不同生境三种杜鹃属植物花特征比较
2022-03-17胡德美董溪吴江明胡莲宇唐明王晓月
胡德美 董溪 吴江明 胡莲宇 唐明 王晓月






摘 要: 生境影響植物花部特征的分化。为探究不同人为干扰强度生境是否影响杜鹃花特征分化和资源分配,以及不同颜色杜鹃花分布情况,该研究以百里杜鹃保护区高、中、低人为干扰强度生境的马缨杜鹃、迷人杜鹃和露珠杜鹃为实验材料,采用五点取样法统计不同生境杜鹃花的植株数量,游标卡尺测量花部特征,光学显微镜统计花粉数,海洋光谱仪测定花色的反射光谱,Gephi软件绘制植株数量的网络图。结果表明:(1)迷人杜鹃植株数量显著多于马缨杜鹃和露珠杜鹃。(2)低强度干扰生境中,马缨杜鹃的营养器官和繁殖器官特征指数大于高、中强度干扰生境;不同干扰强度生境对迷人杜鹃花部特征的影响不明显;中、低干扰强度生境露珠杜鹃的营养器官和繁殖器官特征指数显著大于高强度干扰生境。(3)马缨杜鹃和迷人杜鹃的花色在400~500 nm波长有较高的反射,露珠杜鹃的花色在400~700 nm波长有较高的反射。(4)在高、中强度干扰生境中,迷人杜鹃与其他两种杜鹃花分布紧密,在低强度干扰生境中马缨杜鹃和迷人杜鹃分布更紧密。该研究对杜鹃花的种群管理和生物多样性保护提供重要的理论基础。
关键词: 杜鹃花属, 生境, 花部特征, 花色, 网络结构图
中图分类号: Q948.11
文献标识码: A
文章编号: 1000-3142(2022)01-0161-13
收稿日期: 2020-12-20
基金项目: 国家自然科学基金(31901208);国家自然科学基金委员会贵州省人民政府喀斯特科学研究中心项目(U1812401);贵州省科学技术基金资助项目(2019/1237);贵州师范大学博士科研启动项目(2017/22) [Supported by the National Natural Science Foundation of China(31901208); the Joint Fund of the National Natural Science Foundation of China and the Karst Science Research Center of Guizhou Province(U1812401); Funded by the Science and Technology Foundation of Guizhou Province(2019/1237); Doctoral Research Project of Guizhou Normal University(2017/22)]。
第一作者: 胡德美(1995-),硕士研究生,主要研究方向为进化与传粉生态学,(E-mail)1838319797@qq.com。
*通信作者: 王晓月,博士,副教授,主要研究方向为进化与传粉生态学,(E-mail)wang.xiaoyue1989@163.com。
Comparison of floral traits of three Rhododendron
species in different habitats
HU Demei1,3, DONG Xi1,3, WU Jiangming3, HU Lianyu3, TANG Ming1,2, WANG Xiaoyue1,2*
( 1. State Key Laboratory of Southwest Karst Mountain Biodiversity Conservation of National Forestry and Grassland Administration, Guizhou Normal
University,
Guiyang 550025, China; 2. Key Laboratory of Plant Physiology and Development in Guizhou Province, Guizhou Normal
University, Guiyang 550025, China; 3. College of Life Science, Guizhou Normal University, Guiyang 550025, China )
Abstract: Habitats influent the divergence of floral traits. To test whether habitats change could affect the floral traits and resources allocation of Rhododendron plants and investigate the distribution of Rhododendron plants in different colors. R. delavayi, R. agastum and R. irroratum in high, medium and low intensity human interference habitats were taken as materials. Five-point sampling method was used to count the number of Rhododendron plants in different habitats, floral traits were measured by a vernier caliper, the number of pollen grains were counted by a light microscope, the spectral reflectance of colors was measured by an ocean spectrometer, and the Gephi software was used to draw