浙江省水源地水库浮游动物食性功能群特征及水质响应
2022-03-11徐杭英于海燕晁爱敏周胜利李共国
徐杭英,于海燕,晁爱敏,周胜利,李 平,李共国
1.浙江省生态环境监测中心,浙江 杭州 310007
2.浙江万里学院生物与环境学院,浙江 宁波 315100
生物监测可以系统反映污染物对生物生长的影响及其在生物体内的转化和迁移,在水环境监测与生态健康管理中的重要性日益突出[1]。 生物监测系统因具有快速性、宽谱性、直观性、连续性、经济性等特点,在水质预警的应用中具有明显优势。 目前,生物监测系统在降低突发性污染事件影响、保障城市供水水质安全方面,已成为国际公认和通用的有效方法[2]。 浮游动物是水域生态系统中重要的生物组成部分,容易受到环境因素的影响,在食物链、物质转化、能量流动、信息传递等水域生态过程中起着至关重要的作用。 浮游动物群落变化特征可作为水质监测的指示器[3-4],但是传统分类方法难以体现水生生物的生态功能,为此,生态学家提出了“功能群”的概念。 功能群是从个体的环境适应性、生态位竞争机制角度出发,将具有相同环境适应策略的种类划分为同一个类群,其物种特征同环境的联系更为紧密[5]。 例如:在高有机物负荷的溪流,嗜菌纤毛虫的占比会增加;而在有机物浓度低、冠层开阔的溪流,则会有更多的嗜藻纤毛虫[6]。 功能群研究有助于直观反映生态环境影响生物群落的生态过程,便于提升对不同地域水生生物资源的科学管理水平,对于发展和检验群落生态学理论、更好地理解水生态系统及其生物多样性状况也十分重要,可为运用模型进行生态系统功能定量化研究奠定基础。 功能群的划分需要建立在了解每一种浮游动物的食性、测量其个体大小等前期准备工作的基础上[7],目前国内对浮游植物、底栖动物和鱼类功能群的划分及分析较为深入,但对浮游动物食性功能群的研究较少,关注点主要集中在浮游动物食性功能群对浮游植物的控制作用[8]、与蓝藻暴发的关联[9]以及功能群间相互作用的生态效应上[10]。 本文以地处亚热带的浙江水源地水库为例,通过调查研究浮游动物食性功能群的变化特征,并将其与水库水体综合营养状态指数(Comprehensive of Trophic Level Index,TLIc)进行多元逐步回归与通径分析,找出水体TLIc 的决策因子和限制因子,以期为采用浮游动物食性功能群参数监测水源地水库水质提供思路。
1 材料与方法
1.1 站位布设与采样方法
在浙江省内选取8 座饮用水水源地水库,每座水库取水口处各设置1 个浮游动物采样点(K1~K8)。 8 个浮游动物采样点分布见图1。

图1 水源地水库浮游动物采样点分布示意图Fig.1 Distribution of zooplankton sampling stations in water source reservoirs
