不同生境下濒危植物膝柄木幼树的生态适应性
2022-04-08招礼军权佳惠朱栗琼黄曈金赟
招礼军 权佳惠 朱栗琼 黄曈 金赟



摘 要:膝柄木是我國极度濒危植物,也是广西滨海过渡带天然植被的重要组成树种。为了解光因子对膝柄木天然更新的限制影响,该文对林缘、林窗、林下三种不同光照生境下膝柄木幼树的生理和生长指标的年际变化特征进行了研究。结果表明:(1)光合有效辐射不足影响了膝柄木幼树的生长。林下幼树的地径、株高和叶面积增长量显著降低,而生长于光照充足林缘生境的膝柄木幼树,其生长指标增长量最大。(2)随着光合有效辐射的减少,膝柄木幼树的超氧化物岐化酶(SOD)和过氧化氢酶(CAT)的活性、可溶性蛋白含量、相对电导率和丙二醛(MDA)含量林缘生境显著高于林下生境的幼树,游离脯氨酸含量林下生境的幼树显著高于林缘和林窗生境的幼树。(3)随着光合有效辐射的减弱,林下生境幼树的叶绿素a、叶绿素b、类胡萝卜素含量均高于林缘和林窗的膝柄木幼树,而叶绿素a/b林缘幼树较高;林缘生境的膝柄木幼树净光合速率(Pn)、蒸腾速率(Tr)在三种生境中最高;林下生境的膝柄木幼树的胞间 CO2浓度(Ci)高于其他两种生境,而水分利用效率(WUE)在2—4月林缘生境的膝柄木幼树最高,6—10月林下生境的膝柄木幼树高于其他两种生境,12月三种生境差异不显著。综上结果表明,光照因子是膝柄木幼树时期生长的限制因子之一,生长在林缘生境的幼树因接收到的光照较为充足,其生长状况要显著好于其他生境的幼树。
关键词: 生境, 光照, 膝柄木,幼树, 生态适应性
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2022)03-0501-09
Ecological adaptability of endangered plant
Bhesa robusta sapling in different habitats
ZHAO Lijun, QUAN Jiahui, ZHU Liqiong*, HUANG Tong, JIN Yun
( Forestry College, Guangxi University, Guangxi Key Laboratory of Forest Ecology and Conservation, Nanning 530004, China )
Abstract:Bhesa robusta is a critically endangered plant in China, which also is an important component of natural vegetation in the coastal ecotone of Guangxi. In order to learn about whether the light factor has the effects on the natural regeneration of B. robusta saplings or not, the interannual variation characteristics of physiological and growth indexes of B. robusta saplings under three different photosynthetic habitats of forest edge, forest gap, and understory were studied. The results were as follows: (1) The deficiency of photosynthesis active radiation affected the growth of B. robusta saplings, the growth of ground diameter, plant height, and leaf area of understory saplings decreased significantly, while the growth index of saplings growing in the forest edge with sufficient light was the largest. (2) With the decrease of photosynthesis active radiation, the contents of superoxide dismutase (SOD) and catalase (CAT), soluble protein, relative electrical conductivity, and malonaldehyde (MDA) of the B. robusta saplings under forest edge habitat were significantly higher than those in the understory habitat, and the free proline content of B. robusta sapling under the understory habitat were significantly higher than in forest edge and forest gap habitat. (3) With the decrease of photosynthesis active radiation, the photosynthetic pigment index chlorophyll a, chlorophyll b, carotenoids of B. robusta sapling were increased in the understory habitat, and the chlorophyll a/b ratio of B. robusta sapling was higher in the forest edge habitat; The net photosynthetic rate (Pn) and transpiration rate (Tr) on the photosynthesis physiological target of B. robusta saplings in forest edge habitat were higher than those in understory and forest gap habitats; The intercellular CO2 concentration (Ci) and water use efficiency (WUE) of B. robusta sapling in the understory were higher than those in forest gap and forest edge habitats. And WUE was the highest in the forest edge habitat from February to April, WUE of B. robusta saplings in the understory habitat were higher than those of the other two habitats in June to October, and there were almost no differences between the three habitats in December. All the above results indicate that the light factor is one of the limiting factors during the growth period of B. robusta sapling, and the growth of B. robusta sapling in the forest edge habitat is significantly better than that in the forest gap and understory habitats because of the abundant sunlight.
