基于SRAP分子标记的何首乌种质遗传多样性和群体结构研究
2022-04-01李嘉惠欧晓华刘晓莹卢昌华张宏意何梦玲严寒静
李嘉惠,欧晓华,刘晓莹,卢昌华,张宏意, 2,何梦玲, 2,严寒静, 2*
基于SRAP分子标记的何首乌种质遗传多样性和群体结构研究
李嘉惠1,欧晓华1,刘晓莹1,卢昌华1,张宏意1, 2,何梦玲1, 2,严寒静1, 2*
1. 广东药科大学中药学院,广东 广州 510006 2. 国家中医药管理局岭南药材生产与开发重点研究室,广东 广州 510006
探究何首乌种质的遗传多样性和居群结构,为何首乌种质资源的保护及合理利用提供参考。采用SRAP分子标记方法,利用Structure 2.3.4、NTSYSpc、Popgen 32和MEGA 7软件对数据进行统计,获得何首乌居群的多样性水平、遗传距离、遗传分化程度、聚类结果和群体分析结果。何首乌野生居群遗传多样性大于栽培居群,多样性主要来自居群间。野生居群中,四川和重庆居群的遗传多样性较高,何首乌样品间遗传距离与地理距离无明显相关性;何首乌样品的邻接法(neighbor joining,NJ)聚类结果与地理分布相符,相同采集点样品可聚在一起。结果还显示农户栽培品亲缘关系近,说明品种间存在相互引种情况。居群结构分析表明,大部分何首乌样品的血缘组成较为单一,且=16时,样品分类效果最佳,而分类结果则与NJ聚类基本一致。SRAP分子标记是何首乌遗传多样性估计和种群关系的有效分析工具,而地形复杂性可能是影响何首乌遗传多样性程度的重要因素,另外,广西所产棱枝何首乌在何首乌种群中表现出特异性,与其他种源遗传距离大,支持将其列为何首乌的变种。
何首乌;SRAP;遗传多样性;种质; 亲缘关系
何首乌Thunb为蓼科多年生草本植物,其地下膨大的块根,生则截疟解毒、润肠通便,炮制后可补益精血、固肾乌须,而现代研究表明,何首乌块根具有调血脂和抗衰老等的作用[1]。随着人们生活水平的提高及对保健领域的关注,具有补益功效的何首乌需求量日渐攀升,作为本品主流的野生何首乌在掠夺式的采挖中正以每年约15%的速度递减[2]。有学者调查了我国何首乌野生品种的分布区域,并且对其蕴藏量作了估算,发现调查的60多个样方当中,能采挖出来的一般为何首乌幼根,质量在10 g左右,单个样方的蕴藏量还不足50 g,所走访的湖北、广西田林、贵州和河南嵩县等地的何首乌野生品种也呈下降的趋势[3],而目前人工栽培的规模还未能满足市场的需要,同时长期的扦插繁殖已导致栽培种质的退化[4],何首乌种质资源正面临严峻的挑战。
植物的种质资源,是植物在常年演化过程中,在经历自然选择、人工驯化后形成的可以传递给下一代的遗传物质的总称,凡是携带有遗传信息的野生品种、栽培品种、变种和古老地方品种等都属于种质资源[5]。植物种质资源的多样性,一方面决定了植物种群在自然界中的生存能力和应对环境变化的适应能力,影响生存环境的生态平衡;另一方面,作为人类可利用的资源,它所蕴含的遗传物质可以通过优良种质挖掘,抗性基因的筛选为人类带来效益[6]。因此,为应对何首乌资源日渐匮乏的境况,亟待加强何首乌种质多样性的研究,为其种质保护奠定基础。
目前物种遗传多样性研究方法包括形态学标记方法、生化标记方法、细胞学标记方法和分子标记方法[5]。由于植物形态受环境的影响,因此以形态数据对种质进行研究往往会损失较多的遗传多样性,较难反映物种的真实情况,而分子标记则是DNA分子水平的直接反应,不受外界环境因素的影响,在植物生长的任何部位都可以检测到,位点多且丰富,检测较快[7],因而更能代表种质的多样性。在植物研究领域一般使用的标记方法包括了SSR、ISSR、AFLP、RFLP、RAPD和SRAP等,其中ISSR[8]、RAPD[9]和SRAP(sequence related amplified polymophism)[10]分子标记方法在何首乌遗传多样性研究中已有使用。SRAP标记方法是由Li等[11]于2001年创立的,相对于其它标记方法,它具备分布普遍、可靠性和重复性高、技术难度低和多态性高等特点[12],在国内外已广泛应用于农作物的研究[13]。程远辉[10]在研究重庆地区何首乌遗传多样性时,曾较长时间使用RAPD标记方法进行研究,结果显示扩增条带少,重复性差,所以最终选择SRAP方法进行研究,可见SRAP标记方法在何首乌遗传多样性研究上的优势。
因此,本实验采用SRAP分子标记方法,对全国12个省44个居群的327份何首乌样品进行研究,并以此探究我国何首乌的种质多样性和居群结构,为何首乌种质资源的保护及合理利用提供参考。
1 材料和方法
1.1 材料
于2017—2019年,采集327份样品,经广东药科大学中药学院刘基柱副教授鉴定为蓼科植物何首乌Thunb.。样品覆盖我国12个省44个地点,其中218份为栽培品,109份为野生品,栽培品中包含一份棱枝何首乌(LZ),样品详细信息见表1、2。
1.2 方法
选用植物DNA提取试剂盒(宝生物工程有限公司,产品型号9768)对何首乌叶片的基因组DNA进行提取。
