滇水金凤SAUR基因的克隆及表达分析
2022-03-09童桢开李新艺黄美娟黄海泉
童桢开,李 洋,蔡 斌,李新艺,黄美娟,黄海泉
(西南林业大学 园林园艺学院,国家林业和草原局西南风景园林工程技术研究中心,云南省功能性花卉资源及产业化技术工程研究中心,西南林业大学园林园艺花卉研发中心,昆明 650224)
滇水金凤(ImpatiensuliginosaFranch.)是中国所特有的凤仙花科凤仙花属一年生草本植物,整株无毛,花色以红色为主,斜卵形,旗形花瓣圆形,花瓣延长而形成的漏斗状管状结构为花距[1]。花距是植物重要的观赏特征之一。迄今为止发现滇水金凤的野生种有着多种花距形态,如短距、直距和长距等[2]。花距可以使高级物种的形成率变高,同时花距可以选择自己需要的传粉者,避免无用传粉者浪费花蜜。对花距的研究有助于推断物种形成和进化机制,有助于理解植物与传粉者个体之间的相互作用,以及复杂的植物-传粉者的群落机制。此外,花距还有调节生物入侵的重要功能[3]。但是到目前为止,除了有研究表明对花距发育有调控作用的KNOX基因外[4],对于促进花距伸长和改变的调控基因还是未知。
生长素(auxin,indole acetic acid,IAA)是最早被鉴定出的一类植物激素,几乎在植物生长发育各个阶段都存在IAA响应基因的表达调控[5-6]。早期应答生长素基因(small auxin up RNA,SAUR)是植物IAA早期响应基因中最大的家族,1987年首次在IAA诱导处理的大豆下胚轴中发现SAUR表达[7-8]。目前,已经在许多植物中鉴定出多个SAUR基因家族成员,例如拟南芥72个SAUR基因、水稻58个SAUR基因、玉米91个SAUR基因等[9-11]。由于国际上对凤仙花属植物的研究起步较晚,且滇水金凤具有明显的地域性,目前国内外关于滇水金凤的研究很少,主要有林琼等[12]发现滇水金凤种子具有休眠特性,Vieira等[13]首次报道了凤仙花中黄酮类化合物的成分等。关于具有调控花距生长作用的SAUR基因的相关研究比较罕见。本研究对滇水金凤花距发育SAUR基因进行鉴定和分析,旨在探明SAUR对花距生长发育调控的分子机制,为后续研究凤仙花属植物花距的生长发育调控机理奠定基础。
1 材料和方法
1.1 试验材料
滇水金凤(ImpatiensuliginosaFranch.)来源于昆明市捞鱼河湿地公园及西南林业大学后山实验大棚,取滇水金凤花距器官,以滇水金凤花苞期、花苞开放期、盛花期3个时期花的花苞(图1),取花距尖部、弯部、基部和檐部等4个部位进行试验。
1.2 方 法
1.2.1 总RNA的提取与检测提取不同时期不同部位的RNA,并在课题组前期研究基础上设计出6对SAUR基因特异性引物(表1),把RNA通过逆转录得到SAUR基因cDNA。另提取的滇水金凤DNA核酸,以其为模板扩增得到滇水金凤SAUR基因gDNA。PCR产物经纯化后进行连接反应,经转化、酶切和PCR鉴定后,结果与预期目的片段均相符者,送上海生工生物有限公司进行测序。