于2014—2018 年的1 月、4 月、7 月和10 月,由全省各地方监测站定点、定时(上午09:00—上午11:00)分别采样与送样。 采样和计数过程按《淡水浮游生物研究方法》[11]进行。 定量样品用2.5 L 采水器采集,分别取上、下层水样各10 L,用25 号筛绢制成的浮游生物网(网孔孔径64 μm)过滤浓缩,当即用5%的甲醛溶液固定,带回实验室镜检。 同时,测定水体水温(温度计法)、透明度(透明度盘法)、pH(玻璃电极法,GB/T 6920—1986)、溶解氧(电化学探头法,HJ 506—2009)、总氮(碱性过硫酸钾消解紫外分光光度法,HJ 636—2012)、总磷(钼酸铵分光光度法,GB/T 11893—1989)、氨氮(纳氏试剂分光光度法,HJ 535—2009)和叶绿素a(分光光度法,HJ 897—2017)等水质参数。
1.2 标本鉴定与计算方法
轮虫、枝角类、桡足类分别按《中国淡水轮虫志》[12]、《中国动物志 节肢动物门 甲壳纲 淡水枝角类》[13]、《中国动物志 节肢动物门 甲壳纲 淡水桡足类》[14]鉴定。 轮虫生物量按体积法计算[10],枝角类和桡足类生物量分别按各自的体长-体重回归方程计算[15-16]。 桡足类幼体和成体数量按该种桡足类幼体或成体出现的频率乘以样本总数计算。 多样性指数按Shannon-Wiener[17]方法统计。 水体TLIc 参考《地表水环境质量评价办法(试行)》[18]计算。
1.3 浮游动物食性功能群划分
参考相关研究文献[7-9],根据浮游动物的大小、摄食习性以及浮游动物之间的相互作用,结合实际监测情况,将浙江省水源地水库浮游动物划分为滤食轮虫、捕食轮虫、小型滤食动物、中型滤食动物、中型捕食动物和大型捕食动物6 个浮游动物食性功能群。 大型捕食动物中,仅发现透明薄皮溞(Leptodorakindti)1 种,且出现频率极低,仅见于夏季的K1 站(2018 年,5.0 ind. /L)、秋季的K7 站(2017 年,1.5 ind. /L)和K8 站(2016年,2.8 ind. /L)。 由于数据太少,该类群不宜作为水质预测模型中的变量因子,因此,只划分了5个浮游动物食性功能群。
1.4 统计分析
以水库水体TLIc 为因变量,滤食轮虫丰度(x1)、捕食轮虫丰度(x2)、小型滤食动物丰度(x3)、中型滤食动物丰度(x4)、中型捕食动物丰度(x5)、总浮游动物丰度(x6)、滤食轮虫生物量(x7)、捕食轮虫生物量(x8)、小型滤食动物生物量(x9)、中型滤食动物生物量(x10)、中型捕食动物生物量(x11)、总浮游动物生物量(x12)、物种多样性指数(x13)为自变量,进行水库水体TLIc 与浮游动物食性功能群参数的逐步回归和通径分析[19]。 自变量与因变量之间的相关系数可分解为直接作用系数和间接作用系数。 通径系数能表征自变量对因变量的影响的相对重要性,从而通过比较通径系数绝对值的大小,确定对因变量有显著影响的自变量的主次顺序[20]。 通过计算决策系数,确定主要决策因子(决策系数为最大正值)和限制因子(决策系数为最小负值)[21]。 以上计算分析均应用DPS 数据处理系统[19]完成。