Key words: habitat, illumination, Bhesa robusta sapling, ecological adaptability
膝柄木(Bhesa robusta)是卫矛科(Celastra-ceae),卫矛亚科(Subfam. Celastroideae)的常绿乔木,主要分布在广西,其伴生种主要有豹皮樟、桃金娘等(梁盛业,1988)。膝柄木是高大的乔木,主要生长在滨海过渡带,在稳定滨海过渡带生态系统稳定和抵御台风等自然灾害的侵袭方面具有重要作用(莫竹承等,2008)。膝柄木的野外调查研究发现由于其植株的地理分布相距太远,其种子发芽率与产量低,且大多不育(安家成等,2017),通过有性繁殖增加其种群数量的方法实施难度较大(吴小巧等,2004)。国外对膝柄木的研究寥寥无几,但是国内开展了一些相关研究如对其伴生群落(莫竹承等,2013)、结构特征(徐峰等,1990)、扦插实验(莫竹承等,2008)、愈伤组织(莫竹承等,2015)及种子催芽(钟国贵等,2016)等。随着人类活动领域的扩大,许多原生环境受到了强烈干扰,生物多样性面临着巨大的威胁,且膝柄木自身天然更新困难(莫竹承等,2008),现已成为了国家濒危物种。在1999年公布的《国家重点保护野生植物名录(第一批)》中,已将膝柄木列为一级保护植物。在1997—2001年间广西进行的重点野生植物资源调查结果中显示膝柄木仅存10 株(刘演和宁世江,2002),膝柄木是我国一级保护濒危物种,是该属唯一分布在我国的特有种,现已处于极度濒危状态(黄立新,2006)。
光照是植株从种子萌发、发育、生长和完成自然更新整个生活史关键(孙清琳等,2019)。光因子能够影响植物的形态、分布和变化规律。种子在萌发以后进入幼苗期,幼苗对外界因子的响应与其所处的生境关联较强。生长于林下的幼树,由于郁闭的林冠遮挡了大部分入射的太阳光,因其接收到的光照减少(Parelle & Ducrey,2006),为适应弱光环境,则会通过改变其光合生理特性和生理特性来适应环境,使其形态可塑性发生改变(Givnish,1988)。在膝柄木天然更新的影响因素中,光因子是影响其生长的重要环境因子之一,加强环境中光因子对膝柄木影响的研究,了解光因子在膝柄木天然更新中所起的作用,对于膝柄木种群的扩大具有重要的意义。此外,膝柄木对于研究我国和印度、老挝、新加坡等10 国的植物区系之间的联系以及广西地植物学和植物生态学的研究具有重要的科学意义(梁盛业,1992)。如何科学合理地给膝柄木提供最适宜生长的环境,了解膝柄木的生态适应性及其生长限制因子,可为濒危树种膝柄木的有效保护提供理论指导,同时也为濒危物种的生态恢复和保护提供理论依据。本研究以人工培育之后再移植到原生环境中的膝柄木幼树为研究对象,通过对三种不同生境下的膝柄木幼树进行试验探究,对比不同生境,膝柄木幼树的生长特征、生理特性、光合色素等生长指标,研究观测其对光合有效辐射需求的年际动态变化响应,以揭示光在膝柄木幼树天然更新过程中的限制性影响,同时探究膝柄木濒危原因,从而为膝柄木的天然更新,种群保护和生态恢复提供科学依据。
1 材料与方法
1.1 研究地概况
研究地点位于广西东兴市东部江平镇巫头村,地理位置为108°07′ E、21°32′ N,属湿热带季风气候区,海拔20~40 m。8 月份测定的试验地林缘、林窗和林下光合有效辐射分别为(1 743±138)μmol·m-2·s-1、(1 363±96)μmol·m-2·s-1和(176±28)μmol·m-2·s-1。试验前测定林缘、林窗、林下三种生境土壤pH值为4.1~4.5,有机质含量为20.96~22.35 g·kg-1,全氮为1.21~1.46 g·kg-1,有效磷为4.8~5.2 mg·kg-1,速效钾为31.88~43.83 mg·kg-1,差异不显著。
1.2 材料与试验设计
试验材料采自广西东兴市东部江平镇巫头村2009年人工培育、长势良好的回种到原出生地树林的林缘、林窗、林下三种生境中(通过采集种子进行人工培育)的膝柄木幼树。并于2015年12 月至2016 年10 月期间选取自然条件下生长良好、植株基本一致的林缘、林窗和林下三种不同生境人工种植的膝柄木幼树,对其生长指标(地径、株高、叶面积)进行测定,每种生境下选择幼树各3株,此外在此期间的2、4、6、8、10、12月中旬选取不同生境下膝柄木幼树东南西北四个方向上树冠中部枝条下4~6 片长势良好的成熟叶片,放入冰盒带回实验室,进行生理指标和光合指标的测定。在试验期间及时对幼树进行除草防治病虫害。