1.2.2SAUR基因的生物信息分析运用ExPasy-ProtParam(https://web.expasy.org/protparam/)软件进行SAUR基因编码蛋白质的基本理化性质分析;采用SignalP-4.1(http://www.cbs.dtu.dk/services/SignalP-4.1/)进行蛋白序列信号肽预测;采用GSDS(http://gsds.cbi.pku.edu.cn/)进行基因内含子和外显子预测;采用Protein BLAST(https://blast.ncbi.nlm.nih.gov/Blast.cgi)及DNAMAN软件进行基因的同源比对分析;运用软件MEGA 6.0构建系统发育树。
1.2.3 滇水金凤SAUR基因的表达分析(1)RNA提取和反转录 提取滇水金凤3个时期(花苞期、花苞开放期、盛花期)花距的檐部和距部,及盛花期花距的尖部、弯部、基部和檐部的RNA,逆转录成cDNA备用。
(2)引物设计 根据已获得的6个SAUR基因的cDNA全长序列,设计相关引物(表1)。
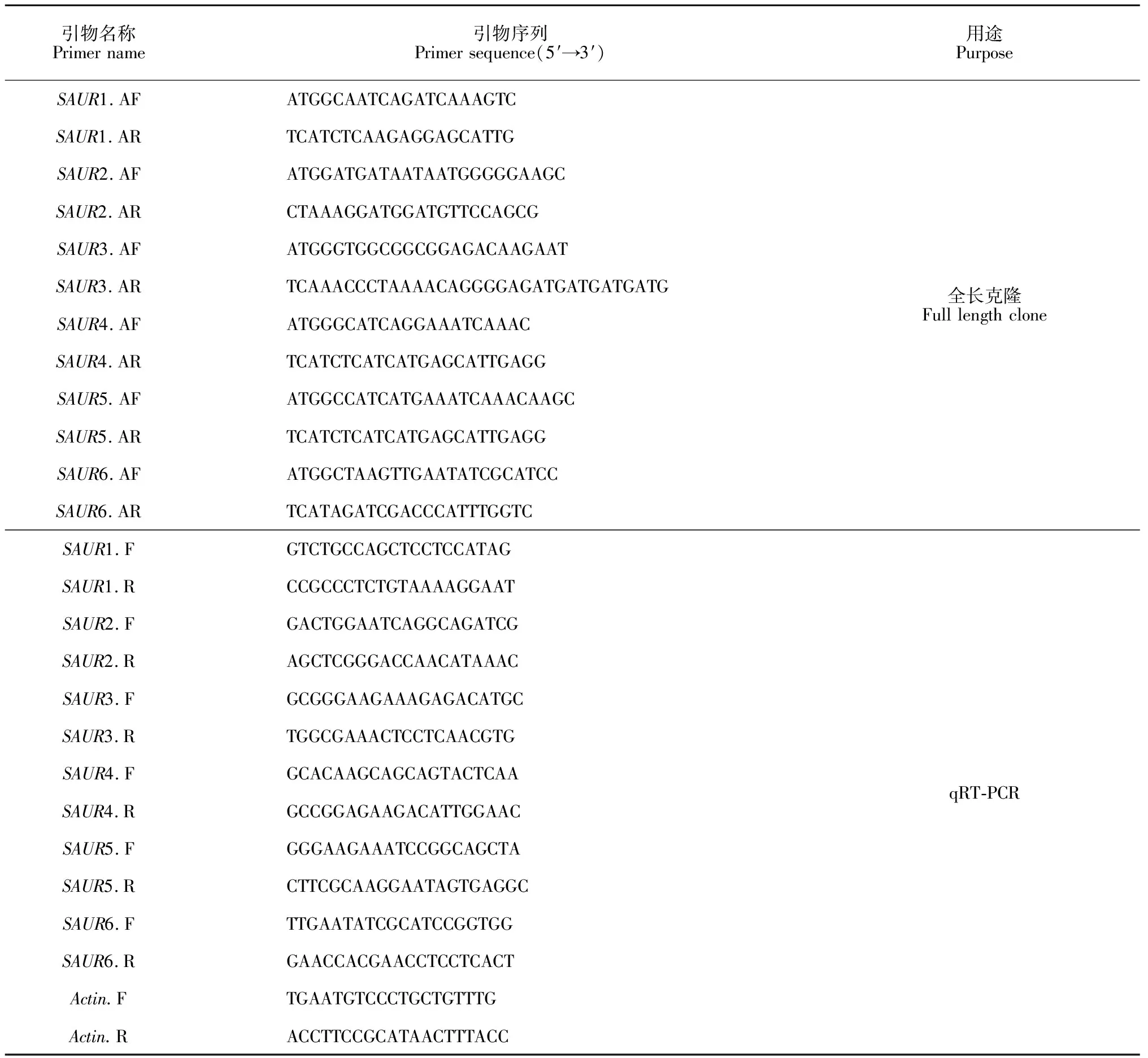
表1 滇水金凤SAUR基因的引物序列
(3)荧光定量PCR 根据qRT-PCR的结果记录下每组数据的Ct值,每组试验重复3次。利用2-ΔΔct计算方法分析不同花距部位基因相对表达量。再对滇水金凤SAUR基因的相对表达情况进行分析和制图,将花苞期作为3个时期的对照组,将花距尖部作为2个花距的对照组,将其统一定为1个单位。
2 结果与分析
2.1 SAUR基因总RNA提取及DNA扩增
对滇水金凤总RNA提取结果测定浓度,其28S、18S rRNA条带清晰,且二者亮度约成2倍关系,其提取质量满足后期试验要求(图2)。
结果表明,SAUR1、SAUR2、SAUR3、SAUR4、SAUR5和SAUR6各基因都位于300~500 bp,且均为单一条带,其中cDNA序列全长分别为351、534、396、333、309和411 bp(图3,A)。
以滇水金凤DNA为模板,采用扩增引物(表1)进行全长gDNA PCR扩增,通过电泳检测,得到的SAUR1、SAUR2、SAUR3、SAUR4、SAUR5和SAUR6基因的目的条带位于300~500 bp,且均为单一条带,说明是特异性扩增(图3,B)。
2.2 滇水金凤SAUR基因编码蛋白的理化性质分析
对SAUR基因编码蛋白质的基本理化性质分析结果显示(表2),根据各基因编码蛋白质的不稳定指数推测,除SAUR3基因编码蛋白质的不稳定指数(instability index)为37.30(<40)属稳定蛋白外,其他5个SAUR基因编码蛋白质的不稳定指数都大于40,皆为不稳定蛋白。对滇水金凤SAUR蛋白序列进行信号肽预测结果表明,所有氨基酸肽链上均不存在信号肽,因此滇水金凤SAUR蛋白均属于非分泌蛋白。对滇水金凤SAUR基因编码蛋白质的亲/疏水性分析结果显示(表3),除SAUR5蛋白的比例较低为-1.478,且总平均亲水指数为0.045,属于疏水性蛋白外,其余SAUR蛋白的比例较高,且总平均亲水指数均为负值,属于亲水性蛋白。对滇水金凤SAUR基因内含子和外显子进行分析,6个滇水金凤SAUR基因家族成员均不含内含子;对滇水金凤SAUR蛋白的功能结构域分析,6个 SAUR蛋白均属于生长素诱导的超家族成员,其中SAUR3含有稳定的PLN3219结构域,其余5个SAUR蛋白均具有稳定的PLN3090结构域。

表2 滇水金凤SAUR基因编码蛋白的基本理化性质

表3 滇水金凤SAUR蛋白亲/疏水性指标
2.3 滇水金凤SAUR基因系统关系分析
