青秆竹花的解剖观察与分析
2022-03-17龙昊初彩华金点坤吕卓王曙光
龙昊 初彩华 金点坤 吕卓 王曙光


摘 要: 為明确自然状态下青秆竹(Bambusa tuldoides)不同发育阶段花器官的形态以及雌雄配子体的发育状态,总结其败育类型,该文通过采用解剖和切片等方法对青秆竹花器官的各部分外观形态以及雌雄配子体的发育过程进行观察,并分析其结实率低下的原因。结果表明:(1)青秆竹小穗为无限花序,下部的小花先发育,但基部具有潜伏芽,因此又具有有限花序的特征;小穗柄不发达,簇生花枝节部。(2)每朵小花拥有内、外稃各1枚,花药6枚,浆片3枚,雌蕊1枚;浆片透明,边缘具有发达的纤毛;子房具棱,子房上部具绒毛,子房1室,侧膜胎座,倒生胚珠,三分枝羽状柱头。(3)青秆竹花药具有4个药室,花药壁由表皮、药室内壁、中层、绒毡层4层结构组成;绒毡层为腺质型,花药发育后期极度退化;小孢子母细胞分裂类型为连续型,产生两边对称型小孢子,花粉粒细胞成熟后为3核。(4)雄蕊和雌蕊出现多种败育类型,可能是导致结实率低的主要原因。综上结果表明,青秆竹花器官的形态结构发育正常,而雌雄配子体发育过程中出现异常,造成了其结实率低。
关键词: 青秆竹, 雄蕊, 雌蕊, 败育
中图分类号: Q945
文献标识码: A
文章编号: 1000-3142(2022)01-0174-09
收稿日期: 2021-10-10
基金项目: 国家自然科学基金(32060379) [Supported by the National Natural Science Foundation of China(32060379)]。
第一作者: 龙昊(1995-),硕士研究生,主要从事竹类植物研究,(E-mail)1024170498@qq.com。
*通信作者: 王曙光,博士,教授,主要从事竹类植物研究,(E-mail)stevenwang1979@126.com。
Anatomical observation and analysis on
floral of Bambusa tuldoides
LONG Hao, CHU Caihua, JIN Diankun, L Zhuo, WANG Shuguang*
( College of Life Sciences, Southwest Forestry University, Kunming 650224, China )
Abstract: In order to clarify the morphology of floral organs and the development status of male and female gametophytes at different development stages of Bambusa tuldoides in wild, and to summarize their abortion types, the appearance morphology of each part of floral organs and the development process of male and female gametophytes of B. tuldoides were observed by means of anatomy and sectioning, and the reasons for the low seed-setting rate were also analyzed. The results were as follow: (1) The spikelets of B. tuldoides were of infinite inflorescences. The florets in the lower part of the spikelets developed first, and there were latent buds developed at the base, which had the characteristics of definite inflorescence; The petioles of spikelets were undeveloped, and the spikelets developed in cluster at each node of the floral branch. (2) Each floret had 1 palea, 1 lemma, 6 stamens, 3 lodicules and 1 pistil; The lodicule was transparent and had well-developed cilia on the edge; The ovary had prismatic protrusions and the upper part of which had villi, and was unilocular and lateral membranous with a anatropous ovule. The stigmas were three-lobed and feathery. (3) The anther of B. tuldoides had four sporangia, and the anther wall was composed of four layers, epidermis, endothecium, middle layer and tapetum; The tapetum was glandular, and degenerated completely after maturity; The meiosis was successive and the microspore tetrads were bilaterally, and the mature pollen grains were three-celled.(4)Several abortion types were found in pistils and stamens, which might be the main reasons for the low seed-setting rate of B. tuldoides. All the above results indicate that the morphological structure of B. tuldoides floral organs is normal, while the female and male gametophytes developed abnormally, which cause the low seed setting rate.