1.3 测定项目与方法
1.3.1 苗木生长指标的测定 在试验开始前,在每株幼树的根茎处用红色布条绑住做好标记,用钢卷尺(精确度为0.1cm)测量膝柄木幼树根部作记号到顶芽的高度。用游标卡尺(精确到0.01 mm)精确测量每株苗木根茎处的直径。同时,记录下每株树的叶片数量,使用激光叶面积仪测量葉面积。根据前后两次测得的数值差计算苗木的地径、叶面积和株高净增长量。
1.3.2 叶片生理生化指标的测定
1.3.2.1 抗氧化酶活性测定 超氧化物歧化酶(superoxide dismutase,SOD)活性采用氮蓝四唑(nitroblue tetrazolium,NBT)光化还原法测定,依据NBT在光下可被还原,被还原后的核黄素在氧化物质的存在下氧化产生O2-,NBT被氧化后的O2-还原成蓝色甲腙,通过测定甲腙在560 nm处的吸光度来测量SOD活性(李合生等,2000)。
过氧化氢酶(catalase,CAT)活性采用紫外光吸收法测定,在单位时间内测定波长240 nm处吸光度的差值(Aebi, 1984)。
1.3.2.2 可溶性蛋白和游离脯氨酸含量测定
可溶性蛋白含量采用考马斯亮蓝G-250染色法测定,依据考马斯亮蓝G-250在稀酸溶液中遇蛋白质变色的原理,通过在波长为520 nm处的吸光度值来计算可溶性蛋白含量(李合生等,2000)。
游离脯氨酸含量使用茚三酮比色法测定,在待测植物叶片中加入3%的磺基水杨酸溶液提取脯氨酸,再在脯氨酸提取液中加入冰醋酸和酸性印三酮,沸水浴加热,变色后加入甲苯离心后在分光光度计波长为520 nm处测定吸光度值(李合生等,2000)。
1.3.2.3 质膜透性和丙二醛(malonaldehyde,MDA)含量测定 MDA含量采用硫代巴比妥酸法(thiobarbituric acid,TBA)测定,将0.5 g的植物样品中加入5%的三氯乙酸(trichloroacetic acid,TCA)研磨,将匀浆离心后取上清液加入0.67%的TBA,煮沸后再次离心,测定上清液在450、532、600 nm处的吸光值(李合生等,2000)。
质膜透性采用电导仪法测定,将2 g大小切割一致的叶片放入真空干燥器内,加入20 mL的去离子水,抽气后缓缓放入空气,待叶片下沉后,取出烧杯静置20 min后用玻璃棒搅拌均匀,用电导仪测其初电导值(S1),测完后将烧杯放入沸水浴中,用自来水冷却至室温后测其煮沸电导值(S2),相对电导率(L)=S1/S2 (李合生等,2000)。
1.3.2.4 光合色素測定 光合色素采用丙酮-乙醇混合液提取法测定(李合生等,2000),依据叶绿素提取液可吸收可见光谱的原理,将待测的叶片剪碎,加入少量的石英砂和碳酸钙粉以及95%乙醇后研磨至组织变白,用95%的乙醇将滤纸上的色素洗入容量瓶中定容,根据665、649、470 nm波长处测得的吸光度值计算光合色素浓度,公式如下:
叶绿素a浓度:Ca (mg·L-1)=13.95 A663-6.88 A645;叶绿素b浓度:Cb (mg·L-1)=24.96 A645-7.32 A633;
叶绿素a/b浓度:Ca/b浓度=Ca /Cb;类胡萝卜素浓度:Cx·c=(1 000×A470-2.05 Ca-114.8 Cb)/245;
相应色素的含量(mg·g-1)=(Ci×V×d)/m。
式中:Ci为相应色素的浓度;V表示提取液体积;d表示稀释倍数;m表示样品鲜重。
1.3.3 叶片光合生理指标测定 选择天气晴朗的上午(9:00—11:00),采用Li-6400光合测定仪对植株叶片净光合速率Pn、胞间CO2浓度Ci、蒸腾速率Tr进行测定。并计算水分利用效率(water use efficiency,WUE)=Pn/Tr。
1.4 数据处理
运用Excel软件整理数据,采用SPSS 25.0软件进行数据处理和分析,用单因素方差分析(one-way ANOVA)结合Duncan检验组间差异性,使用Origin 2018软件绘制图像。表格和图表中的数据均为平均值±标准偏差。
2 结果与分析
2.1 不同生境下膝柄木幼树的生长的影响
