘要的审阅修改。