Key words: Bambusa tuldoides, stamen, pistils, abortion
竹类植物开花周期很长,经过营养生长阶段进入生殖生长阶段大约需要60年或更长的时间,竹子开花现象往往伴随着竹子的死亡,竹子开花現象十分罕见,并且难以预测其开花周期(林树燕等,2010),而且竹类植物的结实率也十分低下,尤其是一些栽培种(杜凡等,2000),竹类植物的这一特点导致竹类植物的生殖器官各方面的研究进展十分缓慢。随着被发现的开花的竹子种类增多,国内对竹子花器官的形态与解剖学研究相关的报道也在逐渐增加,大多是花器官的解剖形态观察与雌雄配子体发育的研究,如雷竹(Phyllostachys praecox)、巨龙竹(Dendrocalamus sinicus)、鹅毛竹(Shibataea chinensis)、硬头黄竹(Bambusa rigida)、新小竹(Neomicrocalamus prainii)、‘霞早’绿竹 (Bambusa oldhami ‘Xia Zao’ ZSX)、孝顺竹(B. multiplex)等竹种(黄坚钦等,1999;王曙光等,2006;林树燕和丁雨龙,2012;林树燕等,2015,2019;初彩华等,2019;李娟等,2020)。
青秆竹(Bambusa tuldoides)为箣竹属(Bambusa)竹种,产于广东、香港、广西以及贵州南部,福建、云南有引栽,秆供建筑、家具、农具等用材,秆表面刮下的竹茹可供药用(易同培等,2008)。目前国内仅有对青秆竹的竹茹(周滢,2014)及光合特性(郭雯等,2019)等方面的相关研究报道,国外关于青秆竹的研究大多与其秆材理化性质相关(Azzini et al., 1988;Sun et al., 2015)。关于花的研究仅有de Souza等(2020)首次对青秆竹发育成熟后的花药形态与解剖结构进行观察,认为花粉粒结构的不正常导致其败育,但该研究并未对青秆竹花器官整体的形态与解剖特征进行系统的描述,也缺乏对其雌雄配子体发育过程的详细观察与描述,因而总结出的败育原因也不全面,除此之外,并未发现其他关于青秆竹花器官的研究,所以目前还缺乏对自然生长状态下的青秆竹花器官各部位具体形态、胚胎发育及败育方面的系统性研究。本研究通过解剖及切片等手段,对青秆竹花器官进行形态与解剖观察,拟解决以下问题:(1)系统的描述青秆竹花器官的形态与解剖特征;(2)分析青秆竹花器官雌雄配子体发育过程;(3)探讨青秆竹自然状态下结实率低的原因。
1 材料与方法
实验材料于2013年8月24日取自西双版纳傣族自治州勐腊县(21°56′ E,101°15′ N),开花的竹丛并未出现即将死亡的迹象,并且竹丛全部为开花状态。通过对开花不同状态的观察,将各个发育阶段的青秆竹小花整个花枝剪下浸泡在FAA固定液中(50%乙醇∶甲醛∶乙酸=18∶1∶1),带回并通过真空泵抽气保存。
对固定保存后的花枝进行解剖,使用体式解剖镜(Olympus HO11)对花器官进行解剖,并对各个部位进行拍照、测量。再将分离出来的不同发育阶段的子房、花药进行脱水、石蜡包埋、切片,切片厚度为7~8 μm,番红固绿染色,中性树胶封片,观察。切片的步骤具体参照李正理等(1996)的石蜡切片方法,在光学显微镜下观察并使用二维测量软件 DS-3000拍照。
2 结果与分析
2.1 花序的结构