苗木的生长状况主要是通过其生长指标反映的。从表1可知林下生境的幼树由于接收到的光照较少,生长受到了抑制,其株高和叶面积的增长量随着时间的增加逐渐减少。生长在林缘的植株生长状况最好,其株高和叶面积的增长量均最大,与林窗的株高增长量和叶面积增长量之间都达到了显著差异(P<0.05);生长在林缘的幼树由于得到的光合有效辐射显著高于林下,因此,林缘与林下的膝柄木幼树的各项生长指标都达到了显著差异(P<0.05)。
2.2 不同生境的膝柄木植株生理生化特征的年际动态变化
SOD、CAT、MDA、可溶性蛋白、游离脯氨酸和相对电导率是植物重要的生理生化指标。图1是林缘、林窗、林下三种生境下膝柄木幼树的生理生化指标的年际变化,其中三种生境中幼树的抗氧化酶活性、可溶性蛋白、相对电导率和MDA的年际变化呈先增加后降低的趋势,游离脯氨酸含量则先降低后增加。生长于林缘的幼树生理指标与林下的差异显著,生长于林缘的幼树的SOD和CAT、可溶性蛋白、相对电导率和MDA含量在一年中显著高于林下,而林下膝柄木幼树的游离脯氨酸含量的年际值则高于林缘和林窗的幼树。
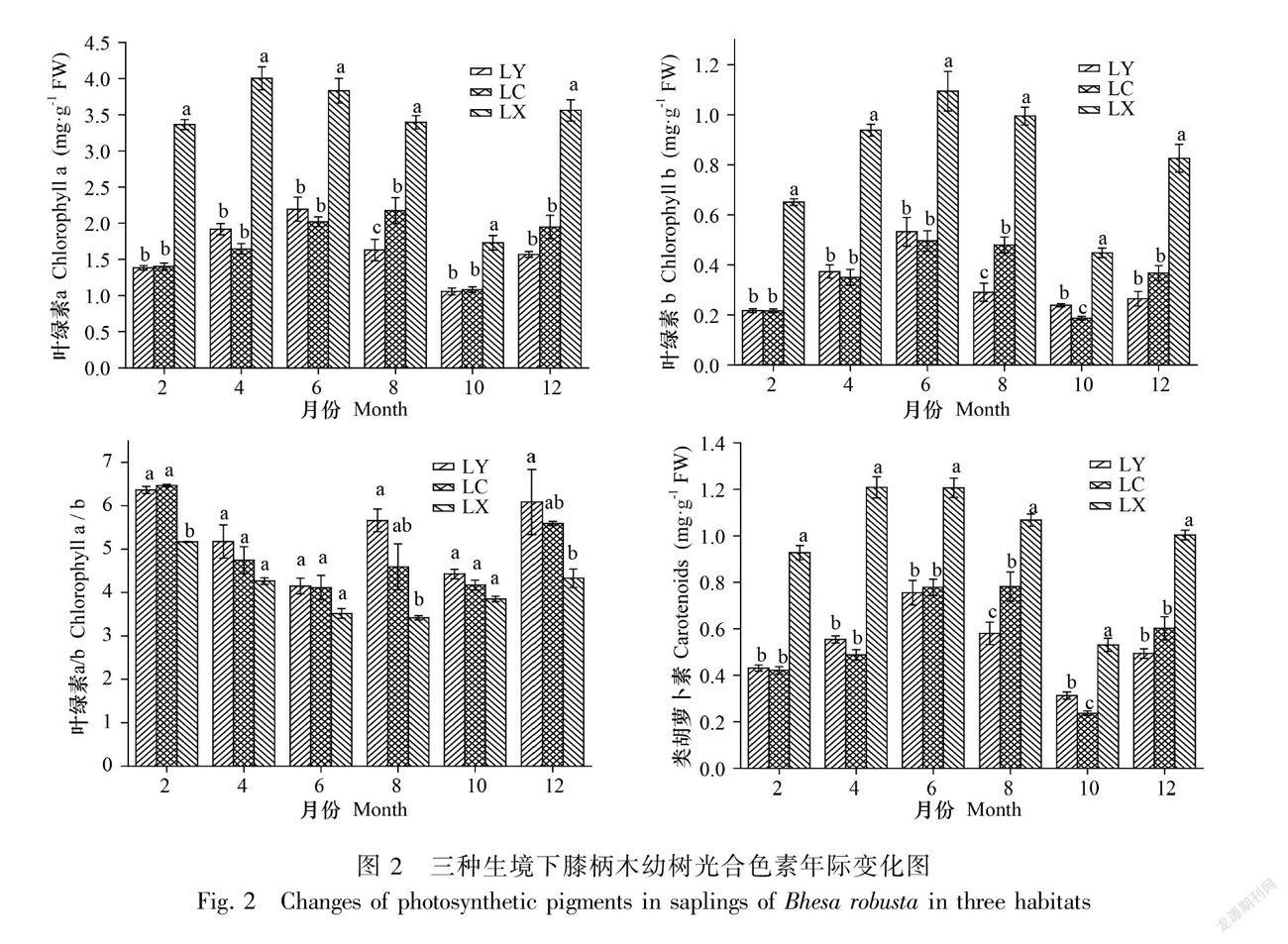
2.3 不同生境对膝柄木幼树光合色素的影响
光合色素体系是植物体内捕获光能的重要光合系统,在本试验中随着不同生境所接受到的光合有效辐射的变化,其光合色素的含量和组成也发生了显著变化。图2表明各生境膝柄木幼树的叶绿素a、叶绿素b以及类胡萝卜素含量在一年中随着光合有效辐射呈先增加后减少的趋势,林下生境的膝柄木幼树的各光合色素含量在一年中显著地高于林缘和林窗生境的幼树,叶绿素a/b的值林下生境的幼树最低。
2.4 不同生境对膝柄木幼树叶片光合生理特征的影响
如图3所示,膝柄木幼树的Pn和Tr的年际变化趋势为先增加后降低。林缘幼树的Pn在6 月份达到了最大值,而林窗幼树的Pn在8 月份达到最大值,林缘幼树的Tr在6 月份有最大值,在8 月下降,在8 月之后Tr又开始增加,而林下生境幼树的Pn和Tr总体上呈先增加后降低的趋势,但年际变化不大。膝柄木幼树的Ci和WUE则是先降低后增加的趋势。林下生境幼树Ci在一年中显著高于林缘生境。2—4月林缘的WUE显著高于林下,在6—12月,林缘幼树的WUE逐渐低于林窗和林下生境,而林下生境幼树的WUE在4 月份开始显著高于其他两种生境。
3 讨论与结论
3.1 膝柄木的生长适应性
植物为了生存要适应环境因子截然不同的生境,处于不利生境时,植株的生长就会受到环境因子的影响,从而使植株的生长受到阻碍,使种群的生存面临威胁。弱光会影响植物的生长,使植物干物质积累减少,影响植株开花结果和植株产量(Parelle & Ducrey,2006)。也有研究表明适当的弱光条件能够促进植株花芽分化,提高植株的花芽数(Menzel & Simpson,1988)。此外,有学者发现一些物种分布于林窗、林缘等不同生境其生长和发育并无显著差异,都能够实现其物种的更新(Hamerlynck & Knapp,1994)。这说明了不同的物种由于其进化出不同的生理特性,对光的适应能力大有不同,而同一物种,处于不同的生境时其生理生态特性也会发生显著的变化(Matsubara et al.,2008)。不同的生境中特有的环境因子对植株的生长发育都具有显著影响(王艺和韦小丽,2009)。
光照不足还容易产生秧苗徒长现象,即植株表现为根系不发达、叶片稀少、植株生长缓慢及早衰现象(侯兴亮等,2002)。膝柄木幼树在林缘、林窗、林下三种生境下,相同的生长指标在不同生境的幼树中差异显著,在本研究中,生长于林缘的膝柄木幼树接受到的光照最好,其地径、株高和叶面积增长量均显著高于林下生长的膝柄木幼樹,光合有效辐射的不足,使得林下生境膝柄木幼树的地径、株高、叶面积增长量相比林缘显著降低了,说明膝柄木幼树时期,需要较好的光照条件,光成为其主要限制因子,膝柄木幼树为喜光物种。
3.2 膝柄木幼树的生理生化适应性