利用前期筛选得到的13对引物对327份何首乌基因组DNA进行扩增,其中反应体系的总体积为20 µL,即10 µL premix(宝生物工程有限公司,产品型号RR902A),上下游引物(引物母液质量浓度为10 ng/µL)各1 µL,DNA模板为40 ng,并用ddH20补充体积至20 µL,另外PCR反应为SRAP标记方法的一般程序,即94 ℃预变性4 min;94 ℃、1 min,35 ℃、1 min,72 ℃、1 min,共5个循环;94 ℃、1 min,50 ℃、1 min,72 ℃、1 min,共35个循环,最后72 ℃延伸5 min。
表1 何首乌栽培品来源
Table 1 Origin of P. multiflorum cultivars
省份地点经度(E)纬度(N)海拔/m时间数量/份样品编号 广东德庆莫村镇太宪村112°05′31″23°21′56″602018-0420TX1~20 德庆新圩镇大同村111°55′51″23°20′07″202018-0420DT1~20 高州石鼓镇白泥塘村110°47′47″21°49′03″202017-0720BNT1~20 新兴天堂镇山塘角村112°01′05″22°34′36″202018-0718STJ1~18 德庆大桥村春牛亭111°47′22″23°08′26″302018-0420CNT1~20 广西贵港109°23°−2019-0526GG1~26 贵州施秉牛大场镇山口村107°48′44″26°20′05″800以上2018-0420SKC1~20 上海上海中医药大学121°35′57″31°11′39″102018-069SZY1~9* 益大本草园121°37′57″31°08′51″−1002018-06 1YD1* 愚园路121°25′57″31°13′15″702018-06 1YYL-1* 江西江西中医药大学115°44′43″28°40′45″602018-06 9JZY1~9* 江苏南京中医药大学118°95′32″32°10′68″−2017-11 8HC1~8* 南京中医药大学118°94′85″32°10′54″−2017-1111YY1~11* 陕西陕西中医药大学108°44′19″34°19′03″343.52019-0420SXZ1~20* 河南河南中医药大学113°48′07″34°46′45″902019-0914HZY1~14* 广东广东药科大学(原种为棱枝何首乌)113°25′12″23°03′36″−2019-11 1LZ1*
*为引种样品,未加*为农户种植样品,“−”代表无海拔数据,下表同
The introduction samples were marked with “*”, while the farmers planting samples were not marked with “*”, in addition “−” means no elevation data
表2 何首乌野生品来源
Table 2 Origin of wild P. multiflorum samples
产地地点经度(E)纬度(N)海拔/m时间数量/份样品编号 广东德庆官圩金光村111°49′45″23°18′18″−2017-09 1JGC1 德庆官圩五福村111°47′39″23°15′31″ 402017-09 2WFC1~2 德庆官圩金林电站111°24′40″23°17′67″ 802017-09 1DZ1 德庆官圩酒厂111°23′23°16′−2017-09 1JC1 高要河台112°23°−2017-09 2HT1~2 高州石鼓镇白泥塘村110°47′47″21°49′03″ 202017-08 1BNT1* 肇庆鼎湖山112°33′03″23°9′42″−2017-10 1DHS1 德庆莫村镇太宪村112°07′33″23°24′35″ 602017-07 2TX1~2* 云浮市大方镇增西村111°34′37″22°56′46″ 962017-06 1ZXC1 茂名110°21°−2019-04 1MM1 重庆北碚区金刚碑106°24′52″29°50′23″−2017-10 5JGB1~5 北碚区西南大学106°25′55″29°49′41″−2017-10 6XNU1~6 南岸区五公里106°34′48″29°30′36″−2017-10 1WGL1 南川区重庆市药用植物园107°12′36″29°07′48″−2019-0116ZWY1~16 四川巴中市南龛山106°45′42″31°50′44″−2017-10 2NKS1~2 巴中市平昌元山镇107°10′29″31°43′13″−2017-10 1YSZ1 简阳市平泉镇104°38′52″30°20′47″−2017 1PQZ1 乐山市峨眉山103°20′20″29°36′24″10352018-05 1EMS1 103°20′03″29°36′13″ 9532018-05 1EMS2 成都市武侯区林荫街104°04′07″30°38′18″ 5192019-0410CD1~10 贵州贵阳市宝福山106°45′05″26°34′37″−2019-04 7BFS1~7 贵阳市小河106°26°32′21″−2019-04 9XH1~9 贵阳市花溪106°26°23′04″−2019-04 3HX1~3 湖南张家界贺龙公园110°29′33″29°22′53″−2018-08 1HLP1 