青秆竹的小穗簇生于花枝各节,每节一般有2至多个小穗,发育程度不一。通常靠近花枝底部的小穗先伸长展开,随着花枝伸长,以花枝为中心向四周开放(图版I :A)。位于花枝中上部小穗开放时间相对较晚,即使下方小穗凋零后上部的小穗也尚未展开。青秆竹小穗基部往往包裹着多枚黄灰色苞片,小穗形状稍扁,呈淡绿色。小穗基部不断通过产生新的花芽生长成新的小穗,而老的小穗逐渐脱落(图版I:A),这具有有限花序的特征。小穗脱落后残留的苞片仍然附着在花枝的各节上。小穗轴节间清晰可见,但小穗柄不发达(图版I:C)。青秆竹每个小穗具有6~7朵小花,靠近小穗上下两端的小花往往发育不正常,靠近小穗基部的花先发育(图版I:B),具有明显的无限花序特征。小穗轴顶宽底窄,节间扁平清楚可见并且宽度也不相同,呈杯状,被短毛(图版I:E)。每枚小穗基部的潜伏芽被1枚苞片包裹。苞片与小花的外稃形态类似(图版I:D)。
2.2 青秆竹小花形态结构
青秆竹每朵小花可分为内稃、外稃、雄蕊、浆片、雌蕊五部分(图版I:F)。青秆竹的小花含内、外稃各1枚,内稃形状相较于外稃偏扁平而且质地较软更易舒展,经测量青秆竹同一小花的外稃长度远大于内稃,这点与《中国竹类图志》记载的不同(易同培等,2008)。外稃内外表面光滑无毛,尖端较钝,有大约19脉(图版I:H)。 内稃对生于外稃内侧,具两脊,覆有稀疏且极短的纤毛(图版I:G)。雄蕊6枚,整体呈淡黄绿色,部分带有红色,先端较尖,基部分两尖(图版I:I)。花药成熟后,花药室纵裂散粉,与毛竹(孙立方等,2012)、鹅毛竹(林树燕和丁雨龙,2012)相似,雄蕊发生褐化(图版I:J)。浆片3枚,呈透明膜质,鱼鳞状,边缘具有较多较长的纤毛(图版I:K),围生于子房下部和内稃相邻,小穗成熟后,浆片通过吸水加厚撑开向外挤压内稃,同时撑开外稃,露出柱头和花药,完成传粉后浆片全部褐化(图版I:L),所以青秆竹花属于开放型花(林树燕等,2010)。雌蕊1枚,未成熟时子房呈乳白色,成熟子房呈淡黄绿色,柱头为三分支羽毛状,张文燕和马乃训(1990)根据柱头的长度及稃片开合情况将子房分为长花柱型与短花柱型,青秆竹花的子房花柱较短,柱头明显长于花柱,为长花柱型。子房倒卵形,下部有明显的突起的棱,底部无毛,越靠近花柱绒毛越长,未成熟的子房上部同样被有绒毛,但成熟子房的绒毛更长更明显(图版I:M-N)。
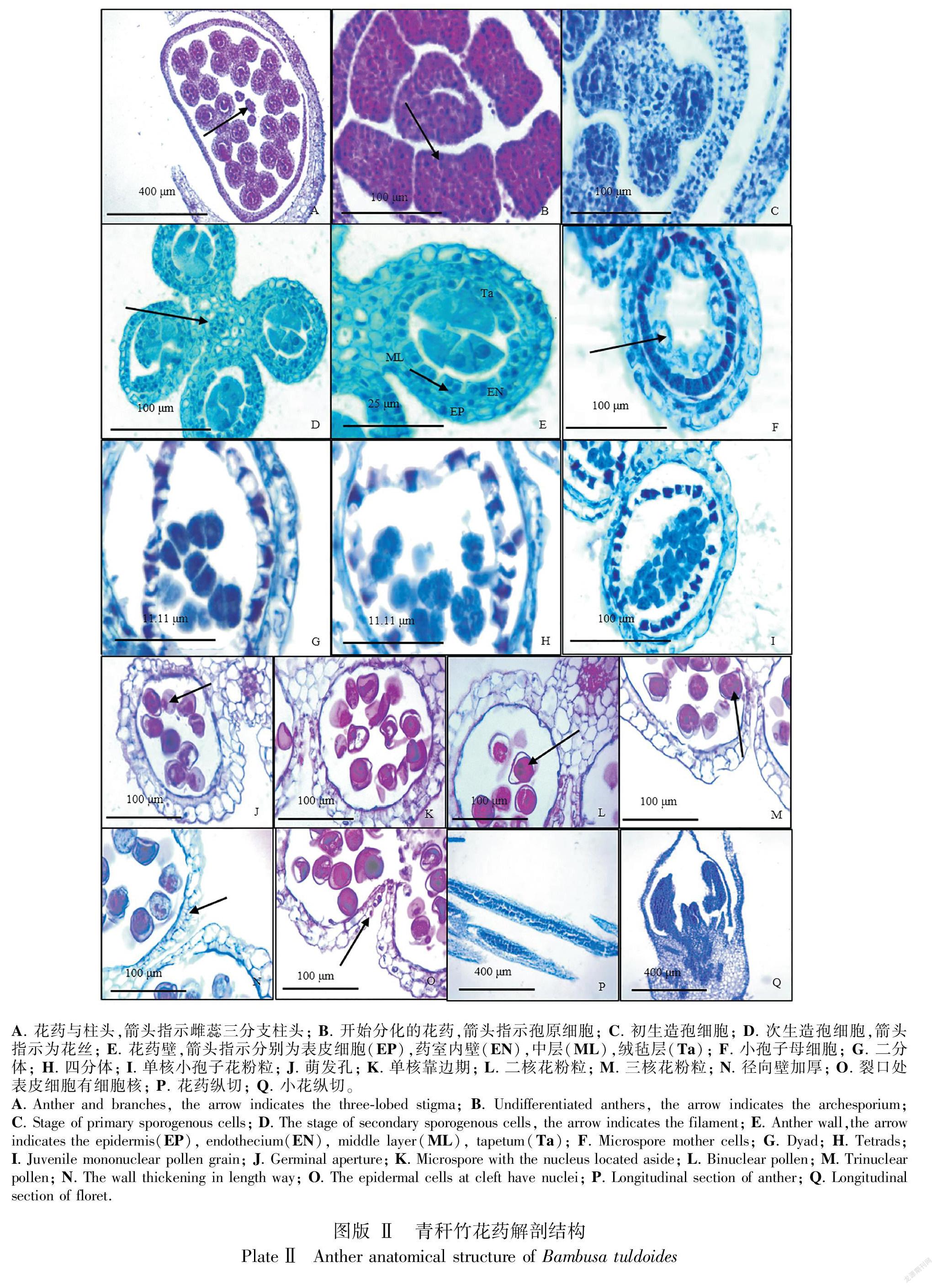
2.3 青秆竹花药形态结构与发育
青秆竹的花药属于基着药,4药室,两两对称,纵裂散粉。通过对青秆竹小花花药进行解剖发现,花药每个药室的横切面形状呈椭圆形,6枚雄蕊将雌蕊包围在中间(图版Ⅱ:A),花药壁的层数随着花药发育时期的不同而发生相应的变化。花药刚开始分化时,药隔组织将花药分为4个药室,在药室内部会分化出孢原细胞,孢原细胞与薄壁细胞相比,通常具有更加显著的细胞核(图版Ⅱ:B)。随后进入初生造孢细胞时期,这是孢原细胞已经通过有丝分裂成为初生造孢细胞(图版Ⅱ:C)。初生造孢细胞时期,药室内造孢细胞数量相对较少,此时期花药壁开始出现分化,但并未出现明显的层与层之间的界限,但细胞层已初见雏形。次生造孢时期与初生造孢细胞时期相比,造孢细胞数量增加,同时花药壁出现明显的分化,由4层细胞组成,由内到外依次是绒毡层(Ta)、中层(ML)、药室内壁(EN)、表皮(EP) (图版Ⅱ:D-E)。腺质型绒毡层,细胞质浓厚,核仁显著。中层细胞往往由1~2层细胞组成,后期受另外两层壁细胞的挤压,严重变形,最后退化消失。次生造孢细胞时期的造孢细胞,细胞质浓厚,细胞间紧密排列。与初生造孢细胞时期相比,此时期药隔组织的薄壁细胞发育变大,不具有浓厚的细胞质,细胞核均靠近细胞壁的一边,可能是由于液泡形成与膨胀的结果;而维管束细胞仍然体积较小,且具备极高浓度的细胞质和明显的细胞核(图版Ⅱ:D-E)。通过对花药进行纵切观察可以发现,次生造孢细胞时期各个花药室的造孢细胞发育程度基本一致(图版Ⅱ:P)。次生造孢细胞发生游离,成为小孢子母细胞,外形变为圆形(图版Ⅱ:F)。