2 结果与讨论
2.1 水库水质理化参数
8 座水源地水库的水体营养状态存在很大差异,K1 站水体营养水平显著高于K3~K6 站(P<0.05),K4、K5 站又显著高于K3 站(表1)。
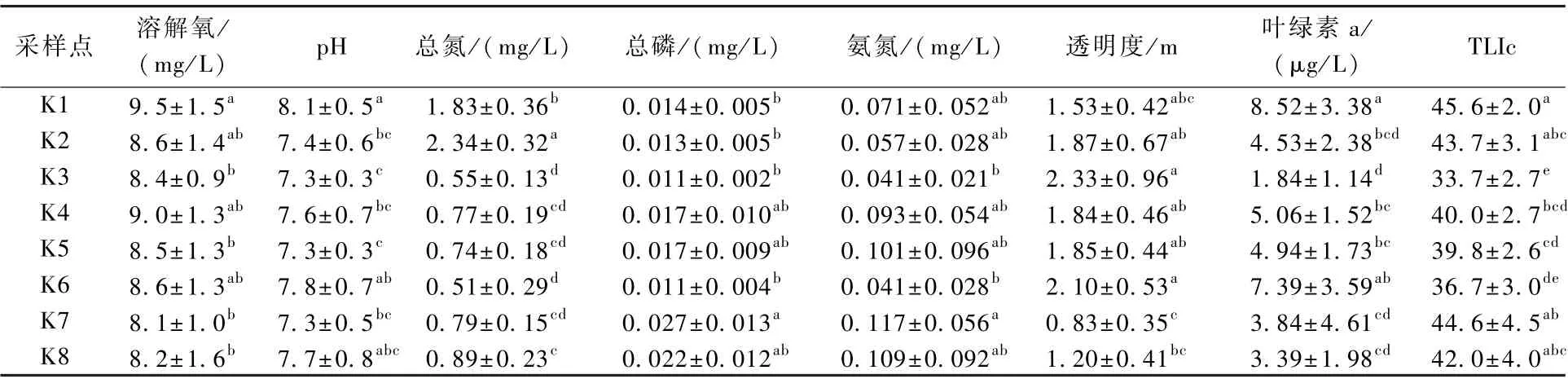
表1 2014—2018 年浙江省水源地水库水质参数(平均值±标准差)Table 1 Water quality parameters of water source reservoirs in Zhejiang Province from 2014 to 2018 (mean ± SD)
此外,K1 站溶解氧含量显著高于K3、K5、K7、K8 站(P<0.05);K1 站pH 显著高于K2~K5、K7 站(P<0.05);K2 站总氮含量显著高于其他各站,K1 站总氮含量显著高于K3~K8 站,K8 站又显著低于K3、K6 站(P<0.05);K7 站总磷含量显著高于K1~K3、K6 站(P<0.05);K7 站氨氮含量显著高于K3、K6 站(P<0.05);K3、K6 站水体透明度均显著高于K7、K8 站,K2 站又显著高于K7站(P<0.05);K1 站叶绿素a 含量显著高于K2~K5、K7、K8 站,K4、K5 站又显著高于K3 站(P<0.05)。
2.2 功能群参数的水平分布和季节变化
调查期间,浙江省8 座饮用水水源地水库浮游动物食性功能群的分类情况及主要代表性种类见表2,浮游动物食性功能群参数和物种多样性指数的水平分布见表3。 由表2、表3 可知,总丰度x6的 变 幅 为 11.4 ind. /L ( K2 站) ~271.3 ind. /L(K1 站),平均值为110.7 ind. /L,以小型滤食动物(71.8 ind. /L,占64.5%)和滤食轮虫(35.0 ind. /L,占31.4%)为主;总生物量x12的变幅 为44.0 μg/L(K2 站) ~ 755.0 μg/L(K1站),平均值为382.6 μg/L,以小型滤食动物(291.2 μg/L,占76.1%)和中型滤食动物(35.0 μg/L,占9.1%)为主;物种多样性指数x13介于2.0~2.4,平均值为2.2。

表2 2014—2018 年浙江省水源地水库浮游动物食性功能群划分Table 2 Classification of zooplankton functional feeding groups in water source reservoirs of Zhejiang Province from 2014 to 2018
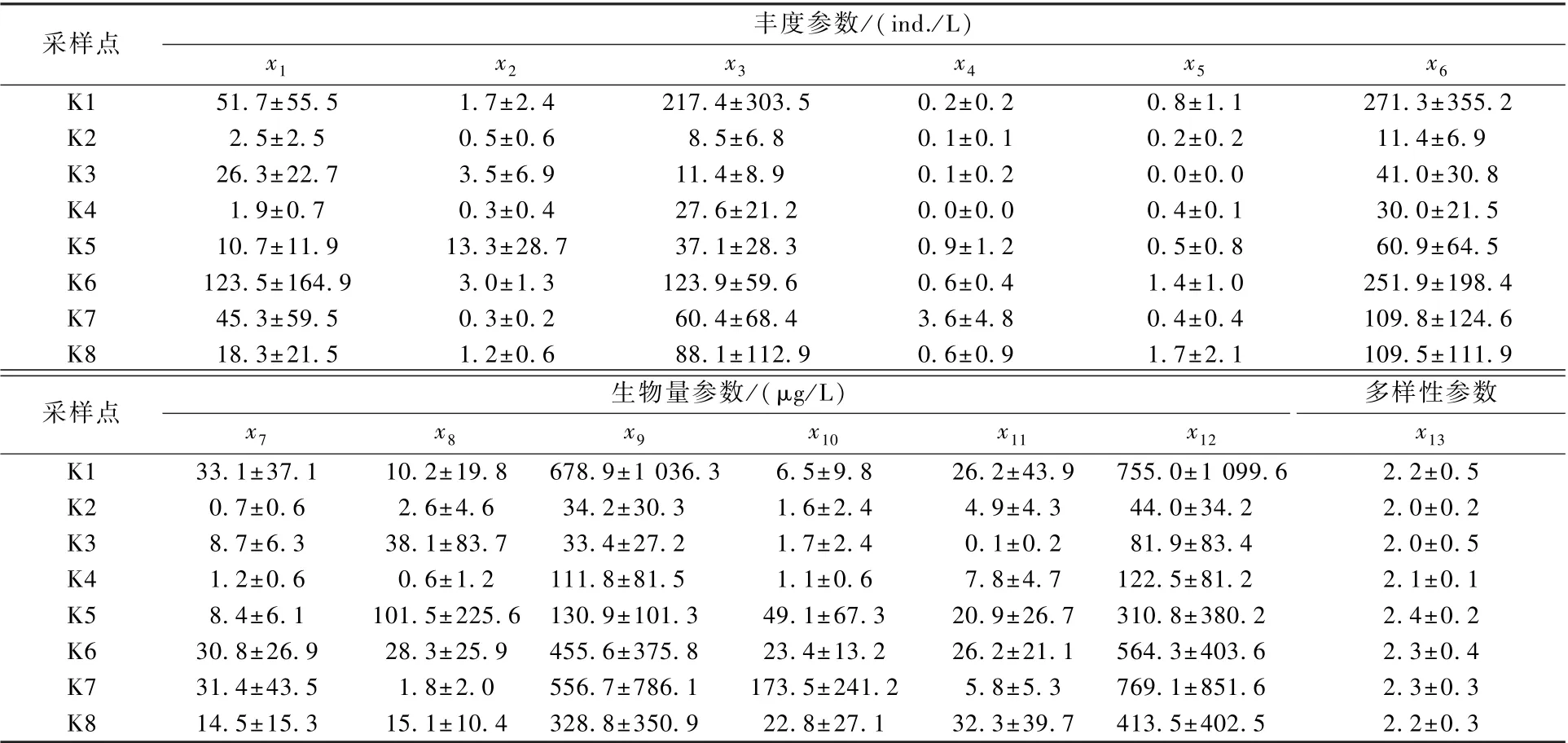