南岳区衡山112°42′48″27°16’47″ 3602019-04 7HS1~7 112°41′40″27°17′36″11602019-04 2HS8~9 南岳树木园112°41′38″27°17’47″12302019-04 3SMY1~3 江苏宜兴市119°34′42″31°10′36″ 902018-05 1YX1 119°33′43″31°10′35″ 902018-05 1YX2 119°33′46″31°10′27″ 902018-05 3YX3~5 安徽广德县119°35′49″31°05′50″ 902018-05 2GD1~2 119°35′45″31°05′44″ 902018-05 1GD3 119°35′59″31°06′02″ 902018-05 1GD4 119°36′04″31°05′40″ 902018-05 1GD5 119°36′16″31°05′41″ 1002018-05 2GD6~7 119°36′17″31°05′42″ 1002018-05 2GD8~9 黄山市浮溪村118°08′30″30°05′12″−2018-08 1FXC1 118°08′57″30°04′44″−2018-08 5FXC2~6
何首乌SRAP-PCR产物通过6%非变性聚丙烯酰胺凝胶电泳和银染法进行条带显示。
1.3 数据统计
使用条带识别软件Quantity-One加人工识别的方式统计非变性聚丙烯酰胺凝胶电泳图上各泳道的条带迁移距离,相同位置有条带则记为“1”,无则为“0”,并建立二维数据矩阵。利用NTSYSpc、Popgen 32、MEGA 7和Structure 2.3.4软件对数据进行统计,获得何首乌种质的遗传多样性参数、遗传距离、样品聚类和分类结果。
2 结果及分析
2.1 13对引物在何首乌种质研究中的多态性分析
如表3所示,13对SRAP引物扩增位点数范围为17~31个,总位点数为318个,其中多态位点数为313个,所有引物均表现出极高的多态性,其中有9对引物多态性百分数为100%,结果表明13对引物扩增条带及引物多态性丰富,SRAP分子标记方法适用于何首乌种质的遗传多样性研究。
2.2 何首乌种质遗传多样性分析
何首乌种质遗传多样性、居群分化及基因交流程度结果如表4所示。野生品遗传多样性大于栽培品,栽培品中的农户种植样品,其奈氏基因多样性()和香农信息指数()均小于0.01,且多态位点百分比小于5%,表明以上居群遗传多样性较少,居群内样品整齐度较高,而引种样品遗传多样性则相对较高;野生品种中,重庆野生居群和四川野生居群的和值分别大于0.2和0.3,其多态性位点百分数分别为71.1%和66.0%,说明2居群野生何首乌遗传多样性较丰富;而贵州、湖南和安徽野生居群遗传多样性则仅次于重庆和四川,其他省份何首乌野生居群遗传多样性则相对较低。此外,栽培品和野生品居群间基因多样性(t)均大于居群内多样性(s),说明种质多样性主要来自居群间。整个何首乌种质资源的基因差异分化系数(st)为0.753 1(>0.500),表明地方群体间遗传分化程度大,但居群间的基因交流值(m)为0.163 9(<1.000)说明居群间的基因交流较少。另外栽培品的分化程度极高,但基因交流则极小,推测是由于栽培过程主要采用营养繁殖造成的。
表3 13对SRAP引物扩增结果
Table 3 Amplified results of 13 SRAP primers combinations
引物对总位点数多态位点数多态性百分数/% ME2EM9 24 24100.0 ME2EM13 23 23100.0 ME2EM18 19 19100.0 ME2EM20 25 25100.0 ME4EM11 24 24100.0 ME6EM8 22 21 95.5 ME8EM3 25 23 92.0 ME8EM4 26 26100.0 ME15EM15 17 17100.0 ME17EM2 26 25 96.2 ME17EM9 29 28 96.6 ME17EM13 31 31100.0 ME18EM8 27 27100.0 13对引物318313 98.4 平均值24.524.1 98.4
2.3 何首乌遗传距离和遗传一致度
居群间的遗传距离和遗传一致度结果见表5。栽培品中,除德庆大桥村CNT居群外,农户种植品居群间遗传一致度均大于0.89,说明农户栽培品种存在相互引种的情况。据调查,CNT何首乌栽培方法与德庆其他地方差异较大,且种苗一般为自留苗,推测这可能是导致遗传距离较大的原因,其块根形态与其它地区显示出明显差异,提示该种源的特异性。同为上海的2个栽培品YYL和YD间遗传距离较小,而棱枝何首乌则与其他所有居群的相似度均极小,遗传相似系数约为0.5;野生品中,广东野生居群和农户栽培品的相似度极高,而贵州的野生居群和其他居群的遗传相似性均相对较低,其值在0.6~0.8,但与四川居群的相似性最大,遗传相似系数为0.804 6。
2.4 何首乌遗传距离和地理距离的相关性
利用样品的经纬度信息,代入2点间地理距离公式,如公式1所示,式中1和2分别表示地方1和地方2的纬度,而1和2则表示地方1和地方2的经度,单位为米,如图1所示,何首乌样品间地理距离和遗传距离间的相关性极低,相关系数值仅为7.326×10−4,说明遗传距离与地理距离无明显相关性。