小孢子母细胞随后进行减数分裂(图版Ⅱ:G-H),绒毡层细胞质因消耗而减少,排列整齐紧密,绒毡层脱离花药壁,中层细胞被挤压吸收。第一次减数分裂,小孢子母细胞分裂形成二分体(图版Ⅱ:G),再由二分体同步分裂形成四分体,产生的四分体呈平面左右两两对称型(图版Ⅱ:H)。青秆竹每个花药中的不同药室之间小孢子母细胞发育时期不同步, 相同药室的小孢子发育时期也有差异,因此青秆竹花药进入减数分裂时期的发育程度并不一致。减数分裂完成后,四分体的四个子细胞彼此分开形成单核小孢子,单核小孢子个体较小,细胞质浓厚,无法观察到液泡等细胞器(图版Ⅱ:I)。同时,花药壁的绒毡层胞质逐渐减少,开始发生解体。单核靠边期绒毡层已经解体,单核小孢子的细胞质液化形成液泡,液泡将小孢子核挤压至边缘,小孢子壁处已经出现萌发孔(图版Ⅱ:J-K)。花粉粒成熟时期,液泡被挤压,细胞质重新填满花粉细胞,细胞核有丝分裂发育成为2核或3核花粉粒(图版Ⅱ:L-M)。
在花药成熟過程中,花药壁中层逐渐被挤压消失,花粉粒成熟以后,绒毡层完全退化为一层薄膜,与药室内壁紧密贴合(图版Ⅱ:J-O)。药室径
向壁显著增厚,纤维层逐渐生成(图版Ⅱ:N),此时成熟的花药壁仅余纤维层和花药壁表皮细胞共2层细胞。花药壁纵向开裂,释放花粉,开口处的表皮细胞体积较小,仍然具有明显的细胞质 (图版Ⅱ:O)。在雷竹雄配子体发育过程的研究中,表皮层在花药释放花粉时有帮助花药壁开裂的作用(初彩华等,2019)。
通过对青秆竹早期的花器官进行纵切可以发现,处于发育早期阶段的花药基部组织形态呈圆形, 并不尖锐, 与成熟的花药基部差别较大, 因此花药基部是在花药发育成熟的过程中逐渐突起变尖的(图版Ⅱ:Q)。通过纵切也可以发现,花药的维管束与小穗轴以及子房的维管束连接在一起(图版Ⅱ:Q)。
2.4 青秆竹花药发育异常
青秆竹开花后未能收获到种子,竹丛基部也未能发现实生苗的存在,因此青秆竹的结实率极低。通过对青秆竹成熟花药进行解剖观察发现,青秆竹花药的败育类型较多,这可能是导致青秆竹结实率低的主要原因。
关于青秆竹花药的败育类型主要总结为以下几种:(1)花药完全发生皱缩,药室几乎完全未形成,花药壁结构不可分辨(图版Ⅲ:A);(2)花药壁表皮细胞发育良好,但花药壁的其他几层细胞发育异常,药室完全皱缩,小孢子缺少发育空间(图版Ⅲ:B);(3)花药壁未开裂,未形成纤维层,花药室中空,无花粉粒形成(图版Ⅲ:C);(4)花药壁表皮细胞发育异常,细胞质过少,纤维层未形成,同时花粉粒发育不良,发生皱缩(图版Ⅲ:D);(5)花粉粒形成时绒毡层未溶解,导致空壳花粉粒遍布药室内部(图版Ⅲ:E);(6)小孢子母细胞无法发生游离及正常分化(图版Ⅲ:F)。
2.5 青秆竹花子房形态结构与发育
青秆竹子房1室,2层珠被,胚着生于远离子房基部的子房侧壁上,属于倒生胚珠,侧膜胎座(图版IV:A-B)。由于青秆竹开花数量有限,每朵小花仅有1枚子房,不同于花药的数量较多,受子房实验材料数量的限制,仅观察到受精后的受精胚的形态(图版IV:B),说明青秆竹部分花器官也能够正常完成授粉受精。经解剖不育小花发现,青秆竹的子房同样出现了发育异常的现象,可总结为3种类型:(1)子房的薄壁细胞发生严重解体(图版IV:C);(2)胚珠发育不完整,无法观察到珠被等结构(图版IV:D);(3)胚珠发生皱缩、退化,紧贴子房壁,这种现象可能是未正常授粉或受精引起的,需要进一步开展研究(图版IV:E)。
