水稻种子耐贮性研究进展
2022-03-02闫蕴韬张海清贺记外
闫蕴韬,何 兮,张海清,贺记外
(湖南农业大学农学院,长沙 410128)
0 引言
水稻在贮藏过程中,种子内部会自发地发生一系列生理变化,如胶体陈化,导致蛋白质凝固、活力丧失,营养物质氧化,导致种子活力降低、成苗率下降等现象[1]。中国水稻生产、贮藏、消费地区主要分布在南方高温高湿地区,现阶段水稻种子贮藏仓库主要是通过控制贮藏温度和湿度以及保持适宜的通气条件从而达到延缓种子衰老和稻谷质变的进程,例如建立充满惰性气体、低温干燥的密闭粮库等,但这些贮藏方式需要消耗大量的能源,同时也造成了许多环境问题,水稻种子贮藏面临严峻的问题[2]。水稻种子自身耐贮性的好坏在一定程度上决定了水稻种子和粮食储藏能力的强弱。因此,对水稻种子耐贮性进行研究具有重要的意义[3]。
种子耐贮性是指完成后熟的种子经过长时间的贮藏后仍然能够保持最原始的种子活力的状态。水稻种子在贮藏的过程中,种子内部不可避免地发生一系列的生理变化,导致种子活力下降或丧失,进而会对农业生产带来重大损失。具有较强耐贮性的种子可以确保种子长期贮藏后仍具有较强的活力与生活力,并保证水稻种子的田间经济产量。如何在水稻种子的贮藏过程中减少内部营养物质的消耗,有效延缓种子衰老和种子劣变是亟待解决的问题。目前科研工作者已经定位到了部分与水稻种子耐贮藏相关的QTL位点,但真正进行功能研究的基因不多。科研人员从水稻种子耐贮藏的调控机制出发,在水稻种子的自我保护机制以及大分子损伤修复机制的过程所涉及到的一些蛋白与基因进行功能分析,从而确定了这些基因对水稻种子耐贮性强弱有重要的意义,但是具体的调控机制不是十分明晰。为了进一步探究种子耐贮的生理生化机制与遗传因素,解析种子耐贮性的分子机理,本研究在已有的研究基础上对可能影响水稻耐贮性的相关基因与蛋白进行综述,对科学探究种子耐贮性具有指导意义。
1 种子劣变与种子老化
种子劣变是致使种子活力降低的不可逆的现象,在种子贮藏过程中无法避免[4]。种子在自然条件下质量衰退以及活力降低的过程称为老化,种子老化的进程受外部的高温高湿环境的影响。种子的老化与劣变同时发生,有老化就有劣变,但种子劣变并不一定由种子老化所引发[5]。劣变与老化是引起种子活力降低的主要因素,在种子收获后至生理成熟,以及后期种子贮藏的过程中都伴随着老化与劣变[6]。Delouche和Caldwell[7]研究了种子劣变过程中种子活力及种子发芽力两者的关系,种子活力与种子生活力呈线性正相关,种子劣变的发生会降低种子活力[8]。然而,由于种子自身的一些特性的存在,种子硬实和种子休眠会对种子老化产生不同的响应机制。有关研究表明,硬实的种子贮藏过程中具有较小程度的劣变,胡贵江[9]通过对45℃与42℃两种不同温度下的棉籽进行72 h的老化处理后发现较高的温度可以更好的破除种子的硬实使其发芽,且种子硬实率与老化处理时间呈负相关。在种子可接受的范围内,随着适当种子老化的湿度的增加或者老化的时间的延长,可以有效促进水稻种子萌发,表明了人工加速老化可以有效破除硬实的种子休眠[10]。
2 种子耐贮性的生理生化机制与生物大分子损伤机制研究