network diagrams of the number of plants. The results were as follows: (1) The plant number of R. agastum was significantly higher than that of R. delavayi and R. irroratum. (2) The trait indexes of vegetative and reproductive organs of R. delavayi in low intensity interference habitat were higher than those in high and medium intensity interference habitats; Different intensity interference habitats had no significant influence on floral traits of R. agastum; The trait indexes of vegetative and reproductive organs of R. irroratum in medium and low intensity interference habitats were significantly higher than those in high intensity interference habitats. (3) The colors of R. delavayi and R. agastum showed higher reflection values at 400-500 nm wavelengths, while colors of R. irroratum showed higher reflection values at 400-700 nm wavelengths. (4) R. agastum was distributed closely with the other two species of Rhododendron in high and medium intensity interference habitats, while R. delavayi and R. agastum were more closely distributed in low intensity interference habitat. This study provides a crucial theoretical basis for the population management and biodiversity conservation of Rhododendron species.
Key words: Rhododendron, habitats, floral traits, flower colors, diagram of community network structure
环境是指生境中的生物和非生物因素(气候、资源等),同时也包括与其他物种的相互作用(竞争、捕食、互利共生等)(Schluter, 2000, 2001)。全球环境变化的不确定性和人为干扰因素的加剧,给植物的进化发育带来很大考验,植物是否能够并采取何种方式应对环境变化成为植物进化生态学研究的焦点问题(代文魁,2017)。
植物花部特征不仅包括花的结构、颜色、气味、分泌物质类型及其产量,还包括某一时间花的大小、开放数量和花在花序上的空间排列模式等(黄双全和郭友好,2000)。不同生境影响植物花部特征的分化(Juan et al., 1995; Warren et al., 2001; Thomas et al., 2004; 李鹂等,2007),从而影响繁殖成功。例如,相较于林缘旷地生境,分布在竹林生境的蝴蝶花(Iris japonica),其单花花冠的长与宽更大,其子房、花部(除花柄)以及单花总生物量更小(王永健等,2010)。降雨多的地区,植物花开口倾向于向下,从而保护植物的雌雄蕊结构(陈颖卓,2012)。花部特征不仅决定有性繁殖的成功率、繁殖后代的遗传组成和适合度(赖家业等, 2007;方海涛等,2007;王伟等,2008),同时也是其对外部环境的响应。按照资源分配理论,植物将有限资源投资到有性繁殖和生长,不同生境中蝴蝶花花部各组分差异性分配显示出不同的有性繁殖策略(王永健等,2010)。
杜鹃花是世界著名的观赏花卉,是世界十大名花之一,在国内外被广泛作为观赏花卉进行培育。杜鹃花属植物的形态、大小、颜色等特征有很大的分化,花形态根据花瓣融合情况的不同可以分为开放的漏斗状、钟状或者管状花(Stevens, 1976),花直径也在20~50 cm之间变异,颜色有白色、黄色、粉色、深红色甚至是紫红色(Ma et al., 2016; Huang et al., 2017)。我国的西南地区是杜鹃花属植物分布和分化中心(庄平等,2012;王禹等,2020),其中贵州百里杜鹃的杜鹃种质资源丰富而独特。近年来对百里杜鹃自然保护区杜鹃花属种植资源的调查发现保护区共有6个亚属、4个组的33种杜鹃花(包括亚种和变种)(张长芹等,2015)。吴倩楠等(2017)应用生态位宽度、生态位相似度以及Pianka 重叠度计算方法对百里杜鹃优势种群生态位的特征进行研究发现马缨杜鹃、迷人杜鹃、露珠杜鹃、映山红是该群落的优势种,马缨杜鹃和露珠杜鹃生态位重叠度降低,是种群对资源利用的长期适应性导致的。目前很少有人研究百里杜鹃保护区及其周围群落中的不同生境是否对杜鹃花的花部特征以及雌雄资源的分配产生影响,且不同颜色的杜鹃花在群落中的分布情况也鲜见研究。