3 讨论与结论
Mc Clure在1934年提出关于竹类植物花序分为determinate inflorescence和indeterminate inflorescence两种理论(Mc Clure和洪效训,1982),20世纪80年代,耿伯介先生对其理论进行了补充说明,将其翻译成无限制花序和有限制花序(耿伯介,1986),无限制花序的小穗为假小穗,基部具有潜伏芽,而后林树燕等(2018)对3种竹子花的花序进行概括总结发现,竹类植物的花序应该属于混合花序,即花枝顶端小穗先发育,而小穗基部的花先开放,同时包含了有限制花序和无限制花序的特点。青秆竹的小穗和小穗上的花都是先从基部开始发育的,具有总状花序的特点。同时,青秆竹的小穗基部存在潜伏芽,成熟的小穗逐渐脱落,而基部不断地孕育新的花芽,并逐渐发育成新的小穗,这又符合有限花序的特征,属于传统意义上的“假小穗”。青秆竹的花序特征与王星等(2021)报道的黄条金刚竹(Pleioblastus kongosanensis)的小穗特征一致,均为小穗发育完成后,其基部又新分化出小穗。因此,青秆竹也应属于混合花序。
竹类植物在自然条件下开花少、授粉难、结实率低,一直都是竹类植物胚胎学研究的主要科学问题(林树燕等,2010),而且因为竹类植物生殖生长的周期很长,使研究工作更加困难。除了气候因素的影响(唐国建等,2016),青秆竹小穗基部不断产生新的小穗,可能也会引起养分的竞争、营养分配不均,进而可能会导致青秆竹花器官的结实率低。有关营养竞争引起败育的报道,如王曙光等(2006)认为巨龙竹的小穗中存在营养竞争,顶端的小花优先发育成熟,而巨龙竹本身此时已经处于濒死状态,营养不足以支撑其他小花的发育。
另外,雌雄配子体的发育状况对小花败育的影响也十分重要。在青秆竹花器官发育过程中的雄蕊败育类型很多,如花药整体发生皱缩;花药壁并未发生开裂,纤维层未形成;花药室中空,无花粉粒形成;花粉粒发育不良,发生皱缩等。绒毡层的作用主要是为小孢子提供营养和结构物质(胡适宜,2015),所以绒毡层发育不正常会导致药室内的小孢子发育成的花粉粒全部为空壳等现象,这类败育现象在其他竹种中也报道过,如在龙竹(Dendrocalamus giganteus)(陈舒怀和谭宏超,1997)、新小竹(Neomicrocalamus prainii)(初彩华等,2019)、绵竹(Bambusa intermedia)(王雨珺等,2017)、硬头黄竹(B. rigida)(李娟等,2020)、青丝黄竹(B. eutuldoides)(唐国建等,2016)、爬竹(Drepanostachyum scandeus)(庞延军等,1994)等竹种均出现诸如花粉粒空穴化、花药室中空、纤维层未形成、花药室皱缩等现象。此外,在青秆竹还发现了小孢子母细胞无法发生正常的游离及分化现象,该现象目前还未见其他竹种的相关报道。李娟等(2020)认为花粉败育可能是竹类植物败育过程中发生的共同现象,是竹类植物结实率低的共同原因。