表3 2014—2018 年浙江省水源地水库浮游动物食性功能群参数(平均值±标准差)Table 3 Horizontal variation of zooplankton feeding functional group parameters in water source reservoirs of Zhejiang Province from 2014 to 2018 (mean ± SD)
水库浮游动物食性功能群x1~x13参数在各采样点间无显著差异(P>0.05)。 所有功能群参数中,变异幅度最大的是x8,K3~K5 站x8的变异幅度均在200%以上。
2014—2018 年水库浮游动物食性功能群参数和物种多样性指数的季节变化见表4。 表4 显示,总丰度x6和总生物量x12的季节变化均表现为夏季>秋季>春季>冬季。 除中型滤食动物丰度x4和生物量x10以春季为最高外,其余主要浮游动物食性功能群参数(如x1、x3、x9等)的季节变化均表现为夏季>秋季>春季>冬季。
2.3 功能群参数与水体营养水平的关系
浮游动物食性功能群参数与水体营养水平之间的逐步回归分析结果见表5。 表5 中,4 个季节的多元回归方程均有效(复相关系数R≥0.463,P≤0.040)。 浮游动物食性功能群参数x3有3 次入选回归方程,x1、x6、x7、x11和x13均有2 次入选回归方程,x10有1 次入选回归方程。 在通径分析过程中发现:x6有2 次成为影响水体TLIc 的重要因子,分别是春季和夏季水体TLIc 波动的限制因子;x10为冬季水体TLIc 波动的限制因子;x7、x13分别是春季、秋季水体TLIc 升高的决策因子。
为探究这些决策和限制因子对水体TLIc 的响应机制,有必要深入分析其直接作用系数、间接作用系数的绝对值的大小,以及相互间的作用途径。 由表6 可见,影响春季、夏季水库水体TLIc 的直接作用系数中,绝对值最大的因子均为x6,作用系数分别达-0. 68、81. 40。 但在春季,x6通过x3对TLIc 发生较大的间接正向作用,间接作用系数总和达0. 57,致使春季x6对TLIc 的综合作用效果最小,相关系数仅为-0. 11。 在夏季,x6通过x3对TLIc 发生较大的间接负向作用,间接作用系数总和达-81. 14,导致夏季x6对TLIc 的综合作用效果与x3相当,相关系数均为0. 26。 在春季,x7对TLIc 的决策系数为较大的正值,且偏相关系数r(y,x7) =0. 519 9(P=0. 001 0),达到极显著水平,因此,x7为春季水体TLIc 升高的决策因子。 在春季和夏季,x6对TLIc 的决策系数均为较小或最小负值,但春季r(y,x6) = -0. 298 8(P= 0. 072 4),未达到显著水平,因而,x6构成春季水体TLIc 波动的限制因子不显著; 而夏季r(y,x6) =-0. 352 7(P=0. 032 3),达到显著水平,因而,x6构成了夏季水体TLIc 波动的限制因子。