我们实地考察发现百里杜鹃自然保护区对杜鹃林干扰强度有差异,主要可分为三种:一是景区核心地带受到人为干扰程度最大(景区保护局重点管理区域和游客观光旅游区),我们把这种生境称为高强度干扰生境(high intensity interference habitat,简称HIIH);二是位于核心景区的外围,受到保护局人为管理和游客干扰的程度较低,称为中强度干扰生境(medium intensity interference habitat,简称MIIH);三是远离核心景区地带的生境,该生境完全没有受到保护局的人为管理和游客干扰,称为低强度干扰生境(low intensity interference habitat, 簡称LIIH)。为比较同种杜鹃花在以上三种生境中的花部特征是否不同,以及探究不同颜色杜鹃花在群落中的分布规律,本文以这三种生境中的优势种马缨杜鹃、迷人杜鹃和露珠杜鹃为实验材料,研究以下问题:(1)不同干扰强度生境中,三种杜鹃花的植株数量是否存在差异?(2)同一种杜鹃花在不同干扰强度生境中,花部特征是否存在变异?(3)三种杜鹃花的花色反射光谱是否存在差异?(4)定量分析不同干扰强度生境中三种杜鹃花植株数量的分布关系。
1 材料与方法
1.1 实验地点与材料
百里杜鹃自然保护区位于贵州省西北部、毕节市中部 (105°45′30″—106°04′45″ E、27°08′30″—27°20′00″ N),属于高原山地过渡地带中的山地地貌类型,海拔在1 400~1 900 m之间(陶云等, 2013;张长芹等, 2015),地处低纬度高海拔地区,属亚热带湿润季风气候,年平均温度12.0 ℃,年降雨量1 120.1 mm(罗贤值等,2012)。主要分布有马缨杜鹃(Rhododendron delavayi)、露珠杜鹃(R. irroratum)、迷人杜鹃(R. agastum)(图1)等杜鹃属植物35个种(含亚种、变种),分为6个亚属、 4个组及9个亚组,花期3—4月(40~50 d)( 黄红霞,2006;陈翔等,2010)。
1.2 实验方法
1.2.1 统计不同生境中三种杜鹃花植株数量
为探究高、中、低强度干扰生境中三种杜鹃花分布情况和数量关系,根据五点取样法(方精云等,2009)在每一种生境中设置5个50 m × 50 m(样方的长×宽)的样方,详细统计每个样方中马缨杜鹃、迷人杜鹃、露珠杜鹃的植株数量,并计算每种杜鹃花占三种杜鹃花总数的百分比。
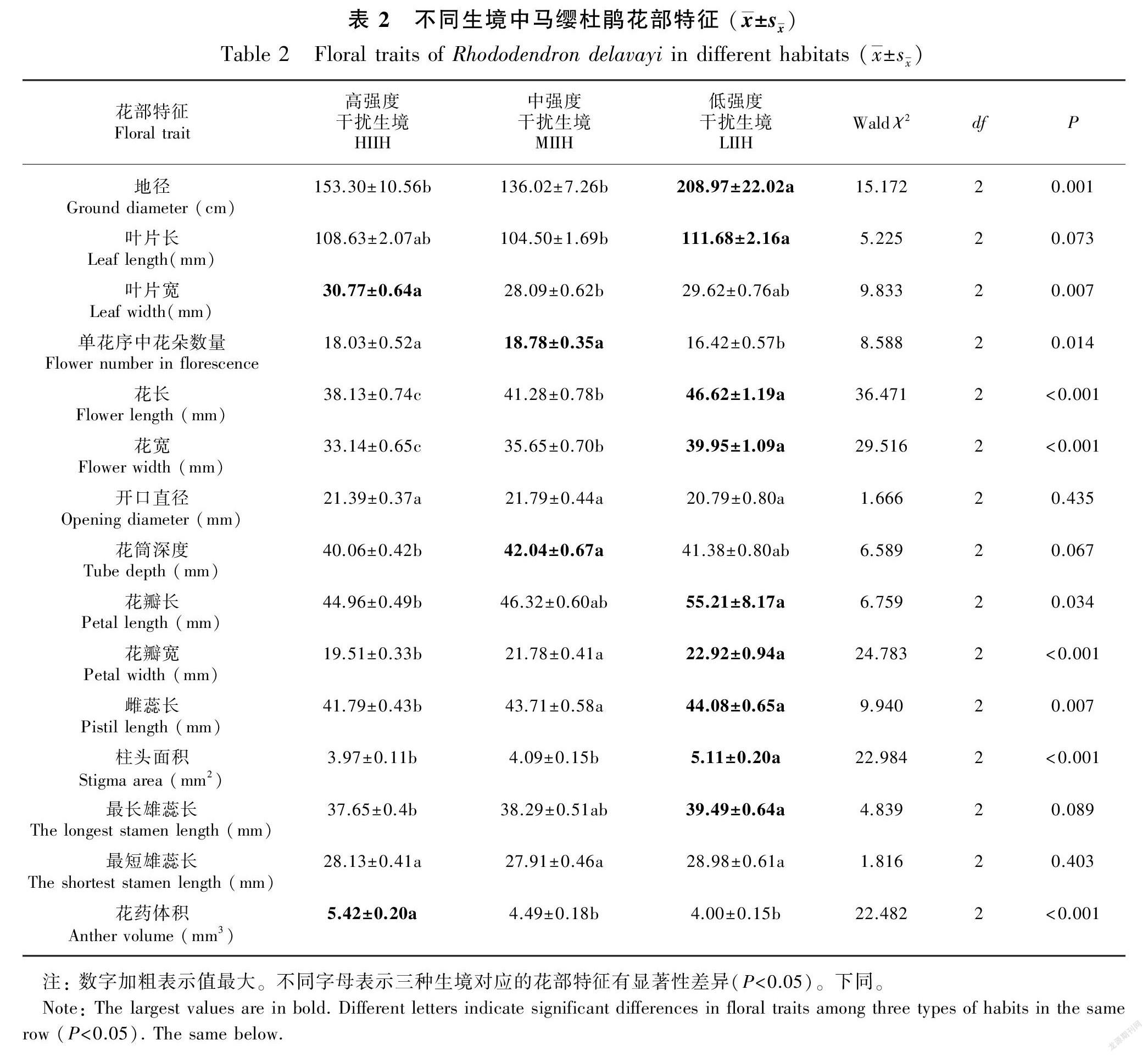
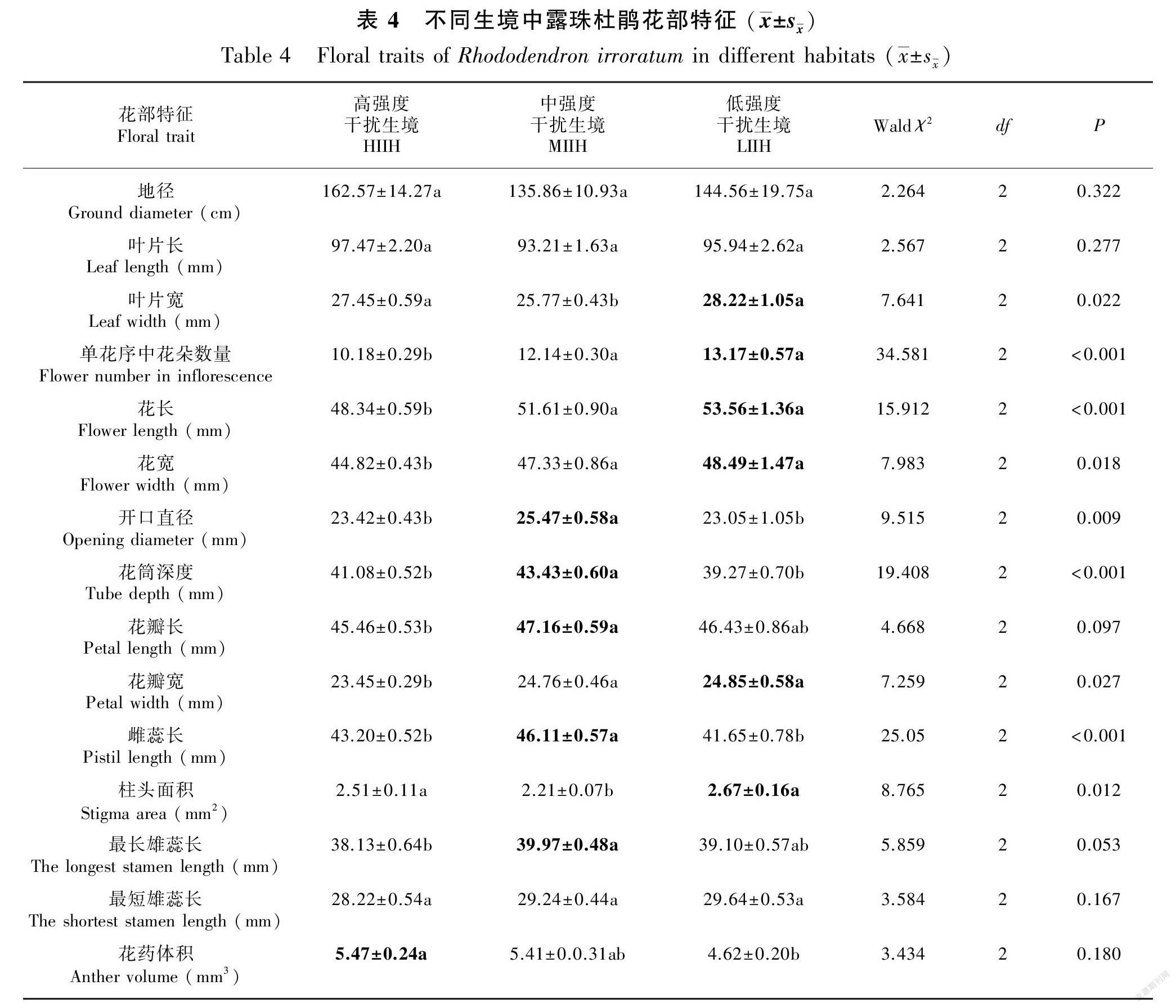
1.2.2 测量不同生境中三种杜鹃花的花部特征 为比较同一种杜鹃花在高、中、低强度干扰生境中的花部特征是否存在差异,根据1.2.1部分设定的样方,每个样方中每种杜鹃花随机选择6株,用卷尺测量每株杜鹃花的地径(精确到0.1 cm)。每一株杜鹃花上随机选择1个单花序,统计该花序上杜鹃花的朵数。随机选择1朵处于盛花期的杜鹃花,用游标卡尺(精确到0.01 mm)测量其叶片长、叶片宽、花长、花宽、开口直径、筒深、花瓣长、花瓣宽、雌蕊长、柱头长、柱头宽、柱头深度、最长雄蕊长度、最短雄蕊长度、花药长、花药宽、花药厚等花部特征指标(图2);为了更直观地表示三种杜鹃花的柱头大小和花药大小,将其柱头近似看成椭圆,用椭圆面积计算公式(S=πab,式中:S表示柱头面积;a表示1/2柱头长;b表示1/2柱头宽,计算它们的柱头面积;将其花药近似看成长方体,用长方体的体积计算公式(V=a×b×c,式中:V表示花药体积;a表示花药长;b表示花药宽;c表示花药厚),计算它们的花药体积。
为比较同一种杜鹃花在高、中、低强度干扰生境中的花粉数量是否存在差异,根据1.2.1部分设定的样方,每个样方中每种杜鹃花随机选择6株,每一株上随机采集一个花苞,将其保存在装有75%酒精的2 mL离心管中。在实验室,先将花苞转移至4 mL离心管中用8 mol·L-1的NaOH溶液软化4 h,然后用镊子充分研磨花药制成花粉悬浊液,并定容至4 mL,充分摇匀后用移液枪取20 μL花粉悬浊液置于载玻片上, 每个样品取3滴, 在光学显微镜下统计其花粉数量 (分别记作a1, a2,
a3),计算花苞中花粉的总数量(a1+a2+a3)/3×200(王晓月等,2019)。
1.2.3 测定三种杜鹃花的花色反射光谱 为比较百里杜鹃自然保护区中三种杜鹃花花瓣颜色反射光谱的差异,在设定的样方中,随机选择处于盛花期的马缨杜鹃、迷人杜鹃、露珠杜鹃各10朵花,用海洋光谱仪(Ocean Optics RPH-1)配备的光纤反射探针(QR400-7-UV-BX)在固定器的作用下呈45°角照射在花瓣上(Johnson & Andersson, 2002),获得三种杜鹃花花瓣颜色的反射值。每隔5 nm将波长在300~700 nm之间的反射值取平均值(Liu et al., 2018),然后在Excel 2010软件中绘制三种杜鹃花花瓣的反射光谱散点图。
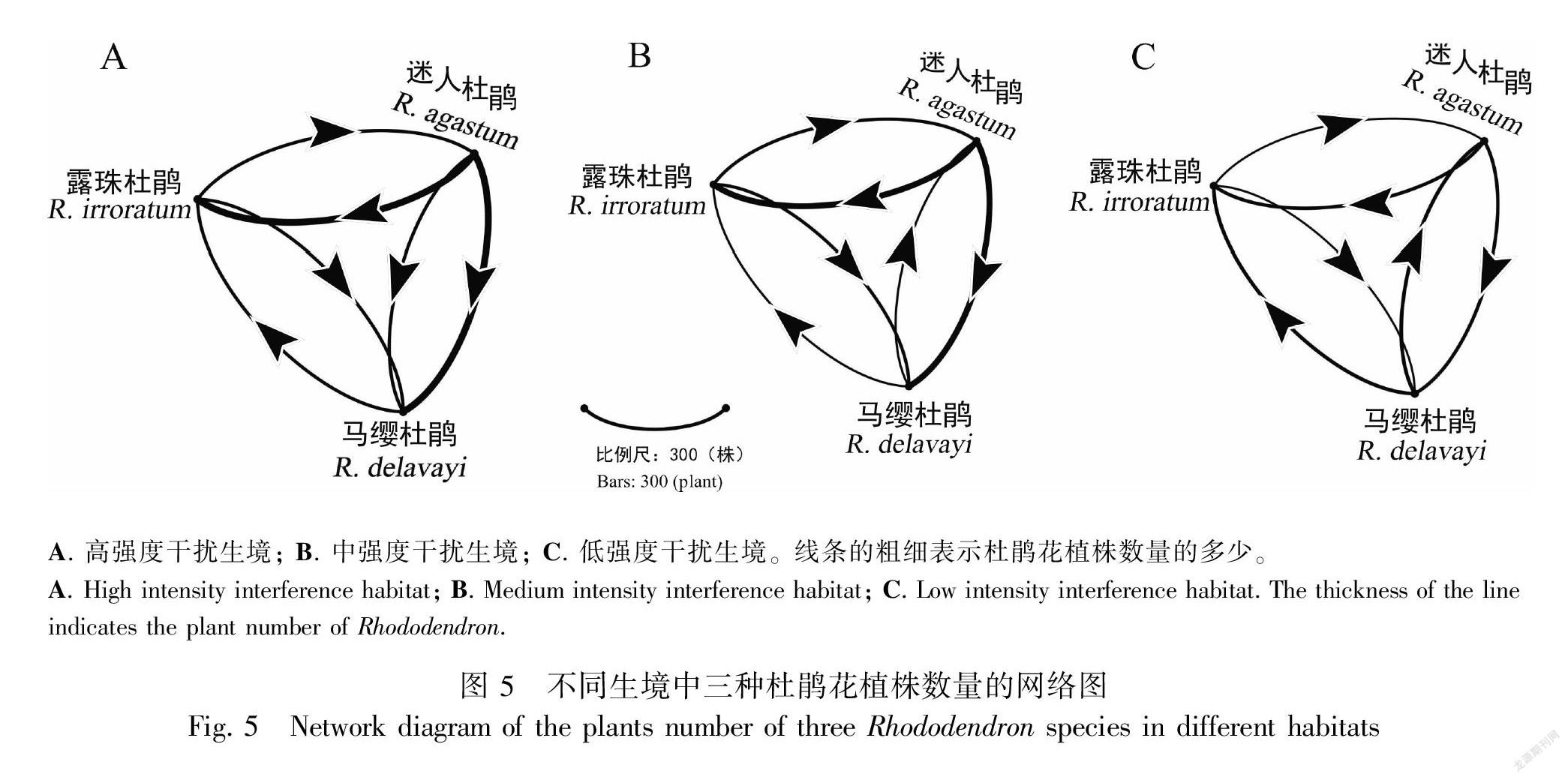
1.2.4 构建三种杜鹃花植株数量的网络图 为定量分析百里杜鹃自然保护区中三种杜鹃花植株数量分布情况,根据1.2.1部分统计的不同生境种三种杜鹃花的植株数量,使用群落分析软件Gephi绘制三种杜鹃花植株数量关系的定量网络图,其中每一种杜鹃花作为定量网络图中的一个节点,三种杜鹃花之间的联系构成网络图连接的线条,杜鹃花植株数量的多少表示三种杜鹃花在群落中连接的强度(方强和黄双全,2012)。
1.2.5 数据分析 为比较同一种杜鹃花在高、中、低强度干扰生境中的花部特征是否存在差异,用广义线性模型(generalized linear model,GLM)中的正态分布模型-恒定函数比较分析,其中花部特征指标作为因变量,不同生境作为自变量;采用泊松分布模型-对数线性函数分析三种杜鹃花在不同生境的植株数量并比较花苞中的花粉数量,其中植株数量、花粉数量作为因变量,不同生境作为自变量。所有的数据分析在SPSS 20.0软件(IBM Inc., New York, NY)中进行。