雌蕊也是竹子繁衍下一代的重要一环,大多数竹种的雌蕊发育大致正常,如慈竹(Neosinocalamus affinis)(Wang et al., 2015)、月月竹(Menstruocalamus sichuanensis)(林树燕等,2009a)、巨龙竹(Dendrocalamus sinicus)(王曙光等,2006)等结实率低都是由其他原因导致的,但对孝顺竹(Bambusa multiplex)(林树燕等,2015)、鹅毛竹(Shibataea chinensis)(林树燕和丁雨龙,2012)、车筒竹(Bambusa sinospinosa)(Wang et al., 2015)觀察研究中,发现了异常发育的胚,如孝顺竹雌蕊中无胚珠发生,子房干瘪,胚珠败育以及鹅毛竹雌蕊中胚囊发育不正常等情况。本研究在青秆竹中发现了子房的薄壁细胞发生严重解体,胚珠发育不完整,胚珠发生皱缩、退化等现象,可见青秆竹雌蕊的发育同样也是影响其结实率的重要因素之一。因此,花粉发育状态异常、胚囊无法正常生长、雌蕊柱头无法正常受精等情况均会导致竹类植物的结实率具有显著的负面影响(林树燕,2009)。
综上所述,在自然生长的条件下,由于营养分配不均等条件导致的大多数败育花药在成熟期之前已经处于异常状态,有些败育类型的花药甚至无法正常释放花粉粒,这与de Souza等(2020)研究得出的花药成熟开裂释放花粉粒之后,花粉粒失去活性的结论不同。另外,雌蕊的败育也是导致结实率低的重要部分,雌蕊和雄蕊任何一方出现异常都会导致种子无法正常形成,de Souza等(2020)并未对雌蕊进行观察研究。
青秆竹小穗下部的小花先发育,又具有潜伏芽,同时具有有限花序和无限花序的特征,属于混合花序。小穗基部不断产生新的花芽,发育成新的小穗。小花包括内、外稃各1枚,浆片3枚,属于开放型花。浆片呈透明膜状,边缘具纤毛。雌蕊1枚,子房为三叉羽毛状柱头,子房上部被绒毛,属于长花柱型。子房单室,侧膜胎座,倒生胚珠。雄蕊6枚,花药4室,成熟花药的花药壁4层,由内向外分别为绒毡层、中层、药室内壁和表皮。花药与子房之间通过维管束与小穗轴进行连接。绒毡层为腺质型,随花药成熟发生退化,仅余一薄层紧贴花药壁,成熟花粉粒为3核花粉粒。青秆竹的花药和子房都出现了大量的败育情况,可能是导致青秆竹小花结实率低的重要原因。
参考文献:
AZZINI A, CIARAMELLO D, SALGADO ALB,et al., 1988. Densidade básica do colmo e fibras celulósicas em progênies de Bambusa tuldoides Munro [J]. Bragantia, 47(2): 239-246.
CHEN SH, TAN HC, 1997. Biological and ecological characteristics of Dendrocalamus giganteus [J]. Yunnan For Sci Technol, (4): 35-38. [陈舒怀, 谭宏超, 1997. 龙竹的生物学和生态学特性 [J]. 云南林业科技, (4): 36-39.]
CHU CH, HUANG L, WANG SG, 2019. Floral morphology and development of female and male gametophytes of Neomicrpcalamus prainii [J]. Acta Bot Boreal-Occident Sin, 39(5): 763-769. [初彩华, 黄玲, 王曙光, 2019. 新小竹花形态结构及雌、雄配子体的发育研究 [J]. 西北植物学报, 39(5): 763-769.]