光照是影响植物生理特性的重要因素之一(杨生保等,2011),植物在长期的进化过程中形成了抵抗不利环境的保护酶,如抗氧化活性酶SOD和CAT等,这些酶组成了植物体内的酶促系统,清除植物产生的大量活性氧,从而维持植物的正常代谢活动(黄尧瑶等,2020)。然而,当植物接受到的有效光合辐射不足时,体内的抗氧化酶活性就会降低,从而使得植物清除自由基的能力降低,植物生长缓慢(杨生保等,2011)。随着生长于林下生境的膝柄木幼树接受到的光合有效辐射减少,林下生境的膝柄木幼树的SOD和CAT酶活性显著低于林缘生境,光因子对膝柄木幼树的调控作用受到了限制,膝柄木幼树清除对细胞有害自由基的能力降低,膝柄木幼树生长受到抑制。可溶性蛋白是植物调控和促进植物代谢的主要作用物质,其含量高低与植物代谢能力强弱有关(段辉国等,2014),光照不足会使植物体内的可溶性蛋白含量降低,从而导致植物的新陈代谢能力降低,植物衰老加快。MDA是反应植物抗逆能力的指标(张恩平等,2001),相对电导率是反映植物膜系统状况的重要生理生化指标,相对电导率增大说明植物叶片受到了损害(朱凤荣,2019)。在本试验中,随着林下生境的膝柄木幼树接收到的光合有效辐射的减少,幼树体内的抗氧化活性酶SOD和CAT含量、可溶性蛋白含量、MDA和相对电导率含量林下生境的幼树均显著低于林缘生境幼树,表明膝柄木幼树在长期光照不足的条件下,可溶性蛋白、MDA含量的合成受到抑制,幼树的细胞膜系统生理活性低,幼树代谢水平较低,代谢能力受到阻碍,植物的生长受到了抑制。一些研究表明,植物在逆境下积累游离脯氨酸是植物受到了伤害的结果,对限制因子反应敏感的植株脯氨酸积累会显著增多(Moftah & Michel,1987)。植物在正常条件下,体内的游离脯氨酸含量较低,但在遇到不利的生长逆境时,游离脯氨酸含量就会大量积累。在本试验中,生长于林下的膝柄木幼树周围郁闭的林荫,接收到的有效光合辐射较少,因此,光照不足使得幼树的生长受到了胁迫,为了更好地适应逆境,生长于林下的幼树体内的游离脯氨酸含量显著增加,以对抗不利的环境(位杰等,2014)。
3.3 膝柄木幼树的光合色素适应性
叶绿素是植物体内吸收能量的主要物质,叶绿素含量的高低,体现了植物光合作用能力的大小(位杰等,2014)。在光照不足的环境中,植物通过增加体内的叶绿素含量来吸收更多的光能,通过提高光合作用的速率以更好地适应弱光条件(吴飞燕等,2012)。光照不足时,植物体内的叶绿素a和叶绿素b的含量会显著增加,与叶绿素a含量相比,叶绿素b增加的幅度较大(Singh & Dey,1988),因此林下生境的叶绿素a/b含量下降。此外,研究发现环境因子对于类胡萝卜素的合成途径具有一定的调控作用(Harding & Shorpshire,1980),光合有效辐射的强弱影响了类胡萝卜素在植物体内的含量高低。在本研究中,随着林下生境的膝柄木幼树接受到的光合有效辐射的减少,幼树体内的叶绿素a、叶绿素b含量和类胡萝卜素含量相比林缘生境的幼树均显著增加,叶绿素a/b的值林缘最高。这是由于林下接收到的入射光较少,光合系统由于为了捕捉更多的太阳能,而增加了光和色素的含量,提高光能转化效率。植物通过这种方式能够将微弱的漫射光充分利用,来保证体内的光合化学反应正常进行,从而提高其光合利用效率,以保证植物能够充分适应林下的生境。
3.4 膝柄木幼树的光合生理适应性
植物的Pn和Tr是植物光合能力大小的直接反映(马新等,2018)。弱光条件下,植物的Pn降低,植物的光合作用减弱(杨冠松等,2014)。光合有效辐射的不足,使得植物的Pn曲线由双峰型转变为单峰型,植物正常的生长发育受到了影响(岳高峰等,2020)。在本研究中,林缘膝柄木幼树的Pn呈双峰曲线,林窗和林下生境的膝柄木幼树呈单峰曲线,这表明生长于林下生境的膝柄木幼树由于接收到的光合有效辐射不足,幼树的Pn曲线由林缘的双峰型曲线转变为了林下的单峰型曲线,弱光超过了植物的耐受范围,对林下生境的膝柄木幼树的正常生长产生了胁迫。Tr是植物调节自身水分亏缺,说明能更好适应环境能力的重要指标(张江涛等,2007),植物的Tr与Pn强度呈正相关(杨冠松,2014),在本试验中,林下生境膝柄木幼树的Tr变化与Pn变化一致,光合有效辐射的不足抑制了植物的Tr,林下幼树的物质运输和营养吸收的生理过程受到抑制,植物的生长发育受到了影响。Ci是植物能量的供给源,CO2进入细胞后主要受植物体内光合速率的影响(Flexas & Medrano,2002)。光合有效辐射不足,使得林下膝柄木幼树叶肉细胞的光合活性降低,幼树在光合作用过程中消耗CO2的能力降低,林下生境膝柄木幼树Ci增高。WUE是表征植物利用水分能力强弱的生理参数,是植物生长发育所需最佳水分供应的重要指标之一(钟平安等,2019),植物叶片的WUE也是反映植物自身蒸腾作用大小、适应生境能力的重要参数(李轲等,2017)。光照强烈的4月到10月,林缘和林窗生境的幼树,蒸腾作用强,水分利用效率下降,生长于林下的膝柄木幼树WUE高。12月至次年2月,林缘和林窗生境幼树的蒸腾作用不强,温度适宜,WUE高于林下生境的膝柄木幼树。本试验表明膝柄木幼树在生长发育阶段在保证其受到充足光照的同时也要避免强光辐射带来的水分消耗。