(1)
表4 何首乌居群遗传多样性
Table 4 Genetic diversity of P. multiflorum populations
居群名居群个体数NaNeHI多态位点数多态位点百分数/%HtHsGstNm STJ181.006 31.002 00.001 30.002 1 2 0.63 DT201.003 11.001 00.000 70.001 2 1 0.31 CNT201.012 61.006 70.004 40.006 7 4 1.26 TX201.003 11.000 70.000 60.001 1 1 0.31 BNT201.009 41.001 00.000 90.001 9 3 0.94 GG261.034 61.007 20.004 90.008 411 3.46 SKC201.000 01.000 00.000 00.000 0 0 0.00 SZY 91.242 11.167 00.093 90.137 57724.21 JZY 91.289 31.182 90.105 00.155 89228.93 YY111.207 51.135 80.076 70.113 36620.75 HC 81.072 31.049 10.027 70.040 623 7.23 SXZ201.103 81.040 20.025 90.040 83310.38 HZY141.289 31.167 90.097 40.146 19228.93 YYL 11.000 01.000 00.000 00.000 0 0 0.00 YD 11.000 01.000 00.000 00.000 0 0 0.00 LZ 11.000 01.000 00.000 00.000 0 0 0.00 广东野生居群131.317 61.184 50.107 90.161 810131.76 重庆野生居群281.710 71.399 90.229 60.344 422671.07 四川野生居群161.660 41.357 50.205 00.311 221066.04 贵州野生居群191.496 91.255 90.150 30.228 515849.69 湖南野生居群131.449 71.232 00.136 10.207 314344.97 江苏野生居群 51.242 11.160 80.091 50.135 0 7724.21 安徽野生居群151.503 11.273 60.162 10.245 416050.30 栽培品2181.896 21.369 80.223 40.348 221889.620.242 40.027 50.886 70.063 9 野生品1091.956 01.477 00.285 40.436 330495.600.276 30.154 70.440 20.635 8 全部样品3271.984 31.454 90.271 70.417 931398.430.268 00.066 20.753 10.163 9
表5 何首乌居群间遗传距离和遗传一致度
Table 5 Genetic distance and genetic consistency between P. multiflorum populations
居群名STJDTCNTTXBNTGGSKCSZYJZYYYHCSXZHZYYYLYDLZ广东野生居群重庆野生居群四川野生居群贵州野生居群湖南野生居群江苏野生居群安徽野生居群 STJ****0.941 50.776 30.906 90.973 00.965 80.995 00.782 60.830 10.775 10.753 90.737 60.750 10.774 80.734 00.534 30.927 70.812 90.791 90.653 90.811 70.779 20.763 4 DT0.060 3****0.765 40.896 90.937 00.929 80.940 20.764 90.833 00.771 20.730 20.728 50.758 70.757 70.730 30.524 90.913 80.813 00.785 90.662 20.793 00.798 20.790 7 CNT0.253 20.267 4****0.830 20.791 00.783 20.781 30.783 60.849 90.789 30.739 40.716 00.723 70.779 90.762 70.491 30.821 90.792 50.783 60.608 10.785 80.758 40.739 9 TX0.097 80.108 80.186 0****0.927 90.920 30.911 90.754 30.830 30.759 30.769 50.692 70.728 70.767 20.732 60.496 70.887 80.788 50.754 20.603 30.773 80.758 30.738 6 BNT0.027 40.065 10.234 50.074 8****0.993 20.978 00.775 20.828 50.758 