DE SOUZA PF, DOS SANTOS CMR, REE J, et al., 2020. Male sterility in Bambusa tuldoides Munro [J]. Protoplasma, 257(4): 911-920.
DU F, XUE JR, YANG YM, et al., 2000. Study on flowering phenomenon and its type of bamboo in Yunnan in past fifteen years [J]. Sci Silv Sin, 36(10): 57-68. [杜凡, 薛嘉榕, 杨宇明,等, 2000. 15年来云南竹子的开花现象及其类型研究 [J]. 林业科学, 36(10): 57-68.]
GENG BJ, 1986. A preliminary study of the inflorescence type arising from bamboo and its variable [J]. J Wuhan Bot Res, 4(4): 323-336. [耿伯介, 1986.试论竹类的花序及其演变 [J]. 武汉植物学研究, 4(4): 323-336.]
GUO W, LEI G, QI LH, et al., 2019. Photosynthetic characteristics and leaf morphological characteristics of five bamboo species of Bambusa in Hainan Island during the rainy season [J]. Sci Silv Sin, 55(8): 63-72. [郭雯, 雷刚, 漆良華, 等, 2019. 海南岛簕竹属5个竹种雨季光合特性与叶片形态结构性状 [J]. 林业科学, 55(8): 63-72.]
HUANG JQ, HUANG HH, HE FJ, 1999. The formation of microspore and the development of male gametophyte of Phyllostachys praecox [J]. J Bamboo Res, 18(3): 55-58. [黄坚钦, 黄华宏, 何福基, 等, 1999. 雷竹的小孢子发生和雄配子体形成 [J]. 竹子研究汇刊, 18(3): 55-58.]
HU SY, 2015. Reproductive biology of angiosperms [M]. Beijing: Higher Education Press: 34-35. [胡适宜, 2015. 被子植物生殖生物学 [M]. 北京: 高等教育出版社: 34-35.]
LI J, WANG YF, CHU CH,et al., 2020. Studies on flower morphology and structure of Bambusa rigida [J]. For Res, 33(1): 28-34. [李娟, 王一方, 初彩华, 等, 2020. 硬头黄竹花的形态与结构研究 [J]. 林业科学研究,33(1): 28-34.]
LIN SY, 2009. Studies on the reproductive biology of Shibataea chinensis and Arundinaria simonii f. albostriatus [D]. Nanjing: Nanjing Forestry University. [林树燕, 2009. 鹅毛竹和异叶苦竹的生殖生物学研究 [D]. 南京: 南京林业大学.]
LIN SY, DING YL, 2012. Development of the male and female gametophytes in Shibataea chiensis (Bambusoideae) [J]. Acta Bot Boreal-Occident Sin, 32(5): 907-914. [林树燕, 丁雨龙, 2012. 鹅毛竹大小孢子及雌雄配子体发育 [J]. 西北植物学报, 32(5): 907-914.]
LIN SY, FU HJ, WAN YW, et al., 2019. Anther development and floral morphology characteristics of Bambusa oldhami ‘Xia Zao’ ZSX [J]. J Nanjing For Univ( Nat Sci Ed), 43(2): 7-13. [林树燕, 傅华君, 万雅雯, 等, 2019. ‘霞早’绿竹花形态特征及花药发育的组织学观察 [J]. 南京林业大学学报(自然科学版), 43(2): 7-13.]
LIN SY, HAO JJ, XIN H, et al., 2009. The megasporogenesis,microsporagenesis and the development of their female and male gametophyte in Menstruocalamus sichuanensis [J]. J Nanjing For Univ(Nat Sci Ed), 33(3): 9-12. [林树燕, 郝娟娟, 辛华, 等, 2009. 月月竹大、小孢子发生和雌、雄配子体发育研究 [J]. 南京林业大学学报(自然科学版), 33(3): 9-12.]
LIN SY, LI J, ZHAO R, et al., 2015. Studies on development of Megasporogenesis and female gametophyte of Bambusa multiplex [J]. J Nanjing For Univ(Nat Sci Ed), 39(4): 51-56. [林树燕, 李洁, 赵荣, 等, 2015. 孝顺竹花芽分化及小孢子发生与雄配子体发育 [J]. 南京林业大学学报 (自然科学版), 39(4): 51-56.]
LIN SY, SHI WW, MIU BB, et al., 2010. Research advance in reproduction biology of bamboos [J]. World Bamboo Ratt, 8(2): 1-6. [林树燕, 石文文, 缪彬彬, 等, 2010. 竹类植物生殖生物学研究进展 [J]. 世界竹藤通讯, 8(2): 1-6.]