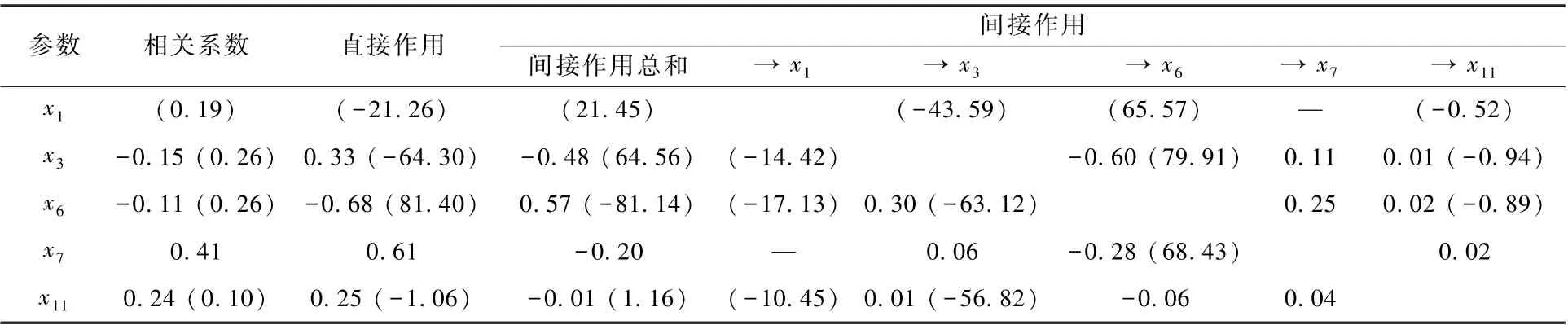
表6 影响春季和夏季水库水体TLIc 变动的主要食性功能群参数的相关与通径分析Table 6 Correlation and path analysis between TLIc value and the main feeding functional groups in the reservoirs in spring and summer
对秋季水体TLIc 影响最大的直接作用因子是x3,作用系数为0.388(表7),但其通过x7对TLIc 发生了较大的间接负向作用,致使x3对秋季水体TLIc 的综合作用效果并非最大。x13的直接作用系数虽最小,仅为0.250,但其通过x3、x7对TLIc的间接作用总和为正向,从而导致x13对秋季水体TLIc 的综合作用效果最大,相关系数为0.253。x13的决策系数虽为最大的正值,但r(y,x13)= 0.270 2(P=0.111 0),未达到显著水平,因此,x13成为秋季水体TLIc 升高的决策因子不显著。
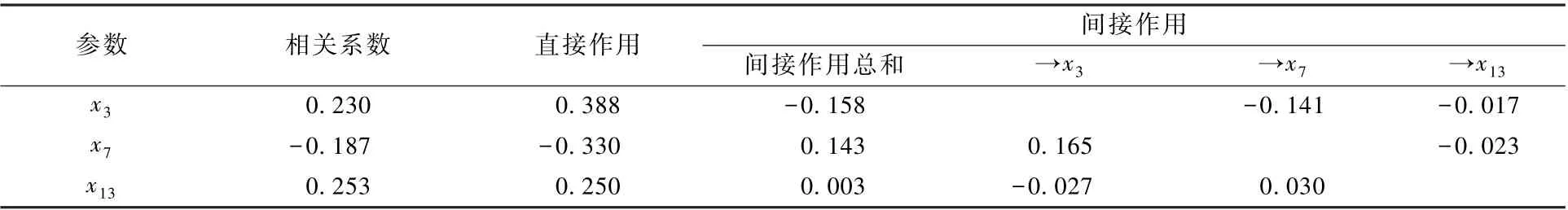
表7 影响秋季水库水体TLIc 变动的主要食性功能群参数的相关与通径分析Table 7 Correlation and path analysis between TLIc value and the main feeding functional groups in the reservoirs in autumn
对冬季水体TLIc 影响最大的直接作用因子为x10,作用系数为-3.787;其次为x2,作用系数为3.733(表8)。 但x2、x10可通过相互之间的生态竞争作用分别对TLIc 产生较大的间接负向、间接正向作用,致使x2、x10对TLIc 的综合作用效果大幅下降, 两者与 TLIc 的相关系数分别仅为-0.112、-0.149。x2、x10对TLIc 的决策系数的负值均较小,且两者差异不大,r(y,x2) = 0.330 0(P= 0.046 1),r(y,x10) = - 0.330 5 (P=0.045 7),均达到显著水平,因此,x2、x10共同构成了冬季水体TLIc 波动的限制因子。 而x1对TLIc 的决策系数虽为较大正值,但r(y,x1) =-0.218 8(P= 0.193 3),未达到显著水平,故x1不是冬季水体TLIc 波动的决策因子。
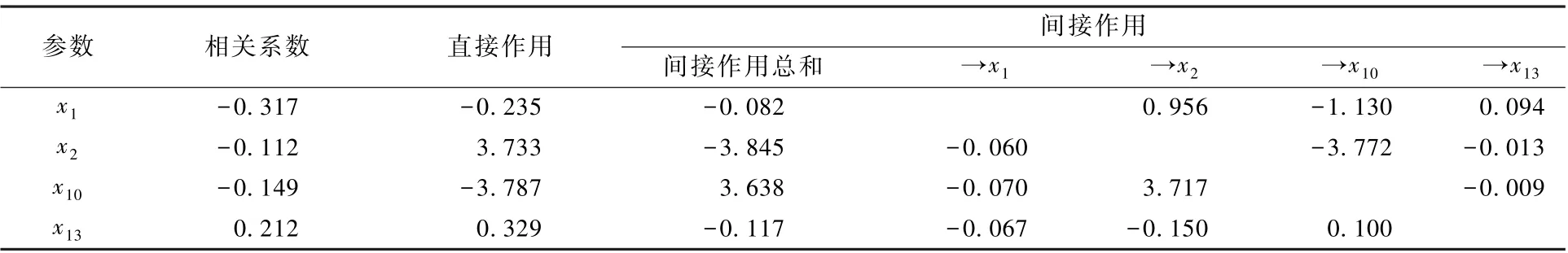
表8 影响冬季水库水体TLIc 变动的主要食性功能群参数的相关与通径分析Table 8 Correlation and path analysis between TLIc value and the main feeding functional groups in the reservoirs in winter