2 结果与分析
2.1 不同生境中三种杜鹃花植株数量
对高、中、低强度干扰生境中三种杜鹃花植株数量的分析结果表明,迷人杜鹃的植株数量(91.00±10.22,x±sx)显著多于(Waldχ2=12.435, P=0.002, df=2)马缨杜鹃(50.00±9.66)和露珠杜鹃(46.87±9.86)的植株数量(将同一种杜鹃花在三种生境中的植株数量合并分析)。具体每种生境中三种杜鹃花植株数量的分析比较结果见表1。
2.2 不同生境中三种杜鹃花的花部特征
广义线性模型分析结果表明,马缨杜鹃、迷人杜鹃和露珠杜鹃的花部特征在高、中、低强度干扰生境表现的差异不同。对不同生境中马缨杜鹃的营养器官研究表明,低强度干扰生境下的地径显著大于高、中强度干扰生境(Waldχ2=15.172, P=0.001, df=2);低强度干扰生境中的叶片长显著大于中强度干扰生境(P<0.05),但与高强度干扰生境无显著性差异(P>0.05);高强度干扰生境中的叶片宽显著大于中强度干扰生境(P<0.05),但与低强度干扰生境无显著性差异(P>0.05)。对不同生境中马缨杜鹃的繁殖器官研究表明,低强度干扰生境中的花宽、柱头面积显著大于高、中强度干扰生境(所有的P<0.01);中、低强度干扰生境中的花瓣宽和雌蕊长度显著大于高强度干扰生境(所有的P<0.05);低强度干扰生境中的花瓣长、最长雄蕊长度显著大于高强度干扰生境(所有的P<0.05),但与中等强度干扰生境中的这两项花部特征指标无显著性差异(所有的P>0.05);高、中强度干扰生境中单个花序的花朵数量显著高于低强度干扰生境(Waldχ2=8.588, P=0.014, df=2);高强度干扰生境中的花药体积显著大于中、低强度干扰生境(Waldχ2=22.482, P<0.001, df=2);高、中、低强度干扰生境中马缨杜鵑的花开口直径、最短雄蕊长度没有显著性差异(所有的P>0.05)(表2)。
对不同生境中迷人杜鹃的营养器官研究表明,高、中、低强度干扰生境中的地径、叶片长、叶片宽都没有显著性差异(所有的P>0.05)。对不同生境中迷人杜鹃的繁殖器官研究表明,高、中、低强度干扰生境中的花宽、花瓣长、花瓣宽、最长雄蕊、最短雄蕊没有显著性差异(所有的P>0.05);高强度干扰生境中的开口直径、花冠筒深度显著大于中、低强度干扰生境(所有的P<0.05);中强度干扰生境中单个花序的花朵数量显著大于高、低强度干扰生境(Waldχ2=19.811, P<0.001, df=2);中强度干扰生境中的花药体积显著大于高强度干扰生境,但与低强度干扰生境没有显著性差异(Waldχ2=11.48, P=0.003, df=2);低强度干扰生境中的柱头面积显著大于中强度干扰生境(Waldχ2=4.499, P=0.034, df=1),但与高强度干扰生境无显著性差异(Waldχ2=3.054, P=0.081, df=1),同时高、中强度干扰生境中的该项花部特征指标也无显著性差异(Waldχ2=0.607, P=0.436, df=1);低强度干扰生境中的雌蕊长度与高、中强度干扰生境无显著性差异(Waldχ2=3.977, P=0.137,df=2)(表3)。综上所述,不同干扰强度生境对迷人杜鹃的营养器官和繁殖器官特征影响不大。
对不同生境中露珠杜鹃的营养器官研究表明,高、中、低强度干扰生境中的地径和叶片长无显著性差异(所有的P>0.05);高、低强度干扰生境中的叶片宽显著大于中强度干扰生境(Waldχ2=7.641, P=0.022, df=2)。对不同生境中露珠杜鹃的繁殖器官研究表明,高、中、低强度干扰生境中的最短雄蕊长度没有显著性差异(Waldχ2=3.584, P=0.167, df=2);中、低强度干扰生境中的花展示(单个花序中的花朵数量、花长、花宽、花瓣宽)显著大于高强度干扰生境(所有的P<0.05);中强度干扰生境中的花开口直径、花筒深度、雌蕊长显著大于高、低强度干扰生境(所有的P<0.05);中強度干扰生境中的花瓣长、最长雄蕊长显著大于高强度干扰生境(所有的P<0.05),但与低强度干扰生境无显著性差异(所有的P>0.05);低强度干扰生境中的柱头面积显著大于中强度干扰生境 (Waldχ2=8.933, P=0.003, df=1), 但与高强度干扰生境无显著性差异(Waldχ2=0.623, P=0.430, df=1);高强度干扰生境中的花药体积显著大于低强度干扰生境(Waldχ2=4.658, P=0.031, df=1)(表4)。
对不同生境中三种杜鹃花的花粉数量分析发现,马缨杜鹃的花粉数量在高、低强度干扰生境中无显著性差异(Waldχ2=0.086, P=0.769, df=1),但都显著高于中强度干扰生境(Waldχ2=14.274, P=0.001, df=2)(图3:A);迷人杜鹃的花粉数量在高、中强度干扰生境中无显著性差异(Waldχ2=0.217, P=0.642, df=1),但都显著高于低强度干扰生境(Waldχ2=9.853, P=0.007, df=2)(图3:B);露珠杜鹃的花粉数量在高、中强度干扰生境中无显著性差异(Waldχ2=1.220, P=0.269, df=1),但都显著高于低强度干扰生境(Waldχ2=14.037, P=0.001, df=2)(图3:C)。
2.3 三种杜鹃花的花色反射光谱
马缨杜鹃和迷人杜鹃花色的反射光谱变化规律比较相近,露珠杜鹃花色的反射光谱与上述二者不一致。具体表现为马缨杜鹃和迷人杜鹃的花色在400~500 nm(红色区域)有较高的反射值,且迷人杜鹃(粉红色)的反射值高于马缨杜鹃(深红色),而露珠杜鹃(黄白色)在400~700 nm之间的反射值较高(图4)。
2.4 不同生境中三种杜鹃花植株数量的网络图