LIN SY, WAN YW, FU HJ, et al., 2018. Research on inflorescence establishment and revision of inflorescence type in bamboo plants [J]. J Nanjing For Univ (Nat Sci Ed), 42(6): 1-6. [林树燕, 万雅雯, 傅华君, 等, 2018. 竹类植物花序建成及花序类型修正 [J]. 南京林业大学学报(自然科学版), 42(6): 1-6.]
LI ZL, 1996. Plant tissue production [M]. Beijing: Beijing University Press. [李正理, 1996. 植物组织制片 [M]. 北京: 北京大学出版社.]
MC CLURE FA, HONG XX, 1982. Reproductive phase of bamboos [J]. J Bamboo Res, 1(2): 104-116. [Mc Clure F A, 洪效训, 2012. 竹子的生殖相(上) [J]. 竹子研究汇刊, 1(2): 104-116.]
PANG YJ, YU FG, HU CH, 1994. Preliminary observation on abnormal development of the statements of Drepanostachyum microphyllum [J]. J Bamboo Res, 13(4): 42-46. [龐延军, 喻富根, 胡成华, 等, 1994. 爬竹雄蕊发育异常的初步观察 [J]. 竹子研究汇刊, 13(4): 42-46.]
SUN J, YU J, ZHANG PC,et al., 2015. Enantiomeric determination of four diastereoisomeric oxyneolignans from Bambusa tuldoides Munro [J]. Phytochem Analy, 26(1): 54-60.
SUN LF, GUO QR, WANG Q, et al., 2012. Flower organs morphology and structure of Phyllostachys edulis [J]. Sci Silv Sci, 48(11): 124-129. [孙立方, 郭启荣, 王青, 等, 2012. 毛竹花器官的形态与结构 [J]. 林业科学, 48(11): 124-129.]
TANG GJ, YANG JM, DING YL, et al., 2016. Studies on the flower morphology and structure in Bambusa eutuldoides [J]. J Nanjing For Univ (Nat Sci Ed), 40(2): 71-75. [唐国建, 杨金梅, 丁雨龙, 等, 2016. 青丝黄竹花形态与结构研究 [J]. 南京林业大学学报(自然科学版), 40(2): 71-75.]
WANG SG, PU XL, DING YL, et al., 2006. The structure of reproductive organs and development of the female and male gametophyte of Dendrocalamus sinicus [J]. Bull Bot Res, 26(3): 270-274. [王曙光, 普晓兰, 丁雨龙, 等, 2006. 巨龙竹生殖器官形态结构及雌、雄配子体的发育 [J]. 植物研究, 26(3): 270-274.]
WANG SG, PU XL, DING YL, et al., 2016. Reproductive characteristics of three bamboo species [J]. Pakistan J Bot, 47(6): 2301-2308.
WANG X, JIANG MY, LIN SY, et al., 2021. Inflorescence architecture of male and female gametophytes of Pleioblastus kongosanensis [J]. J Anhui Agric Univ, 48(1): 9-14. [王星, 姜明云, 林树燕, 等, 2021. 黄条金刚竹的花序建成及雌雄蕊发育 [J]. 安徽农业大学学报, 48(1): 9-14.]
WANG YJ, LUO J, CHEN NN, et al., 2017. Floral morphology and development of female and male gametophye of Bambusa intermedia Hsueh et Yi [J]. Bull Bot Res, 37(4): 492-498. [王雨珺, 罗剑, 陈楠楠, 等, 2017. 绵竹花形态结构及雌、雄配子体的发育研究 [J]. 植物研究, 37(4): 492-498.]
YI TP, SHI JY, MA LS, et al., 2008. Iconographia Bambusoidearum Sinicarum [M]. Beijing: Science Press: 146-147. [易同培, 石军义, 马丽莎, 等, 2008. 中国竹类图志 [M]. 北京: 科学出版社: 146-147.]
ZHOU Y, 2014.Process of extraction of Bambusa tuldoides Munro by orghogonal design [J]. J Pract Chin Med, 30(1): 67-68. [周滢, 2014. 竹茹水煎工艺正交实验研究 [J]. 实用中医药杂志, 30(1): 67-68.]
ZHANG WY, MA NX, 1990. Vitality of bamboo pollens and natural pollination in bamboo plants [J]. For Res, 3(3): 250-255. [张文燕, 马乃训, 1990. 竹类植物花粉的生活力和自然授粉 [J]. 林业科学研究, 3(3): 250-255.]
(责任编辑 周翠鸣)
2624501186219