60.762 70.722 00.741 80.770 60.723 40.519 00.917 40.805 10.779 30.631 30.794 40.778 00.765 2 GG0.034 80.072 80.244 30.083 00.006 8****0.970 80.767 80.820 80.754 40.759 00.725 40.734 40.763 70.722 90.518 60.912 50.802 30.774 70.627 90.788 60.770 20.757 6 SKC0.005 00.061 70.246 80.092 20.022 30.029 6****0.781 70.829 40.774 70.759 10.731 40.748 90.773 60.732 70.534 60.928 10.812 10.788 50.649 10.809 30.777 40.763 9 SZY0.245 10.268 00.243 80.282 00.254 70.264 20.246 3****0.837 40.864 50.798 80.780 80.756 20.899 10.815 90.572 20.849 80.880 90.852 50.749 90.846 90.840 30.850 7 JZY0.186 20.182 70.162 60.186 00.188 20.197 50.187 10.177 4****0.827 90.788 40.751 60.780 80.811 60.787 00.556 00.890 90.869 50.831 90.676 40.846 60.801 40.820 7 YY0.254 80.259 80.236 60.275 30.276 30.281 80.255 30.145 60.188 9****0.797 90.821 60.778 60.815 20.960 90.575 50.844 20.882 90.867 60.699 40.856 50.867 40.882 0 HC0.282 50.314 40.302 00.262 00.270 90.275 80.275 60.224 70.237 70.225 8****0.717 80.727 70.780 90.773 70.540 40.779 00.804 00.779 80.640 30.768 10.803 20.759 5 SXZ0.304 40.316 70.334 00.367 20.325 70.321 00.312 80.247 50.285 60.196 50.331 6****0.750 90.745 40.775 30.576 30.779 40.812 40.857 60.680 00.777 60.797 60.824 8 HZY0.287 60.276 20.323 30.316 50.298 60.308 70.289 20.279 50.247 50.250 30.317 90.286 5****0.746 20.733 00.604 30.801 90.819 90.826 90.707 50.799 10.775 30.801 8 YYL0.255 20.277 40.248 60.265 00.260 50.269 60.256 70.106 40.208 70.204 30.247 30.293 90.292 7****0.783 00.540 90.832 40.837 40.810 10.698 70.815 20.801 50.810 6 YD0.309 20.314 40.270 90.311 20.323 70.324 40.311 00.203 40.239 50.039 90.256 60.254 60.310 60.244 6****0.544 00.800 70.827 10.802 00.648 30.808 70.835 70.833 9 LZ0.626 80.644 50.710 60.699 80.655 90.656 60.626 30.558 30.587 00.552 60.615 50.551 10.503 70.614 60.608 8****0.569 60.627 60.642 10.631 50.589 50.598 10.642 0 广东野生居群0.075 10.090 10.196 10.119 00.086 20.091 60.074 60.162 70.115 50.169 30.249 70.249 20.220 80.183 50.222 30.562 8****0.880 50.863 00.714 10.857 60.840 70.843 8 重庆野生居群0.207 20.207 10.232 50.237 60.216 70.220 20.208 10.126 90.139 80.124 50.218 10.207 80.198 50.177 50.189 80.465 80.127 3****0.911 60.795 40.881 30.855 90.892 6 四川野生居群0.233 40.240 90.243 80.282 