构建三种杜鹃花植株数量的定量网络图结果表明,在高、中强度干扰生境中,迷人杜鹃的植株数量显著高于露珠杜鹃和马缨杜鹃(所有的P<0.05),因此,在高、中强度干扰生境中,迷人杜鹃与其他两种杜鹃花都分布紧密,同时马缨杜鹃与露珠杜鹃在这两种干扰强度生境中也分布紧密;在低强度干扰生境中,露珠杜鹃的植株数量显著低于马缨杜鹃和迷人杜鹃(Waldχ2=51.562, P<0.001, df=2),因此在该生境中,马缨杜鹃和迷人杜鹃分布更紧密(图5)。
3 讨论与结论
本研究主要比较不同干扰强度生境马缨杜
鹃、迷人杜鹃和露珠杜鹃花部特征的分化和资源分配,并探究不同颜色杜鹃花在群落中的分布情况。实验结果表明,百里杜鹃自然保护区中迷人杜鹃植株数量显著多于马缨杜鹃和露珠杜鹃;马缨杜鹃和露珠杜鹃的花特征对人为干扰因素更敏感,在低干扰强度的生境中长势最好,而迷人杜鹃的花部特征受不同干扰强度生境的影响不大;在高、中强度干扰生境中迷人杜鹃(粉红色)与马缨杜鹃(深红色)和露珠杜鹃(黄白色)都分布紧密,在低强度干扰生境中马缨杜鹃和迷人杜鹃分布更紧密。
生境差异会导致物种内花部特征分化。不同生境对景宁玉兰的萌枝形态和生殖特征(果实大小、单果种子数以及每从结实率等)影响极大(杜有新等,2018)。报春花科珍珠菜属植物Lysimachia arvensis 不同的居群花瓣颜色有分化,大部分居群是红色花,蓝色花的分布频率随着纬度的升高而下降,随着温度和光照时间的增长而增加(Arista et al, 2013)。花部特征反映了植物对某一类群传粉者的适应(Fgri et al., 1966; Charles et al., 2004; 黄双全,2007)。杜鹃花属植物在不同生境中部分花部特征表现出的差异可能与不同生境中传粉者的转变和访问频率差异有关。在高干扰强度生境中,当杜鹃花处于盛花期时,既是传粉者访问频率最高的时期,也是游客参观的旺季。游客的高频干扰会降低杜鹃花属植物传粉者(鸟类或者蜂类)的访问频率,植物为了保障一定的繁殖率则倾向于自交,这会影响植物花部特征的分化。有研究表明,更长的单花期、单花序展示的增大、花粉数量的增加等花部性状的变化,是与其传粉环境相互适应的,并且与交配系统转变也一致(代文魁,2017)。较多的花朵数量可以增大花展示,更能吸引传粉者访花,马缨杜鹃在高干扰强度生境中单花序的花朵数量最多,迷人杜鹃在中干扰强度生境最多,露珠杜鹃在中、低干扰强度生境中最多。开口直径和花筒深度作为传粉者进入花内部的两个重要花部特征参数,能影响传粉者的种类,较小的开口只能接受体积较小的访问者访问,反之亦然,而花筒的深浅则可以说明传粉者吻长的长度。马缨杜鹃在中强度干扰生境中的花筒深度显著大于高强度干扰生境,迷人杜鹃高强度干扰生境下的开口直径和花筒深度均显著大于中、低强度干扰生境,露珠杜鹃在中强度干扰生境中开口直径和花筒深度显著大于高、低强度干扰生境。植物个体内雌蕊长度的变异被认为是与繁殖成功率相关的参数,前人做了大量关于植物个体内花特征变异和性功能特化的研究(Lloyd, 1980; Diggle, 1995, 1997; Granado-Yela et al., 2017),且有研究表明长花柱可能通过促进花粉的落置或者促进花柱中花粉管的生长而提高植物的繁殖成功(Campbell, 1989; Bernasconi et al., 2007),王晓月等(2019)对梅花个体内花柱长度的变异及其对繁殖成功的影响研究表明,长柱型为母本的梅花有利于花粉在柱头上的萌发以及花粉管的伸长, 并能显著提高坐果率。我们的研究结果显示,中、低强度干扰生境中马缨杜鹃的雌蕊长度显著长于高强度干扰生境下的雌蕊,高强度干扰生境下迷人杜鹃的雌蕊长度显著长于中强度干扰生境的雌蕊,中强度干扰生境中露珠杜鹃雌蕊长度显著高于高、低强度干扰生境。因此,在今后继续开展不同生境三种杜鹃花传粉者种类和访问频率研究非常必要。
不同生境物种多样性、根系微生物种类和植物自身的生活习性会导致杜鹃花属植物在不同生境中花部特征的差异。比如露珠杜鹃有耐干旱和瘠薄的特性(王加国等,2020),高强度干扰生境下,人工管理干预大,反而不是露珠杜鹃最适宜的生境;景区为了营造景观效果,人为砍伐了群落内的其他物种,营造杜鹃纯林,使群落受到极大破坏,人为改变了群落结构及物种组成,降低了物种多样性, 凋落物覆盖度降低, 加速土壤蒸发, 不利于水土保持,同时导致群落稳定性降低,土壤真菌微生物的种类增多,抵御病虫害的能力下降(陈明梅等,2019;陈胜群等,2019;任纬恒等,2020)。
杜鹃花属植物在不同生境中花部特征表现出的差异可能是基因与群落水平共同作用的结果。花色是受基因调控的表型,有研究表明分布于百里杜鹃保护区的马缨杜鹃和露珠杜鹃种内个体在花色上的变异尤为明显,马缨杜鹃花色由血红色至粉红变异,露珠杜鹃花色由乳黄至淡粉红色变异(黄承玲等,2016),植物花色的变异会影响传粉者的访花偏好(Gigord et al., 2001; Takahashi et al., 2015;唐汉青等,2019)。对群落中马缨杜鹃、迷人杜鹃和露珠杜鹃的物种数量统计发现,迷人杜鹃的物种数量显著高于马缨杜鹃和露珠杜鹃。迷人杜鹃是Balfour于1917年发表的新种,1996年David F. Chamberlain在杜鹃花属修订时提出其可能是杂交形成的,并推测其父本和母本是马缨杜鹃和大白杜鹃(黄承玲等,2016),基于其花色等形态特征的假设,前人对其做了分子生物学方面的证实(Zhang et al., 2007; Zha et al., 2008)。直到2010年,Zha等(2010)才首次证实迷人杜鹃是马缨杜鹃和露珠杜鹃杂交的F1代,迷人杜鹃成为将杂交后代误定為种类的典型例子,这说明迷人杜鹃可能遗传了双亲的稳定遗传性状,具有较强的适应能力和抗干扰能力。自然群落中普遍存在异种花粉转移现象(Fang & Huang, 2013, 2016),在大多数植物中异种花粉转移会造成种间花粉干扰,对植物的生殖成功不利,但是对于马缨杜鹃和露珠杜鹃来说,两者之间的异种花粉转移无疑是有利的,产生了稳定遗传的杂交后代迷人杜鹃。