对包含滇水金凤、石榴、马铃薯等34个物种的氨基酸序列和系统亲缘关系进行分析,结果(图4)显示:滇水金凤SAUR1、SAUR2、SAUR3、SAUR4、SAUR5和SAUR6基因的氨基酸序列与其他植物的相似性分别为73.50%、65.68%、63.47%、82.80%、75.80%和47.34%,其中,SAUR6基因与其他6种植物的相似性最低为47.34%。SAUR4基因与其他5种植物的相似性为82.80%。而亲缘关系树包括3个分枝,SAUR1、SAUR4和SAUR5处于一个分枝,SAUR3和SAUR6处于一个分枝,SAUR2单独处于一个分枝中。其中SAUR4和SAUR5处于同一个小分枝,推测其为旁系同源;而SAUR3和SAUR6在同一个分枝中的2个单独分枝,推测其为直系同源。
2.4 滇水金凤花距发育过程中SAUR基因的表达分析
2.4.1 不同时期距部及檐部的表达滇水金凤6个SAUR基因在不同时期距部及檐部的表达情况如图5。在距部,以花苞期为参照,随着花开放时期的变化,基因SAUR1、SAUR3和SAUR5的相对表达量先上升后下降,而基因SAUR2、SAUR4和SAUR6则是先下降后上升。基因SAUR3在始花期和盛花期相对表达量分别为花苞期的5 878倍和5 694倍,有显著性差异(P<0.005)。
在檐部,以花苞期为参照,随着花的开放,所有6个SAUR基因的相对表达量均呈现出先上升后下降的趋势,且SAUR3、SAUR5和SAUR6基因始花期檐部的相对表达量均有较大上调,分别是花苞期的216倍、164.4倍和194.5倍,无显著性差异。
从图5不同时期不同部位的表达分析可知,相对始花期和盛花期,花苞期是表达量较低的一个时期;在距部(图5,A),SAUR1、SAUR3和SAUR5基因在始花期发挥作用,SAUR2、SAUR4和SAUR6基因则在盛花期发挥作用;而在檐部(图5,B),所有基因均在始花期发挥作用。推测在檐部SAUR基因于始花期促进细胞的分裂和伸长,而到了盛花期作用急剧减缓。
2.4.2 不同部位始花期及盛花期的表达滇水金凤6个SAUR基因在始花期及盛花期不同部位中的表达情况如图6。在始花期(图6,A),以花距尖部为参照,通过相对表达量可见,SAUR1-2基因在4个部位不表达或表达量很少;SAUR3基因弯部相对表达量约为6.5倍,其他部位表达量很少;SAUR4基因在4个部位均有一定量表达;SAUR5基因在4个部位均有一定量表达,檐部显著增长,基部表达量高至36.0倍,有显著性差异(P=0.029);SAUR6基因在基部有较高表达量,在其他部位少量表达。
在盛花期(图6,B),以花距尖部为参照,通过相对表达量可见,SAUR1-3基因在4个部位表达量很少或不表达;SAUR4-6基因在几个部位中相对表达量均较高,以SAUR5和SAUR6基因最高,尤其是SAUR6基因在檐部相对表达量高达339.4倍,有显著性差异(P=0.013);6个SAUR基因在尖部都有较稳定的表达。
从图6不同时期不同部位的表达分析可以看出,在始花期,除了SAUR5和SAUR6在花距基部有较高的表达,以及SAUR3花距弯部表达量较高外,其他部位各基因表达量并无太大区别;而在盛花期,SAUR5和SAUR6基因在花距各个部位都有着非常高的表达。推测在盛花期SAUR5和SAUR6基因在花距各个部位促进细胞的分裂和伸长。
3 讨 论