10.249 40.255 20.237 70.159 60.184 10.142 10.248 70.153 70.190 10.210 60.220 60.443 00.147 30.092 6****0.804 60.867 20.864 50.890 7 贵州野生居群0.424 90.412 20.497 40.505 30.459 90.465 30.432 20.287 80.391 00.357 50.445 90.385 70.346 00.358 60.433 40.459 70.336 80.228 90.217 4****0.764 30.702 00.785 0 湖南野生居群0.208 60.232 00.241 10.256 50.230 20.237 50.211 50.166 20.166 60.154 90.263 90.251 50.224 20.204 30.212 30.528 40.153 60.126 40.142 50.268 7****0.848 40.876 8 江苏野生居群0.249 50.225 40.276 60.276 70.251 00.261 10.251 80.174 10.221 50.142 30.219 10.226 20.254 50.221 30.179 50.514 10.173 50.155 60.145 70.353 90.164 4****0.887 0 安徽野生居群0.270 00.234 80.301 30.302 90.267 60.277 60.269 30.161 70.197 60.125 50.275 10.192 70.220 90.210 00.181 60.443 20.169 80.113 60.115 70.242 10.131 40.1199****
“****”上方为居群间的遗传一致度,“****”下方为居群间的遗传距离
The top of “****” in the table is the genetic consistency between populations, and the bottom of “****” is the genetic distance between populations

图1 何首乌样品间遗传距离和地理距离的相关性
2.5 何首乌样品的聚类分析和主成分分析
采用邻接法(neighbor joining,NJ)构建何首乌样品的聚类树,结果如图2所示。何首乌样品主要分成4大分支,第1分支为棱枝何首乌,它与其它种源表现出明显的不同,因此独立成支;第2分支包括四川的EMS1-2和贵州的所有野生品,而河南郑州的样品则聚类为第3支,其余的所有样品组成第4分支。第4分支中,农户种植的栽培品相似度较高,聚为一小支,而广东野生品则分散聚类在该小支中,此外,除重庆和四川的样品聚类结果较为分散外,其余采自相同省份和相同地点的样品基本能聚在一起,说明重庆和四川的野生品遗传多样性较大,样品间存在较大差异。
样品主成分分析分布图见图3,图中样品距离越小代表样品间的遗传关系越接近,反之则说明亲缘关系越远。利用主成分分析对样品进行分类,主要可以把何首乌样品分成3个类群,分类结果和NJ聚类结果相似,棱枝何首乌单独作为一个类群,第2类群为贵州省的所有样品和四川峨眉山的2个样品,而第3类群则由其他样品组成。
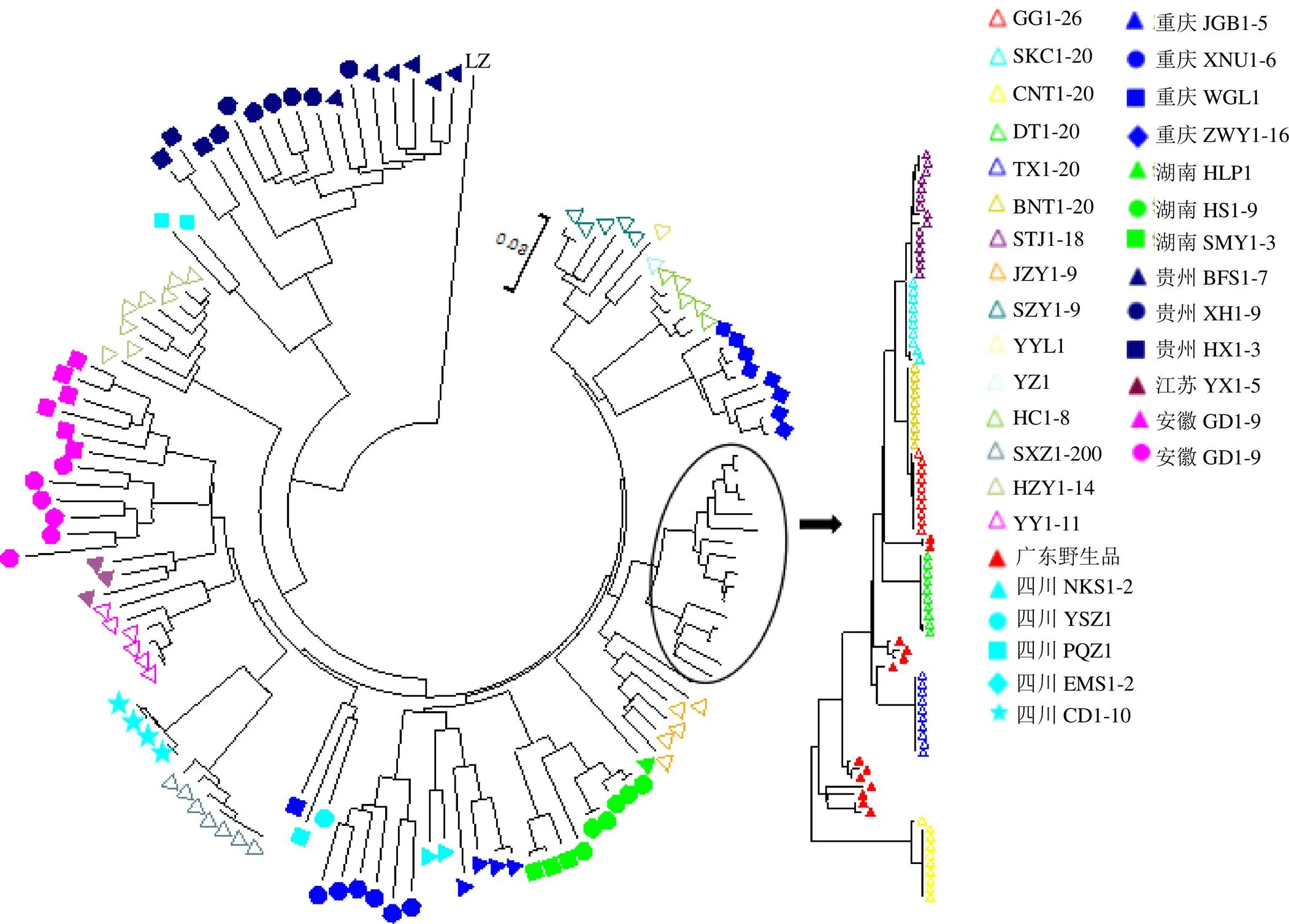
几何形状代表何首乌样品,空心图形为栽培品,实心为野生品,其中填充颜色相同即代表该野生何首乌来自相同的省份
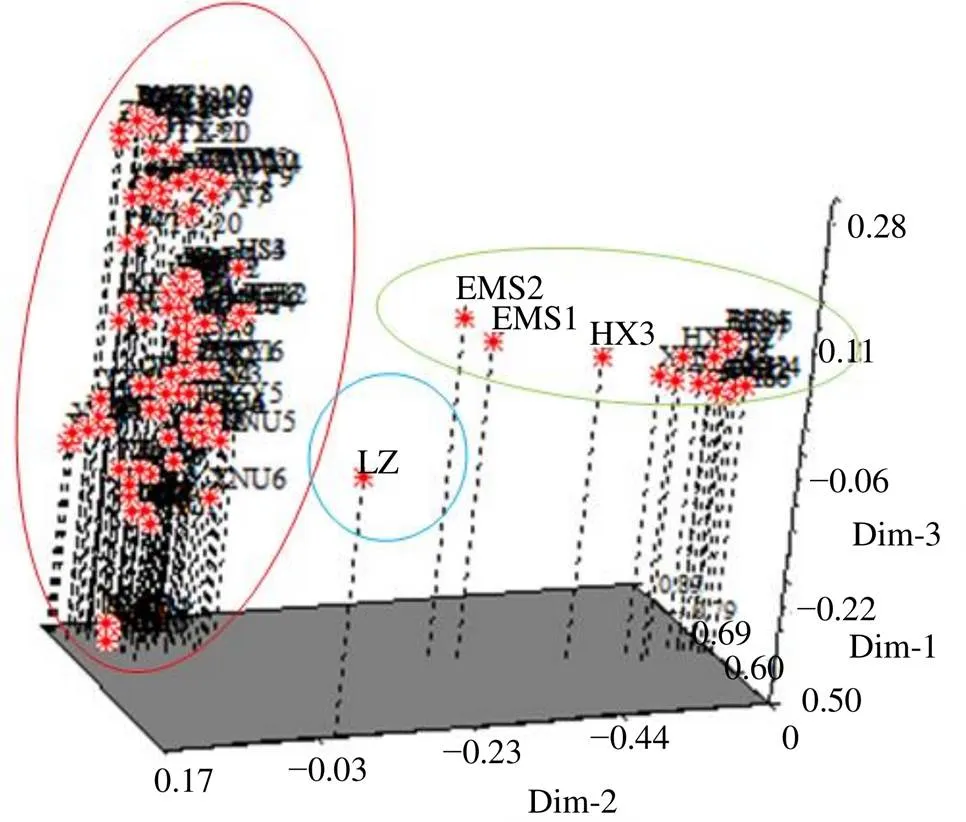
图3 何首乌样品主成分分析
2.6 何首乌群体结构分析
利用Structure 2.3.4软件对何首乌种质进行群体结构分析,假定群体数目值为1~23,每个值重复运算10次,根据Evanno等[14]的方法计算出Δ值,如图4所示,结果表明当=16时,其Δ值最大,说明种群分成16个类群时最佳,并由此获得群体结构图,如图5所示,不同颜色代表不同类群,每个长方形表示一个样品,纵坐标为各类群种质占其祖先成分的比例,根据刘丽华等[15]的研究,单个样品某血统≥0.600说明该样品血统及遗传基础较单一,可归类到对应的类群中,<0.600则具有混合血缘,即遗传结构比较复杂且没有明确的类群归属特性,经样品归纳如表6所示,何首乌327份样品中,319份样品在某一类群中的≥0.600,说明何首乌群体中大部分种质的遗传结构较简单,而EMS(1~2)、HLP1、JGB1、DZ1、JGC1及HT(1~2)样品则血统相对复杂。此外结果还表明,相同采集点样品血缘组成相似,可大致归在同一类群中,其中除CNT血统较为特殊外,其余农户栽培品遗传基础相同均归为一类。和NJ聚类相似,群体结构的分类是NJ聚类4大分支基础上的细分和调整,不同的地方是在NJ聚类图中单独成支的棱枝何首乌(LZ)此处则与重庆和四川的野生品WGL1、PQZ1和YSZ1分在同一类。
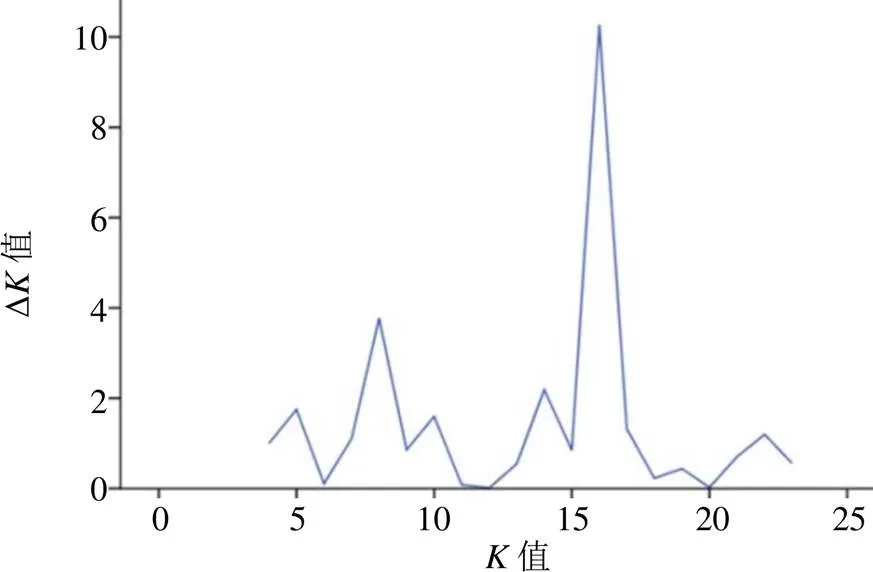
图4 ΔK值运算结果
3 讨论
3.1 SRAP分子标记方法在何首乌种质研究中的有效性