植物花部特征主要受到环境的选择压力(王慧,2014;Babu et al., 2016)。人类对自然界的频繁干扰会对传粉者与植物之间关系产生严重影响,如生境片段化产生的小居群导致传粉者资源匮乏或丧失,外来种引入导致传粉者竞争,气候变暖会导致植物与传粉者之间的错配等,使开花植物正常的传粉进程受到限制,从而直接影响植物雌性和雄性的繁殖适合度(Aguilar et al., 2006; Memmott et al., 2007; Hegland et al., 2009)。黄承玲等(2016)对百里杜鹃的杜鹃属资源调查时发现杜鹃原生境面临着严峻考验,杜鹃原始次生林带自然更新能力弱; 旅游基础设施建设导致局部环境发生改变,旅游活动对杜鹃群落的影响明显; 人为干扰使杜鹃原生境明显破碎。对百里杜鹃自然保护区杜鹃花属资源的保护与管理,需要科研工作者和管理部门不断创新管理方式和坚持资源可持续发展道路。
致谢 感谢中国百里杜鹃自然保护区管理委员会王灵军和其他工作人员,以及贵州师范大学硕士研究生唐汉青对野外工作的支持和帮助,河南科技大学方强副教授,张涛硕士研究生对数据分析的协助。
参考文献:
AGUILAR R, ASHWORTH L, GALETTO L, et al., 2006. Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis [J]. Ecol Lett, 9(8): 968-980.
ARISTA M, TALAVERA M, BERJANO R, et al., 2013. Abiotic factors may explain the geographical distribution of flower colour morphs and the maintenance of colour polymorphism in the scarlet pimpernel [J]. J Ecol, 101(6): 1613-1622.
BABU R, PAUDEL M, SHRESTHA M, et al., 2016. Coevolutionary elaboration of pollination-related traits in an alpine ginger (Roscoea purpurea) and a tabanid fly in the Nepalese Himalayas [J]. New Phytol, 211(4): 1402-1411.
BERNASCONI G, LANG DJ, SCHMID B, 2007.Microgametophyte population sizes and plant reproductive output in the insect-pollinated Prunella grandiflora (Lamiaceae) [J]. New Phytol, 173(2): 393-400.
CAMPBELL DR, 1989. Measurements of selection in a hermaphroditic plant: variation male in male and female pollination success [J]. Evolution, 43(2): 318-334.
CARLOS GY, LUIS B, LUIS C, et al., 2017.Unusual positional effects on flower sex in an andromonoecious tree: Resource competition, architectural constraints, or inhibition by the apical flower? [J]. Am J Bot, 104(4): 608-615.
CHEN MM, LI WJ, YANG R, et al., 2019. Effects of Rhododendron delavayi litters on soil moisture evaporation in Baili Azalea Scenic Area [J]. Water Soil Conserv Bull, 39(6): 60-65. [陈明梅, 李苇洁, 杨瑞, 等, 2019. 百里杜鹃景区马缨杜鹃林凋落物对土壤蒸发的影响 [J]. 水土保持通报, 39(6): 60-65.]
CHEN SQ, WEI XL, AN MT, et al., 2019. Species diversity of plant communities at different succession stages in Baili Rhododendron Nature Reserve of Guizhou Province [J]. Acta Bot Boreal-Occident Sin, 39(7): 1298-1306. [陈胜群, 韦小丽, 安明态, 等, 2019. 贵州百里杜鹃自然保护区不同演替阶段植物群落物种多样性 [J]. 西北植物学报, 39(7): 1298-1306.]
CHEN X, HUANG JY, XIE H, et al., 2010. A study on plant resource of Rhododendron (Ericaceae) in Nature Reserve of Azalea Forest [J]. Sci Guizhou, 28(4): 26-34. [陳翔, 黄家勇, 谢华, 等, 2010. 百里杜鹃自然保护区杜鹃属植物资源研究 [J]. 贵州科学, 28(4): 26-34.]