3 讨论
3.1 滤食轮虫的丰度和生物量及水质响应
随着渔业生产的发展,水库浮游动物小型化趋势明显,轮虫的数量不断上升,甲壳类浮游动物的数量不断下降[22]。 滤食轮虫以细菌、藻类和有机质为食。 一方面,在温度、食物条件得到改善的春季,滤食轮虫会大量繁殖,有些轮虫还具备反捕食的形态特征、行为特征或生活史对策[23],从而使x1参数很快上升;但另一方面,生物量信息往往更能反映该功能群的生态积累过程,即滤食轮虫不断消耗水体中的浮游植物、有机微粒等营养物质,将其转化为自身物质贮存起来并参与能量流动。 因此,x7参数能更好地反映春季水库水体营养水平,是TLIc 升高的决策因子(表6)。 另外,春季是小型滤食动物开始生长繁殖的季节,此时其丰度较低,x3参数仅为夏季峰值的12.4%(表4)。 可见,x6通过x3间接促进春季水库水体TLIc 升高的幅度有限,这可能就是x6未能构成春季水体TLIc 波动的限制因子的原因。
3.2 小型滤食动物的丰度及水质响应
小型滤食动物的丰度占浮游动物总丰度的64.5%,生物量占76.1%,是水库中最重要的功能群。 常见种有长额象鼻溞(Bosminalongirostris)、颈沟基合溞(Bosminopsisdeitersi)、尖额溞,幼年期的短尾秀体溞(Diaphanosomabrachyurum)、僧帽溞(Daphniacucullata) 和微型裸腹溞(Moina micrura),以及哲水蚤桡足幼体(copepodids of Calanoida)、 剑 水 蚤 桡 足 幼 体( copepodids of Cyclopoida)和无节幼体(Nauplius)等。 一年四季中,春、夏、秋3 个季节的x3参数均入选了水质预测模型回归方程。 在夏季,食性相同的滤食轮虫、小型滤食动物的丰度和生物量均达到了峰值(表4),可见这两类重要的浮游动物食性功能群在夏季存在竞争排斥。 这与LERUSTE 等[24]研究浮游植物功能群时得出的“一些重要类群在夏季存在竞争排斥”的研究结论类似。 通径分析表明,在夏季,x6主要是通过x3的增大促使水体营养水平下降,并成为水体TLIc 波动的限制因子(表6);在秋季,x3是促进水库营养水平升高的最大直接作用因子,同时,x3也可通过x7(食物等生态资源竞争)间接促使TLIc 下降(表7)。 另外,秋季浮游动物的种类多、丰度大,x13对TLIc 的直接作用、间接作用总和虽均为正值,但这种促进作用并不显著。 这或许就是秋季水库营养水平不能用x13来评价的原因之一(表7),况且在处于中营养水平的水源地水库,x13是随着水体营养水平的升高而增大的。 究其原因,还是传统的生物多样性指数没有对所有物种在体型、形态发育和摄食行为等方面进行区别对待[25]。
3.3 中型滤食动物的生物量及水质响应