本实验结果表明,利用SRAP分子标记技术研究何首乌栽培和野生种群的遗传多样性,其扩增条带及多态性条带较丰富,且通过SRAP标记数据对种群进行划分,分类结果和种质来源相符,证明该方法能够反映何首乌种质遗传多样性和遗传差异的真实情况,目前相关研究也证明,SRAP分子标记是遗传多样性估计和种群关系的有效分析工具[16]。
3.2 何首乌居群遗传多样性
经种质遗传多样性分析,野生居群的多样性大于栽培居群,尽管引种品居群内及居群间样品存在一定的遗传多样性,但由于农户种植品种存在相互引种的情况,且居群内种源一致度较高,因此相较于野生居群,栽培品居群整体的多样性较低。而野生居群中,四川和重庆居群的遗传多样性较高,湖南、贵州和安徽居群次之,而广东和江苏居群相对较低。通过地形分析发现,四川、重庆位于我国西南地区,地处第1和第2阶梯中,地形海拔高、重峦叠嶂、复杂多变,湖南和安徽虽处于第3阶梯,然而两地样品采集地分别为衡山和黄山,海拔较高,而贵州的3个样品采集点也位于云贵高原中,其它采样点则处于南方丘陵地区和华东、华中平原地带,由此可见,地形复杂性和海拔可能是影响遗传多样性程度的重要因素。也有研究发现利用ISSR[8]技术,野生和栽培何首乌的多样性无明显差异,经核对文献,本课题组认为样品采集数量和采样点分布范围及研究方法的不同造成了结果的不一致。此外,基因交流是减少遗传漂变的重要途径,然而何首乌为自交物种[10],因此居群基因交流少,而遗传分化程度高。
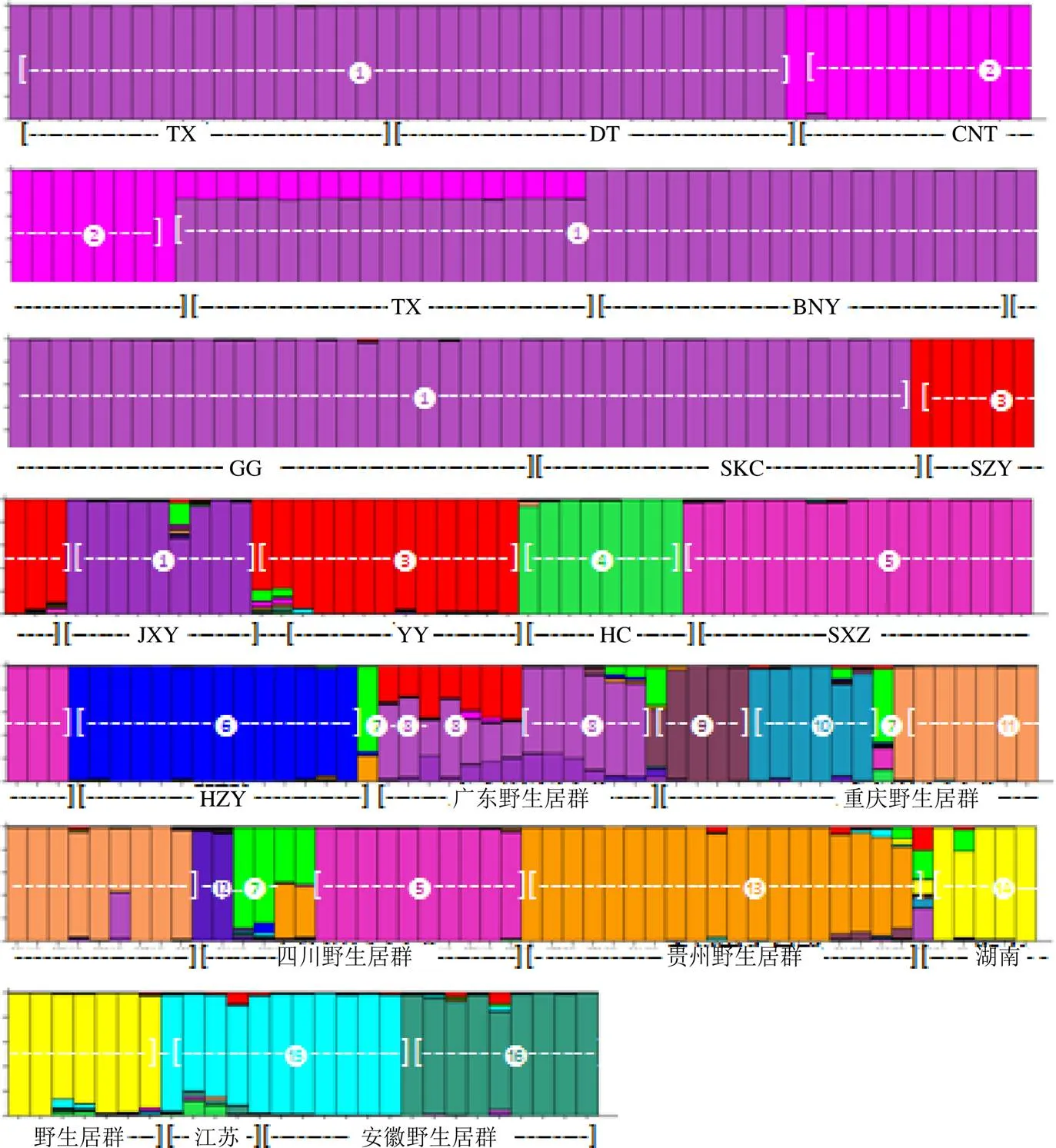
图5 何首乌群体结构
表6 何首乌样品分类结果
Table 6 Classification results of P. multiflorum samples
Q值组别样品编号 ≥0.6001TX(1~20)、DT(1~20)、BNT(1~20)、STJ(1~18)、GG(1~26)、SKC(1~20)、JZY(1~9) 2CNT(1~20) 3SZY(1~9)、YD1、YYL1、YY(1~11) 4HC(1~8) 5SXZ(1~20)、CD(1~10) 6HZY(1~14) 7LZ1、WGL1、PQZ1、YSZ1 8WFC(1~2)、JC1、DHS1、BNT*1、ZXC1、MM1、TX*(1~2) 9JGB(2~5) 10XNU(1~6) 11ZWY(1~16) 12NKS(1~2) 13BFS(1~7)、XH(1~9)、HX(1~3) 14HS(1~9)、SMY(1~3) 15YX(1~5)、FXC(1~6) 16GD(1~9) <0.600EMS(1~2)、HLP1、JGB1、DZ1、JGC1、HT(1~2)
3.3 遗传距离和地理距离的关系
研究结果表明何首乌样品间遗传距离与地理距离无明显相关性。首先栽培品存在相互引种的情况,再者西南地区地形复杂,居群多样性高,尽管西南地区内部采样点相距较近,但地形差异造成的遗传分化致使遗传距离相对较大,2个原因共同导致两者不呈明显的相关性。
3.4 NJ聚类结果和STRUCTURE居群结构分析结果及比较
NJ聚类和STRUCTURE群体分析是种质亲缘关系研究及样品分类的方法,相较于NJ聚类,STRUCTURE既可以研究每个样品的血统构成,还可以明确种群的最佳类群数,而聚类分析则是根据样品间的差异程度进行分类,类群数一般为人为所定,缺乏一定的科学性,不过聚类图能清楚呈现类群间的亲缘关系,因此结合2种方法能更深入地探究种质的群体结构。
利用遗传距离构建的聚类结果和以STRUCTURE软件获得的居群结构分析结果对种源的分类基本一致,聚类特点与已有研究相似,即聚类结果与地理分布相符,相同采集点样品可分在同一类中[8-10,17-19]。群体结构分析显示,大部分何首乌样品的血缘组成较单一,相同采集点样品血缘组成相似,可大致归在同一类群中,其中除CNT血缘较为特殊外,其余农户栽培品遗传基础相同均归为一类。值得关注的结果是农户种植品种和广东野生品种聚为一支,说明样品间亲缘关系较接近,从侧面可反映目前农户栽培品可能由广东野生品种驯化而来,并且广东野生品间并未出现明显分化。另外,四川EMS和贵阳的样品与其它样品遗传距离大,独立成支,有研究人员在何首乌种质研究中也发现,贵州居群叶片侧叶脉有较大程度的白斑,同时该居群与其它居群遗传距离较大,基因交流值仅为0.01[8],而本实验也同样发现贵阳的居群叶脉有较大的白斑,可能是样品位于云贵高原的特殊地形所致。同时,棱枝何首乌在何首乌种群中表现出极大的特异性,与其他种源遗传距离大,支持将其列为何首乌的变种。
上述结果可以把何首乌种质特点总结为:由于较快的遗传进化速度,为适应多变的环境,何首乌形成不同的地理种源,相同环境下何首乌遗传距离小,相似度较高,且血缘组成相同,而地理距离较近,但不同地形环境的何首乌存在差异,因此地形的复杂性和海拔是何首乌种群遗传多样性的重要影响因素。由于何首乌为自交的物种,基因交流少,因此地理环境所造成的居群遗传分化未能通过基因交流而得到减少,所以何首乌居群间的遗传分化较大,最终导致何首乌大部分样品的血统组成单一,但是不同生长环境居群的血缘组成不同的现象。
利益冲突 所有作者均声明不存在利益冲突
[1] 杨丽. 中药学 [M]. 第2版. 北京: 人民卫生出版社, 2010: 28.