种子完成生理成熟后,内部生命活动的进行仍伴随着种子老化和劣变。老化劣变机制是涉及种子生理生化变化和种子新陈代谢等一系列复杂的调控机制,目前对这一机制的研究尚不清晰,生理生化及细胞学方面的研究表明导致贮藏期种子老化的因素有:脂膜过氧化与膜系统损伤,蛋白质合成能力下降与蛋白质损伤,内源激素及有毒物质的积累,抗氧化能力下降,DNA损伤等。Pillay[11]提出了老化劣变起因的多种假设,主要包括:营养物质损耗的假说;激素变化的假说;有毒物质积累的假说;生物大分子变性的假说以及功能结构如膜、线粒体解体的假说;Bewley等[12]提出种子贮藏期间DNA与RNA的降解以及种子内部氧化还原稳态也与种子的耐贮性相关。在种子发芽率下降,幼苗生长势降低之前,种子内部会发生一系列的生理变化,张兆英[13]研究表明种子的老化或种子劣变主要表现在细胞膜完整性的丧失和细胞内部脂类物质氧化程度的加深,细胞内各种酶活性的降低,呼吸作用也会受到一定程度的破坏,种子内部营养物质不断消耗,以及ABA、GA含量的变化。Delouche[7]预测了种子在劣变过程中发生变化的生理生化顺序为生物膜系统的降解,呼吸作用与营养物质的消耗,种子抗逆性变弱,种子耐贮能力下降,种子田间成苗能力减弱等。
2.1 膜脂过氧化与膜系统损伤
细胞质膜由不连续的磷脂双分子层构成,在磷脂双分子层上附着部分球状蛋白。这些球状蛋白在保护细胞内部物质的完整性以及调节各种物质出入细胞膜方面具有重要的作用。Agmon等[14]提出了关于脂质过氧化如何导致细胞死亡的4个假说。第一个假说:脂质过氧化引起的细胞成分的变化会造成大范围膜损伤,使膜上的洞孔打开,导致离子稳态的丧失。第二个假说:由于脂质的化学变化,破坏了细胞膜完整性,破坏了细胞膜结构。第三个假说:主要造成膜损坏的不是膜内部含有的脂质成分,而是膜内蛋白。第四个假说:氧化产生了不饱和脂肪酰胺片段,这些片段会对其他细胞的正常生命活动造成损害并可能致死。
颜启传等[15]发现水稻种子浸出液中可溶性糖的电导率和含量指数随着种子老化程度的加深而增加,说明种子老化与细胞膜通透性有关,种子老化程度越高,种子膜系统的损害越严重。
2.2 蛋白质合成能力下降和蛋白质损伤
Pilly[11]发现大豆种子蛋白质合成能力下降导致种子劣变。Rajjou等[16]研究发现使用蛋白质抑制剂放线菌酮处理后的拟南芥种子很难发芽,认为加速老化后种子活力的下降与蛋白质合成能力密切相关。蛋白质共价键自发损伤也是种子老化的主要原因之一,L-异天冬氨酸-甲基转移酶(PIMT)是一种依赖于S-腺苷甲硫氨酸的重组酶,种子老化中异天冬氨酸产生自发性残留物致使蛋白质损伤[17],而PIMT可修复其蛋白质损伤。与DNA修复相似,蛋白质修复也是对已损伤的蛋白质结构进行溯原,在拟南芥中发现当蛋白质L-异天冬氨酸-甲基转移酶I(Protein L-isoaspartylmethyltransferase,PIMT1)过度积累时,拟南芥体内异天冬氨酸的含量减少,种子寿命与种子活力增强,相反PIMTI累积减少时,种子寿命缩短,种子活力下降[18]。水稻中的OsPIMT1缺失可以降低老化后种子的发芽率,在老化的过程中OsPIMT1编码的PIMT修复酶通过减少体内有害蛋白的积累来维持种子胚胎的活力,从而提高老化后种子的寿命与活力[19]。
2.3 内源激素及有毒物质的变化
种子老化的过程中,伴随着内源激素含量的变化,老化后的细胞及生理系统对激素的反应能力也会丧失。ABA(Abscisic acid)、GA(Gibberellins acid)、IAA(Indole-3-acetic acid)作为植株生长的主要调控激素在调控种子的成熟、发芽的过程中具有不可或缺的作用,对种子的休眠与耐贮性具有重要的影响。付华等[20]以4种牧草种子为研究材料,探究了不同老化处理对种子萌发过程中乙烯释放量的影响和乙烯释放量与发芽率的关系,结果表明,随着种子劣变程度加深,乙烯释放量逐渐下降,释放高峰推迟,老化过程中的代谢产物如乙醇、醛、酮等有毒物质的积累会对破坏种子的正常生理活动,最后导致种子丧失生命力。
2.4 抗氧化能力下降