中型滤食动物如成体短尾秀体溞、僧帽溞、微型裸腹溞、汤匙华哲水蚤等,其生物量数据比丰度更大,因而包含了更多的生态过程信息。 中型滤食动物的生物量占浮游动物总生物量的9.1%,仅次于小型滤食动物(76.1%)。 在冬季,x10和x2共同构成了水体TLIc 波动的限制因子,其作用方式相互依赖、作用效果相互对立,可通过彼此的相互作用间接促使TLIc 上升或下降(表8)。 这可能缘于它们属于生存策略完全不同的生态功能群:中型滤食动物以滤食细菌、藻类、有机质和原生动物为生,可净化水质,有一些种类还可作为水体寡污性指示种类,如老年低额溞(Simocephalus vetuloides)和长刺溞(Daphnialongispina)等[26];捕食轮虫主要为肉食性的晶囊轮虫(Asplanchna),多见于α-中污带至β-中污带水体。 安睿等[7]在观察三环泡湿地浮游动物功能群时发现,中型滤食动物与捕食轮虫功能类群的生物量之间存在一定的负相关性。 本研究发现,这两种功能群的浮游动物的竞争排斥发生在冬季,此时的x2最大,而x10最小(表4)。 值得注意的是,无论是丰度还是生物量,冬季不同水库的捕食轮虫与中型滤食动物、中型捕食动物之间均存在着极显著的正相关性(n=40,r≥0.901,P<0.01)。 肉食性的晶囊轮虫会选择性地捕食带卵轮虫[23],但关于捕食轮虫是否也能通过捕食中型滤食动物、中型捕食动物的卵或幼体,从而成为浮游动物种群相对稳定的内部调节者,还有待于进一步观察。
中型捕食动物生物量数据包含的生态信息也比其丰度更大,故x11入选了春、夏季的水质模型方程。 春季的主要种类为许水蚤(Schmackeria),夏季的主要种类为广布中剑水蚤(Mesocyclops leuckarti)、近邻剑水蚤(Cyclopsvicinus)和台湾温剑水蚤(Thermocyclopstaihokuensis)。
综上所述,x7是春季水体TLIc 发展的决策因子;x6构成了夏季水体TLIc 波动的限制因子,主要是通过x3的增大促使水体TLIc 下降;x13促进秋季水体TLIc 升高的作用不显著;x2和x10共同构成了冬季水体TLIc 波动的限制因子,两类功能群可通过相互间的竞争间接促使TLIc 升高或降低。 明确浮游动物食性功能群参数与TLIc 的关系及相互作用机制,可为基于浮游动物食性功能群参数的水源地水质监测工具的开发提供新的思路。
4 结论
浙江省水源地水库浮游动物的丰度和生物量均以小型滤食动物为最高,滤食轮虫(丰度)和中型滤食动物(生物量)次之。 多元回归与通径分析表明:小型滤食动物丰度分别入选了春、夏、秋3 个季节的水库水体TLIc 多元回归方程,且总浮游动物丰度成为春、夏季水体TLIc 波动的限制因子,均通过小型滤食动物丰度的间接作用实现;滤食轮虫丰度和中型滤食动物生物量均入选了冬季水库水体TLIc 多元回归方程,且后者又是冬季水库水体TLIc 波动的限制因子。 通过合理的浮游动物功能分群和多参数数据统计分析,能获得较好的水体综合营养水平模型。