CHEN YZ, 2012.The role of rain and pollinators in the evolution of floral traits [D]. Wuhan: Wuhan University: 6-13. [陈颖卓, 2012. 雨水和传粉者在花部特征演化中的作用 [D]. 武汉: 武汉大学: 6-13.]
DAI WK, 2017. Variations in floral traits among populations and the contributions to pollination adaption in two Pedicularis species [D]. Wuhan:Wuhan Botanical Garden, Chinese Academy of Sciences: 1-15. [代文魁, 2017. 两种马先蒿属植物花部性状居群间的分异与传粉适应 [D]. 武汉: 中国科学院武汉植物园: 1-15.]
DAVID GL, 1980. Sexual strategies in plants. i. an hypothesis of serial adjustment of maternal investment during one reproductive session [J]. New Phytol, 86(1): 69-79.
DU YX, WU WJ, LIU YJ, et al., 2018. Morphological and reproductive characteristics of the sprouts of Magnolia sinostellata thickets in different habitats [J]. Acta Ecol Sin, 38(23): 8417-8424. [杜有新, 吴伟建, 刘跃钧, 等, 2018. 不同生境下景宁玉兰灌丛萌株形态及其生殖特征 [J]. 生态学报, 38(23): 8417-8424.]
FANG HT, SIQIN BT, 2007. Blossom character and insect pollination of Prunus mongolica Maxim. [J]. Chin J Ecol, 26(2): 177-181. [方海涛, 斯琴巴特, 2007. 蒙古扁桃的花部综合特征与虫媒传粉 [J]. 生态学杂志, 26(2): 177-181.]
FANG JY, WANG XP, SHEN ZH, et al., 2009. Methods and protocols for plant community inventory [J]. Biodivers Sci, 17(6): 533-548. [方精云, 王襄平, 沈泽昊, 等, 2009. [植物群落清查的主要内容、方法和技术规范 [J]. 生物多样性, 17(6): 533-548.]
FANG Q, HUANG SQ, 2012. Progress in pollination networks: network structure and dynamics [J]. Biodivers Sci, 20(3): 300-307. [方强, 黄双全, 2012. 传粉网络的研究进展:网络的结构和动态 [J]. 生物多样性, 20(3): 300-307.]
FANG Q, HUANG SQ, 2013. A directed network analysis of heterospecic pollen transfer in a biodiverse community [J]. Ecology, 94(5): 1176-1185.
FANG Q, HUANG SQ, 2016. A paradoxical mismatch between interspecific pollinator moves and heterospecific pollen receipt in a natural community [J]. Ecology, 97(8): 1970-1978.
FENSTER CB, ARMBRUSTER WS, WILSON P, et al., 2004. Pollination syndromes and floral specialization [J]. Ann Rev Ecol Evol S, 35(1): 375-403.
FGRI K, VAN DER, PIJL L, 1966. The principles of pollination ecology [M]. Oxford: Pergamon Press: 184-197.
GIGORD LD, MACNAIR MR, SMITHSON A, 2001.Negative frequency dependent selection maintains a dramatic flower color polymorphism in the rewardless orchid Dactylorhiza sambucina (L.) Soo. [J]. Proc Natl Acad Sci USA, 98(7): 6253-6255.
HEGLAND S, JOAR N, ANDERS L, et al., 2009. How does climate warming affect plant-pollinator interactions? [J]. Ecol Lett, 12(2): 184-195.
HUANG CL, HUANG JY, MA YP, 2016. Baili Rhododendron of Guizhou Province—a map of Rhododendron resources [M]. Beijing: China Forestry Press: 29-43. [黃承玲, 黄家湧, 马永鹏, 2016. 贵州百里杜鹃——杜鹃属资源图志 [M]. 北京: 中国林业出版社: 29-43.]
HUANG HX, 2006. Studies on tourism application and Rhododendron resource investigation in Baili Rhododendron National Forest Park [D]. Beijing: Beijing Forestry University: 5-7. [黄红霞, 2006. 百里杜鹃国家森林公园杜鹃花属植物资源调查与旅游应用研究 [D]. 北京: 北京林业大学: 5-7.]
HUANG SQ, 2007. Study on plant-pollinator interaction and its significances [J]. Biodivers Sci, 15(6): 569-575. [黄双全, 2007. 植物与传粉者相互作用的研究及其意义 [J]. 生物多样性, 15(6): 569-575.]
HUANG SQ, 2014. Most effective pollinator principle of floral evolution: evidence and query [J]. Life Sci, 26(2): 118-124. [黄双全, 2014. 花部特征演化的最有效传粉者原则:证据与疑问 [J]. 生命科学, 26(2): 118-124.]
HUANG SQ, GUO YH, 2000. Advances in the study of pollination biology [J]. Chin Sci Bull, 45(3):225-237. [黄双全, 郭友好, 2000. 传粉生物学的研究进展 [J]. 科学通报, 45(3): 225-237.]
HUANG ZH, SONG YP, HUANG SQ, 2017.Evidence for passerine bird pollination in Rhododendron species [J]. Aob Plant, 9(6): 62-65.
JOHNSON SD, ANDERSSON S, 2002. A simple field method for manipulating ultraviolet reflectance of flowers [J]. Can J Bot, 80(12): 1325-1328.
JUAN A, AMOTS D, 1995. Variations in habitat, season, flower traits and pollinators in Dimorphic narcissus tazetta L. (Amaryllidaceae) in Israel [J]. New Phytol, 129(1): 135-145.
LAI JY, PAN CL, QIN WG, et al., 2007. The floral syndrome and its pollination adaptability in Kmeria septentrionalis (Magnoliaceae) [J]. Acta Bot Yunnan, 29(3): 303-308. [赖家业, 潘春柳, 覃文更, 等, 2007. 单性木兰花部综合特征及其传粉适应性 [J]. 云南植物研究, 29(3): 303-308.]
LI L, DANG CL, 2007. Floral syndrome and breeding system of Erigeron breviscapus [J]. Acta Ecol Sin, 27(2): 571-578. [李鹂, 党承林, 2007. 短葶飞蓬(Erigeron breviscapus)的花部综合特征与繁育系统 [J]. 生态学报, 27(2): 571-578.]
LIU CQ, NIU Y, PENG DL, et al., 2018. Are superior ovaries damaged by the bills of flower-visiting birds and does this preclude adaptation to bird pollinators? [J]. Bot J Linn Soc, 187(3): 499-511.
LUO ZX, LUO YY, ZHANG CG, 2012. Analysis on the characteristics of precipitation change in Dafang County in recent 50a [J]. Weather Guizhou, 36(2): 28-29. [罗值贤, 罗玉友, 张成刚, 2012. 大方县近50a来降水变化特征分析 [J]. 贵州气象, 36(2): 28-29.]
MA YP, XIE WJ, SUN WB, et al., 2016. Strong reproductive isolation despite occasional hybridization between a widely distributed and a narrow endemic Rhododendron species [J]. Sci Res, 6(1): 46-91.