在种子发育、成熟、加工与贮藏以及萌发期间会遇到各种逆境胁迫,胁迫因子会对种子的品质造成很大的影响,进而对作物生产造成不可逆的影响[21]。细胞内活性氧(ROS)状态的失衡和随之而来的氧化损伤与种子生存能力的丧失有关。在种子劣变的过程中,ROS积累、脂肪酸过氧化所产生的代谢物质,会对细胞的结构与物质如:细胞膜、核酸、蛋白质大分子物质、碳水化合物、脂质造成不可逆的损害[22]。
2.5 DNA损伤
一般情况下,种子活力高低和出芽速度依赖于种子吸胀吸水过程中激活的DNA修复机制,同时种子劣变发生伴随着染色体畸变、端粒长度改变、DNA损伤、DNA甲基化[23]。
种子体内DNA损伤主要来自氧化应激反应[24],种子吸胀萌发时伴随着ROS诱导的DNA损伤,导致种子劣变。DNA损伤会直接损害细胞的功能,在种子内部发生的DNA单碱基突变有可能遗传给后代。拟南芥的ATOGG1是一种DNA裂解酶,参与碱基切除修复以消除DNA中的8-OXOG基因。该基因的过量表达抑制种子劣变的发生,转基因种子中8-OXOG表达量降低,可以在一定程度上修复损伤的DNA,使种子在非生物胁迫下表现出较强的萌发能力[25]。DNA甲基化在种子生育和生存能力的保持上有很重要的作用[26]。RNA也会被ROS破坏,由于其单链结构和缺乏修饰系统,更容易被氧化[27]。氧化的mRNA会抑制蛋白质的合成和降解,因此进行生命活动的种子细胞在mRNA转录水平上受到了阻碍,导致种子寿命降低。
3 水稻种子耐贮性的遗传学研究
3.1 水稻种子耐贮性QTL定位
种子活力与自身发育、贮藏条件等外界因素的影响密不可分,同时也受遗传因素的调控。随着当今DNA分子标记技术和基因组作图技术的发展,越来越多的与种子活力和耐贮相关的QTL被检测到。Li等[28]在南京、连云港、金湖等不同的种植环境下种植‘Koshihikari’(轮回亲本)和‘Kasalath’构建的回交重组自交系(BIL)群体,在水稻9号染色体上R10783S与R1751两标记之间检测到了种子耐贮性QTL位点qSS-9,该位点解释表型变异为10.63%~33.10%,通过染色体片段置换(CSSL)群体验证了该位点。Miura[29]最先在9号染色体G103与R1751区域检测到与种子寿命相关的主效QTL位点qLG-9。随后不久,Sasaki等[30]、Xue等[31]、Jiang等[32]通过构建不同的遗传群体均在9号染色体上检测到了与qLG-9重叠的主效QTL,分别命名为 RC9-2、qRGR-9、qMT-SGC9.1和 qDTSGC9.1。刘喜等[33]利用分子标记技术构建了4个携带不同大小的‘Koshihikari’纯合F4家系,将qSS-9精细定位在Indel标记Y7与Y13之间。沈圣泉等[34]以‘珍汕97B’/‘密阳46’构建的RIL群体作为材料,在温度45℃,湿度100%的条件下对群体材料进行人工老化处理,以老化处理后的发芽率占对照发芽率的相对比值作为衡量指标,检测到了2个耐贮主效QTL,并证明该位点的加性×加性上位性效应大于加性效应。同时还发现根据耐贮的时间的不同,起主效作用的QTL也不同,7天的老化处理检测到基因qSS9-1起主要作用,而在14天处理下检测到四号染色体基因qSS4对增加种子耐贮性起主要作用。此外,Xue等[31]使用‘IR24’/‘Asominori’的重组自交系检测了1、3和9号染色体上与种子耐贮性相关的3个QTL。柳武革等[35]以水稻品种‘广恢122’和籼稻‘G99’杂交后代构建的F2:3群体为研究材料,在9号染色体上定位到2个与贮藏特性相关的发芽率QTL。