[2] 黄志海, 徐文, 张靖, 等. 中药何首乌全球生态适宜性分析 [J]. 世界中医药, 2017, 12(5): 982-985.
[3] 刘海华, 艾伦强, 唐春梓, 等. 何首乌野生资源调查及存在的问题 [J]. 湖北农业科学, 2012, 51(24): 5718-5721.
[4] 黄和平, 王键, 黄璐琦, 等. 何首乌资源现状及保护对策 [J]. 海峡药学, 2013, 25(1): 40-42.
[5] 史顺敏, 俞年军, 于凡, 等. 丹参种质资源的研究进展 [J]. 现代中医药, 2016, 36(6): 113-117.
[6] 徐海明. 种质资源核心库构建方法的研究及其应用 [D]. 杭州: 浙江大学, 2005.
[7] 陈红波, 余金芮, 杨春先, 等. 药用植物研究中的分子标记技术应用进展 [J]. 湖北农业科学, 2017, 56(13): 2401-2405.
[8] 方清茂. 栽培与野生何首乌种质资源遗传多样性的ISSR分析[A] // 中药饮片质量分析与中药鉴别技术交流研讨会论文集[C]. 北京: 北京晟勋炎国际会议服务中心, 2009: 45-51.
[9] 王凌晖, 曹福亮, 汪贵斌, 等. 何首乌野生种质资源的RAPD指纹图谱构建 [J]. 南京林业大学学报: 自然科学版, 2005, 29(4): 37-40.
[10] 程远辉. 基于SRAP分析的重庆何首乌种质资源整理研究 [D]. 重庆: 西南大学, 2007.
[11] Li G, Quiros C F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in[J]., 2001, 103(2/3): 455-461.
[12] Robarts D W H, Wolfe A D. Sequence-related amplified polymorphism (SRAP) markers: A potential resource for studies in plant molecular biology (1.) [J]., 2014, 2(7): 1400017.
[13] Jia S Z, Yan Z M, Wang Y H,. Genetic diversity and relatedness among ornamental purslane (L.) accessions unraveled by SRAP markers [J]., 2017, 7(4): 241.
[14] Evanno G, Regnaut S, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study [J]., 2005, 14(8): 2611-2620.
[15] 刘丽华, 王立新, 赵昌平, 等. 光温敏二系杂交小麦恢复系遗传多样性和群体结构分析 [J]. 中国生物化学与分子生物学报, 2009, 25(9): 867-875.
[16] Zhang R P, Wu J, Li X G,. An AFLP, SRAP, and SSR genetic linkage map and identification of QTLs for fruit traits in pear (L.) [J]., 2013, 31(3): 678-687.
[17] 张宏意, 袁林林, 饶秋红, 等. 不同产地何首乌叶绿体psbA-trnH基因序列分析 [J]. 中草药, 2018, 49(5): 1146-1149.
[18] 白明明, 孙小芹, 郭建林, 等. 基于psbA-trnH分析的何首乌野生居群遗传多样性 [J]. 植物资源与环境学报, 2012, 21(2): 36-44.
[19] 严寒静, 房志坚, 余世孝. 不同种源何首乌的ITS序列分析及其亲缘关系研究 [J]. 西北植物学报, 2008, 28(5): 922-927.
Genetic diversity and population structure ofgermplasm based on SRAP molecular marker
LI Jia-hui1, OU Xiao-hua1, LIU Xiao-ying1, LU Chang-hua1, ZHANG Hong-yi1,2, HE Meng-ling1,2, YAN Han-jing1,2
1.School of Traditional Chinese medicine, Guangdong Pharmaceutical University, Guangzhou 510006, China 2. Key Laboratory of Production & Development of Cantons Medicinal Materials, National Administration of Traditional Chinese Medicine, Guangzhou 510006, China
Toinvestigate the genetic diversity and population structure ofgermplasm, in order to provide reference for the protection and rational utilization ofgermplasm resources.Base on SRAP molecular marker method, the diversity level, genetic distance, genetic differentiation degree, clustering results and population analysis results ofpopulation were obtained by using Structure 2.3.4, NTSYSpc, Popgen 32 and MEGA 7 software to make statistics on the data.The genetic diversity ofwas mainly from intergroup, and the genetic diversity of wild population was greater than that of cultivated population. In the wild population, the genetic diversity was high in Sichuan and Chongqing population, and the genetic distance between samples ofhad no obvious correlation with geographical distance; The NJ clustering results of the samples were consistent with the geographical distribution, and the samples from the same collection point could be clustered together. In addition, the results also showed that the cultivars of peasant households were closely related, indicating the existence of mutual introduction between the cultivars. Population structure analysis showed that most samples had single consanguinity, and the best classification result which was consistent with NJ clustering result could be obtained when= 16 was used.SRAP molecular marker technique is an effective analysis tool for genetic diversity estimation and population relationship studies of. The complexity of terrain may be an important factor affecting the genetic diversity of. In addition,var.from Guangxi shows specificity inpopulation and has a large genetic distance from other provenances, therefore, it is suggested to classify it as a variety of.
Thunb.; SRAP; genetic diversity; germplasm
R286.12
A
0253 - 2670(2022)07 - 2115 - 11
10.7501/j.issn.0253-2670.2022.07.022
2021-08-09
广东省科技厅项目(2017A020213023)
李嘉惠(1994—),女(汉族),广东肇庆人,硕士研究生,主要从事中药资源开发与品质评价研究工作。E-mail: 787517019@qq.com
严寒静(1972—),女(汉族),重庆人,硕士研究生导师,博士学位,主要从事中药资源开发与品质评价研究工作。E-mail: yanhanjing1211@163.com
[责任编辑 时圣明]