MEMMOTT J, CRAZE P, WASER N, et al., 2007.Global warming and the disruption of plant-pollinator interactions [J]. Ecol Lett, 10(8): 710-717.
PAMELA KD, 1995. Architectural effects and the interpretation of patterns of fruit and seed development [J]. Ann Rev Ecol System, 26(1): 531-552.
PAMELA KD, 1997.Ontogenetic contingency and floral morphology: The effects of architecture and resource limitation [J]. Int J Plant Sci, 158(6): 99-107.
PAUDEL BR, SHRESTHA M, BURD M, et al., 2016. Coevolutionary elaboration of pollination-related traits in an alpine ginger (Roscoea purpurea) and a tabanid fly in the Nepalese Himalayas [J]. New Phytol, 211(4): 1402-1411.
REN WH, TANG M, WANG LJ, et al., 2020. The characteristics of the soil fungus community of Rhododendron delavayi Franch in Baili Rhododendron Nature Reserve [J]. Genom Appl Biol, 39(3):1172-1177. [任緯恒,唐明,王灵军,等, 2020. 百里杜鹃保护区马缨杜鹃土壤真菌群落特征 [J]. 基因组学与应用生物学, 39(3): 1172-1177.]
SCHEMSKE DW, BIERZYCHUDEK P, 2001.Evolution of flower color in the desert annual Linanthus parryae: Wright revisited [J]. Evolution, 55(7): 1269-1282.
SCHEMSKE DW, BIERZYCHUDEK P, 2007. Spatial differentiation for flower color in the desert annual Linanthus parryae: was wright right? [J]. Evolution, 61(11): 2528-2543.
SCHLUTER D, 2000.The ecology of adaptive radiation [M]. Oxford: Oxford University Press: 172-178.
SCHLUTER D, 2001. Ecology and the origin of species [J]. Trend Ecol Evol, 16(3): 372-380.
STEVENS PF, 1976. The altitudinal and geographical distributions of flower types in Rhododendron section Vireya, especially in the Papuasian species, and their significance [J]. Bot J Linn Soc, 72(5): 1-33.
TAKAHASHI T, TAKAKURA K, KAWATA M, 2015.Flower color polymorphism maintained by overdominant selection in Sisyrinchium sp. [J]. J Plant Res, 128(6): 933-939.
TAO Y, HUANG CL, HUANG JY, et al., 2013. Study on community of Rhododendron irroratum Baili Rhododendron Scenic Spot in Guizhou [J]. Agric Sci Anhui, 41(15): 6791-6793. [陶云, 黄承玲, 黄家湧, 等, 2013. [贵州百里杜鹃风景名胜区露珠杜鹃群落研究 [J]. 安徽农业科学, 41(15): 6791-6793.]
THOMAS F, CHRISTIAN K, 2004.Altitudinal differences in flower traits and reproductive allocation [J]. Flora, 199(1): 70-81.
WANG H, 2014. Divergence in floral traits under the selection of pollinators in sympatric Corydalis species [D]. Wuhan: Wuhan University: 7-12. [王慧, 2014. 同域分布紫堇属植物传粉选择压力下的花部特征分化 [D]. 武漢: 武汉大学: 7-12.]
WANG JG, LI WJ, ZHANG HJ, et al., 2020. Study the diversity and stability of plant communities in Baili Rhododendron Scenic Area in Guizhou Province [J]. For Resouc Manag, (2): 120-125. [王加国, 李苇洁, 张红军, 等, 2020. 贵州百里杜鹃景区植物群落多样性与稳定性研究 [J]. 林业资源管理, (2): 120-125.]
WARREN J, MACKENZIE S, 2001.Why are all colour combinations not equally representedas flower-color polymorphisms? [J]. New Phytol, 151(1): 237-41.
WANG W, DAI HG, CHEN FD, et al., 2008. Correlation between floral traits of chrysanthemum (Dendranthema morifolium) and insect visitors [J]. J Plant Ecol, 32(4): 776-785. [王伟, 戴华国, 陈发棣, 等, 2008. 菊花花部特征及花冠精油组分与访花昆虫的相关性 [J]. 植物生态学报, 32(4): 776-785.]
WANG XY, ZHU XX, YANG J, et al., 2019. Variation in style length and the effect on reproductive success in Chinese plums (Armeniaca mune) [J]. Biodivers Sci, 27(2):159-167. [王晓月, 朱鑫鑫, 杨娟, 等, 2019. 梅花个体内花柱长度的变异及其对繁殖成功的影响 [J]. 生物多样性, 27(2): 159-167.]
WANG YJ, FANG X, ZHONG ZC, 2010. Effect of different habitats on floral, fruit and seed traits of Iris japonica Thunb. [J]. Acta Ecol Sin, 30(17): 4628-4635. [王永健, 方兴, 钟章成, 2010. 不同生境对蝴蝶花花部与果实特征的影响 [J]. 生态学报, 30(17): 4628-4635.]
WANG Y, ZHANG GH, HE JS, et al., 2020. Research progress of Rhododendron flower color [J]. World For Res, 33(5): 19-24. [王禹, 张广辉, 赫京生, 等, 2020. 杜鹃花色研究进展 [J]. 世界林业研究, 33(5): 19-24.]
WU QN, DONG JW, ZHENG Y, et al., 2017. Niches of main plant species in Baili Rhododendron National Forest Park [J]. J Nanjing For Univ(Nat Sci Ed), 41(2): 175-180. [吴倩楠, 董建文, 郑宇, 等, 2017. 百里杜鹃国家森林公园优势种生态位研究 [J]. 南京林业大学学报(自然科学版), 41(2): 175-180.]
ZHANG CQ, HUANG CL, HUANG JY, et al., 2015. Investigation of germplasm resources of genus Rhododendron in Baili Nature Reserve in Guizhou [J]. Plant Diver Resour, 37(3): 357-364. [张长芹, 黄承玲, 黄家勇, 等, 2015. 贵州百里杜鹃自然保护区杜鹃花属种质资源的调查 [J]. 植物分类与资源学报, 37(3): 357-364.]
ZHANG JL, ZHANG CQ, GAO LM, et al., 2007. Natural hybridization origin of Rhododendron agastum (Ericaceae) in Yunnan, China: inferred from morphological and molecular evidence [J]. J Plant Res, 120(3): 457-463.
ZHA HG, RICHARD IM, SUN H, 2010.Asymmetric hybridization in Rhododendron agastum: a hybrid taxon comprising mainly F1s in Yunnan, China [J]. Ann Bot, 105(1): 89-100.
ZHA HG, RICHARD IM, HANG S, 2008. Morphological and molecular evidence of natural hybridization between two distantly related Rhododendron species from the Sino-Himalaya [J]. Bot J Linn Soc, 156(1): 119-129.
ZHUANG P, ZHENG YR, SHAO HM, et al., 2012. An assessment on the adaptability of Rhododentron plants under ex situ conservation [J]. Biodivers Sci, 20(6): 665-675. [莊平, 郑元润, 邵慧敏, 等, 2012. 杜鹃属植物迁地保育适应性评价 [J]. 生物多样性, 20(6): 665-675.]
(责任编辑 周翠鸣)
1272501186234