综上所述,生长于光照充足、环境开阔的林缘生境下膝柄木幼树的生长状况要显著优于其他两种生境,林缘生境膝柄木幼树各生长指标增长量最大,生长发育状况最好,光合能力和光合效率高。光照不足限制了林下膝柄木幼树的株高、地径和叶面积的增长。在膝柄木幼树的天然更新和种群的恢复与扩大时,应保证其处于较佳的光照环境,更有利于其正常的生长发育。膝柄木幼树对弱光环境响应和适应的差异主要表现在生物量分配及生长差异上。野外的膝柄木幼树大都生长于林下,其天然更新受到阻碍,因此,在膝柄木幼树的生态恢复过程中可以采用疏伐、透光伐等手段来降低上层林冠的郁闭度,或者移苗种植等途径来促进膝柄木幼树的自然更新,保证其正常生长。
参考文献:
AEBI H,1984. Methods in enzymology: oxygen radicals in biological systems(Vol105) [M]. Academic Press: 121-126.
AN JC, ZHANG ZY, HUANG MP, 2017. Genetic relationship of three individuals of Bhesa sinica by RAPD[J]. Guangxi For Sci, 46(4): 406-408. [安家成, 张照远, 黄妹平, 2017. 膝柄木三个单株RAPD的亲缘关系[J]. 广西林业科学, 46(4): 406-408.]
DUAN HG, WANG B, HUANG ZX, et al., 2014. Effects of chitosan on seed germination and seedling physiological index of hulless barley[J]. Henan Agric Sci, 43(3): 16-19. [段辉国, 王彬, 黄作喜, 等, 2014. 壳聚糖对青稞种子萌发和幼苗生理指标的影响[J]. 河南农业科学, 43(3): 16-19.]
FLEXAS J, MEDRANO H, 2002. Drought-inhibition of photosynthesis in C3plants:Stomatal and non-stomatal limitations revisited[J]. Ann Bot, 89(2): 183-189.
GIVNISH TJ, 1988. Adaptation to sun and shade:A whole-plant perspective[J]. Aust J Plant Physiol, 15:63-93.
HAMERLYNCK EP, KNAPP AK, 1994. Leaf-level responses to light and temperature in two co-occurring Quercus (Fagaceae) species: mplications for tree distribution patterns[J]. For Ecol Manag, 68(2-3): 149-159.
HARDING RW, SHORPSHIRE WJ, 1980. Photocontrol of carotenoid biosynthesis[J]. Ann Revpllant Physiol, 31: 217-238.
HOU XL, LI JF, XU XY, 2002. Effects of low light on morphological and physiological indexes of tomato at different growth stages[J]. Acta Hortic Sin, 29(2): 123-127. [侯興亮, 李景富, 许向阳, 2002. 弱光处理对番茄不同生育期形态和生理指标的影响[J]. 园艺学报, 29(2): 123-127.]
HUANG LX, 2006. Several plants in danger of extinction in China[J]. For Guangxi, (5): 58. [黄立新, 2006. 我国濒临灭绝的几种植物[J]. 广西林业, (5): 58.]