本研究从滇水金凤花距中克隆出6个SAUR基因特异全长,6个SAUR基因均不含内含子,这与已报道的SAUR基因大多不含内含子结论一致[14-15]。仅在部分物种中有不同,如拟南芥中有1个SAUR基因含有内含子,玉米的58个SAUR基因中有6个,番茄的99个SAUR基因中有3个,马铃薯SAUR基因中的9个,柑桔的70个SAUR基因中有10个含有内含子[7-8, 16-17]。研究表明,无内含子基因的选择性剪接发生率通常较低,某些SAUR家族基因的功能可能是稳定的基因[18],是否说明滇水金凤中SAUR家族基因相较其他物种更为稳定,这为后续探讨该基因在凤仙花属植物中的表达模式及分子机理提供了一定参考。
大部分SAUR基因在启动子区域含有几个生长素反应性元件(AuxRE)[19]。大多SAUR蛋白包含的SSD特异性结构域约60个氨基酸残基,主要由疏水氨基酸组成,高度保守[20]。本研究发现,除SAUR3为稳定蛋白外,其他5个滇水金凤SAUR蛋白成员均为不稳定蛋白,除SAUR5为疏水性蛋白外,其余的5个均为亲水性蛋白。所有基因都属于生长素诱导的超家族成员,在对滇水金凤6个SAUR蛋白的研究中发现,除了SAUR3含有稳定的PLN3219结构域,其余5个SAUR蛋白均有着稳定的PLN3090结构域,SAUR所有成员在蛋白结构上的高度保守可能对其生物学功能的发挥有重要作用,这与Kant S[21]所报道的一致。
滇水金凤6个SAUR基因的氨基酸序列分别与34种如罂粟、石榴、大麻等植物氨基酸序列进行对比分析,具有较高的相似性和种属保守特征,但不同SAUR基因即使在同一物种内也会存在较大差异[22]。本研究构建SAUR基因系统发育树中发现,SAUR1与罂粟处在一个小分枝中亲缘关系较近。SAUR4和SAUR5处于一个小分枝中,与大麻亲缘关系比较近;SAUR2与咖啡亲缘关系较近;与SAUR3亲缘关系较近的是木薯、橡胶树和大叶栎,而与SAUR6亲缘关系较近的是水蜜桃、葡萄和澳洲棉等。有研究对植物SAUR基因的进化进行分析,发现苔藓植物可能是植物的祖先,而SAUR基因在单子叶植物和真子叶植物中独立存在[23]。研究为凤仙花属植物属内系统进化、亲缘关系及种属分类提供了一定的基础数据。
SAUR的功能仍然是神秘的,前期有研究表明许多SAUR基因对盐、干旱和稻瘟病有响应。SAUR69通过抑制生长素的极性运输来提高番茄果实对乙烯的敏感性,从而影响生长素-乙烯化的转变,揭示了SAUR69介导了生长素和乙烯之间的关系[24]。SAUR26亚家族被鉴定为生长热响应性,显示与温度相关的自然多态性,影响植物对当地温度适应的热效应[25]。而本研究就SAUR基因对滇水金凤花距不同时期、不同部位的研究表明,细胞的分裂和伸长可能主要发生在始花期的檐部,且基因SAUR3发挥着非常重要的作用。而在水稻柱头中也发现了OsSAUR54基因可能促进花粉管生长,在棉花中发现GhSAUR33同样具有类似的功能[18, 26]。同时本研究中基因SAUR在檐部的表达模式可能是在始花期促进了细胞的分裂和伸长,而到了盛花期作用急剧减缓。从2个时期4个部位的表达分析中推测,SAUR基因对花距的细胞的分裂和伸长的作用主要发生在基部和檐部;同时在不同时期花距不同部位中均检测到了SAUR基因的表达,表明SAUR基因参与了植物生长和应激反应的多个过程,为滇水金凤花距发育、物种形成和进化研究提供了一定的基础数据和理论依据。