HUANG YY, DENG MH, PENG CX, et al., 2020. Studies on the response of lily petal antioxidant enzyme system to drought stress[J]. Acta Hortic Sin, 47(4): 788-796. [黄尧瑶, 邓明华, 彭春秀, 等, 2020. 百合花瓣抗氧化酶系统对干旱胁迫响应的研究[J]. 园艺学报, 47(4): 788-796.]
LI HS, SUN Q, ZHAO SJ, et al., 2000. Principles and techniques of plant physiological and biochemical experiments [M]. Beijing: Higher Education Press: 134-261. [李合生, 孙群, 赵世杰, 等, 2000. 植物生理生化实验原理和技术 [M]. 北京: 高等教育出版社: 134-261.]
LI K, JING YS, TAN MX, et al., 2017. Effect of different irrigation amounts on physiology and water use efficiency of rice at jointelongation and booting stage[J]. Jiangsu Agric Sci, 45(13): 60-63. [李轲, 景元书, 谭孟祥, 等, 2017. 不同灌水量对拔节孕穗期水稻生理及水分利用率的影响[J]. 江苏农业科学, 45(13): 60-63.]
LIANG SY, 1988. The Bhesa sinica was first discovered in China[J]. Guangxi For Sci Technol, (1): 34. [梁盛业, 1988. 膝柄木属植物在我国首次发现[J]. 广西林业科技, (1): 34.]
LIANG SY, 1992. A rare and endangered plant—Bhesa sinica[J]. Plants, (3): 5. [梁盛业, 1992. 珍稀濒危植物——膝柄木[J]. 植物杂志, (3): 5.]
LIU Y, NING SJ, 2002. Status quo and evaluation of natural resources of emphasis protective wilding plants in Guangxi[J]. Guangxi Sci, 9(2): 124-132. [刘演, 宁世江, 2002. 广西重点保护野生植物资源的现状与评价[J]. 广西科学, 9(2): 124-132.]
MA X, DONG P, JIANG JY, et al., 2018. Diurnal variation of photosynthesis rate of Xanthoceras sorbifolia and correlation with eco-physiological factors[J]. SW Chin J Agric Sci, 31(6): 1267-1271. [馬新, 董鹏, 姜继元, 等, 2018. 文冠果光合生理日变化及其与生理生态因子的相关性研究[J]. 西南农业学报, 31(6): 1267-1271.]
MATSUBARA S, KRAUSE GH, SELTMANN M, et al., 2008. Lutein epoxide cycle, light harvesting and photoprotection in species of the tropical tree genus Inga[J]. Plant Cell Environ, 31(4): 548-561.
MENZEL CM, SIMPSON DR, 1988. Effect of continuous shading on growth, flowering and nutrient uptake of passionfruit[J]. Elsevier, 35(1-2): 77-88.
MO ZC, FAN HQ, LI LX, et al., 2008. The surviving situation and restorative counter-measures of endangered plant Bhesa sinica[J]. J Guangxi Acad Sci, 24(2): 134-137. [莫竹承, 范航清, 李蕾鲜, 等, 2008. 濒危植物膝柄木生存现状及其恢复对策[J]. 广西科学院学报, 24(2): 134-137.]
MO ZC, PANG WW, LIU Y,et al., 2015. Study on callus induction of Bhesa robusta leaves[J]. J CS Univ For & Technol, 35(10): 13-17. [莫竹承, 庞万伟, 刘珏, 等, 2015. 膝柄木叶片诱导愈伤组织研究[J]. 中南林业科技大学学报, 35(10): 13-17.]
MO ZC, XU JQ, CHEN SY, 2013. Phenological characters of dominant trees in Syzygium hancei Monsoon forest[J]. Guangxi Sci, 20(3): 193-198. [莫竹承, 徐剑强, 陈树宇, 2013. 红鳞蒲桃季雨林重要树种的物候特征[J]. 广西科学, 20(3): 193-198.]
MOFTAH AE, MICHEL BE, 1987. The effect of sodium choloride on solute potentialand praline accumulation in soybean leaves[J]. Physiol Plant, 83(2): 238-240.
PARELLE JP, DUCREY M, 2006. Light acclimation and photosynthetic response of beech (Fagussylvatica L.)saplings under artificial shading or natural Mediterranean conditions[J]. Ann For Sci, 63(3): 257-266.
SINGH VP, DEY S, 1988. Effect of low light stress on growth and yield of rice[J]. Indian J Plant Physiol, 31 (1): 84-91.
SUN QL, ZHANG RH, YI SG,et al., 2019. Responses of germination and seedling emergence of Stipa bungeana to key environmental factors[J]. Acta Ecol Sin, 39(6): 2034-2042. [孙清琳, 张瑞红, 易三桂, 等, 2019. 长芒草(Stipabungeana)种子萌发与出苗对关键环境因子的响应[J]. 生态学报, 39(6): 2034-2042.]
WANG Y, WEI XL, 2009. The effect of different light intensity on seedling ecological adaptability of two karst tree species[J]. Guizhou Agric Sci, 37(5): 139-142. [王艺, 韦小丽, 2009. 两个喀斯特森林树种幼苗对不同光照的生态适应性[J]. 贵州农业科学, 37(5): 139-142.]
WEI J, WU CY, WANG HL, et al., 2014. Effects of different light intensity on physiological characteristics in leaves of Ziziphus jujuba cv. Huizao[J]. Henan Agric Sci, 43(12): 112-116. [位杰, 吴翠云, 王合理, 等, 2014. 不同光照强度对灰枣叶片生理特性的影响[J]. 河南农业科学, 43(12): 112-116.]
WU FY, YI LT, LI XP, et al., 2012. Effect of different light intensity on intensity chlorophyll content and chlorophyll fluorescence in Lithocarpus glaber[J]. J NE Agric Univ, 43(4): 88-92. [吴飞燕, 伊力塔, 李修鹏, 等, 2012. 不同光照强度对石栎幼苗叶绿素含量及叶绿素荧光参数的影响[J]. 东北农业大学学报, 43(4): 88-92.]
WU XQ, HUANG BL, DING YL, 2004. The advance on the study of protection of rare and endangered plants in China[J]. J Nanjing For Univ (Nat Sci Ed), 28(2): 72-76. [吴小巧, 黄宝龙, 丁雨龙, 2004. 中国珍稀濒危植物保护研究现状与进展[J]. 南京林业大学学报(自然科学版), 28(2): 72-76.]
XU F, MOU JP, LIANG SY, 1990. Wood anatomy of he Bhesa—New recorded enus of Chinese Celastraceae[J]. Chin Bull Bot, 7(3): 52-54. [徐峰, 牟继平, 梁盛业, 1990. 中国卫矛科新记录属——膝柄木属的木材解剖(简报)[J]. 植物学通报, 7(3): 52-54.]
YANG GS, ZHANG AL, SHEN SK,et al., 2014. Effects of shading on the photosynthetic characteristics and chlorophyll fluorescence of the endangered plant Yunnanopilia longistaminata[J]. Guihaia, 34(6): 806-811. [杨冠松, 张爱丽, 申仕康, 等, 2014. 遮荫对濒危植物甜菜树光合生理和叶绿素荧光特性的影响[J]. 广西植物, 34(6): 806-811.]
YANG SB, PATI GL, WANG BK, et al., 2011. Effects of sunlight on processing tomato plug-seedling and its physiology[J]. Xinjiang Agric Sci, 48(9): 1617-1623. [楊生保, 帕提古丽, 王柏柯, 等, 2011. 光照对加工番茄穴盘苗植株形态及生理的影响[J]. 新疆农业科学, 48(9): 1617-1623.]
YUE GF, HAN ZQ, XUE ZW, et al., 2020. Effects of shading on flowering period and photosynthetic characteristics of peony[J]. Jiangsu Agric Sci, 48(17): 166-170. [岳高峰, 韩志强, 薛志伟, 等, 2020. 不同遮阴对牡丹花期和光合特性的影响[J]. 江苏农业科学, 48(17): 166-170.]
ZHANG EP, ZHANG SH, SI LT, et al., 2001. Effects of NaCl stress on the membrance lipid peroxidation in cotyledon of cucumber seedlings[J]. J Shenyang Agric Univ, 32(6): 446-448. [张恩平, 张淑红, 司龙亭, 等, 2001. NaCl胁迫对黄瓜幼苗子叶膜脂过氧化的影响[J]. 沈阳农业大学学报, 32(6): 446-448.]
ZHANG JT, LIU YQ, ZHAO PH, et al., 2007. Comparison of photosynthetic and physiological characteristics of occidental poplar young plants[J]. J CS Univ For Technol, 27(4): 8-11. [张江涛, 刘友全, 赵蓬晖, 等, 2007. 欧美杨无性系幼苗的光合生理特性比较[J]. 中南林业科技大学学报, 27(4): 8-11.]
ZHONG GG, XIE ST, SU FB,et al., 2016. Study on seed germination of rare and endangered plant Bhesa sinensis[J]. J N Hortic, (5): 83-85. [鐘国贵, 谢绍添, 苏付保, 等, 2016. 珍稀濒危植物膝柄木种子催芽研究[J]. 北方园艺, (5): 83-85.]
ZHONG PA, SHAO D, HUANG YJ,et al., 2019. The photosynthesis characteristics and instantaneous water-use efficiency of Capsicum annuum under different light conditions[J]. Acta Ecol Sin, 38(7): 2065-2071.[钟平安, 邵东, 黄英金, 等, 2019. 不同光环境下辣椒光合特性和瞬时水分利用效率[J]. 生态学杂志, 38(7): 2065-2071.]
ZHU FR, 2019. Effects of drought stress on physiological and biochemical characteristics of three herbaceous plants[J]. Hubei Agric Sci, 58(6): 88-91. [朱凤荣, 2019. 干旱胁迫对三种草本植物生理生化特性的影响[J]. 湖北农业科学, 58(6): 88-91.]
(责任编辑 李 莉)
收稿日期:2020-11-05
基金项目:国家自然科学基金(31260093);广西自然科学基金(2013GXNSFAA019057)[Supported by National Natural Science Foundation of China (31260093); Natural Science Foundation of Guangxi (2013GXNSFAA019057)]。
第一作者: 招礼军 (1970-),博士,教授,主要从事树木生理生态研究,(E-mail)zhlj-70@163.com。
通信作者:朱栗琼,副教授,主要从事植物形态与生理生态研究,(E-mail)liqiongzhu@163.com。