Zeng等[36]以籼稻品种‘ZYQ8’与粳稻品种‘JX17’进行杂交,收集F1代花粉进行培养,经自然加倍与人工秋水仙素处理加倍后获得染色体片段替换系‘CSSL45’和‘CSSL29’,通过分子标记辅助选择与‘JX17’进行连续3次回交,成功构建加倍单倍体(DH)群体,通过对得到的种子进行人工老化处理在第9,11,12号染色体各检测到一个主效QTL,命名为qLS-9、qLS-11和qLS-12,解释的表型变异分别为12.2%,18.2%和10.6%。其中qLS-9的加性效应为9.21,来自‘JX17’的qLS-9位点可增强水稻种子的耐贮性;qLS-11和qLS-12的加性效应分别为12.30和8.65,来自ZYQ8的qLS-11和qLS-12位点可以增强种子耐贮性。Hang等[37]将‘Sasanishiki’‘/Habataki’‘/Sasanishiki’的回交重组自交系(BIL)群体在自然条件下贮藏两年,经过不同时间的人工老化处理后进行QTL分析,在后代自然老化的条件下检测到6个QTL,大部分QTL位点均与籼稻品种‘Habataki’定位到的QTL位点表现出协同效应。
在自然储存的条件下,Sasaki等[30]用‘Milyang 23’和‘Akihikari’构建的重组自交系群体(F7)的种子分别贮藏1年,2年和3年,定位到4个与种子寿命相关的QTL(RC7、RC9-1、RC9-2和RC9-3)。Jiang等[32]采用类似的处理方法,在两个重组自交系群体中检测到了7个与种子耐贮性相关的QTL。
Li等[28]利用‘Koshihikari’和‘Kasalath’构建的回交群体的种子在自然老化处理32个月和48个月进行种子耐贮性QTL,在第2、3、4、6、9和11号染色体上各检测到1个QTL位点。虽然这些检测到QTL位点能我们对水稻种子耐贮性有了进一步的了解,但是通过比较发现在自然与人工两种处理下检测到的QTL并不完全一致[38]。任淦等[39]通过自然老化和人工老化处理对‘珍汕97’/‘明恢63’衍生的重组自交系群进行了种子耐贮性QTL定位,在室温条件下储存3年条件下检测到3个QTL(qNRP1、qNGR3-1、qNGR3-2),分别位于第1和第3染色体上,且定位于‘珍汕97’的qNRP1和qNGR3-2表现出协同效应,对表型变异贡献率分别达到8.5%和14.5%。在人工老化箱(温度50℃、湿度95%)老化处理72小时条件下,分别在第1、8和9号染色体上定位到了4个QTL,分别命名为qAGP1、qAGP8、qAGR9-1和qAGR9-2。本文在水稻图谱上对前人检测到的水稻种子耐贮性QTL进行了标注(图1)。发现除了10号染色体外,在其他染色体上均检测到了水稻种子耐贮性QTL位点,这表明水稻种子耐贮性是典型且复杂的数量性状。
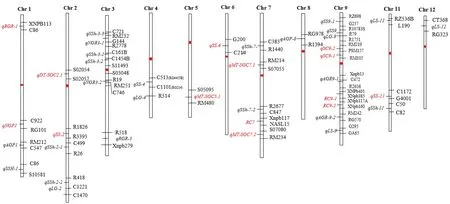
图1 水稻种子耐贮性相关QTLs定位
3.2 水稻种子耐贮性调控机理研究
种子自身的保护机制和生物大分子损伤修复机制是种子耐贮性的主要调控机制[40]。种子发育中后期所具有的脱水耐性可以使种子获得生活力和耐贮性。
种子的成熟过程中,种子体内糖分的积累,热激蛋白(HSP),胚胎发育晚期丰富蛋白(LEC),激素调节相关的基因也是调控种子的耐贮性的关键[41]。糖与激素的相互作用是植物调控自身生长发育的重要途径。Li等[42]通过过量表达ZmGOLS2,ZmRS及AtSTS来提高拟南芥中的棉子糖低聚寡糖(RFO)的含量,极大提高了拟南芥的种子活力,但水稻中棉子糖相关研究尚未见报道。
海藻糖是由两个葡萄糖分子组成的二糖,在植物的抗逆反应中充当信号分子[43]。水稻中仅存在一个海藻糖酶(TRE)基因OsTRE1,该基因同时受低温、高盐、干旱胁迫以及外源ABA诱导,过量表达OsTRE1可以加快海藻糖的降解进程,提高水稻的抗逆性。热激蛋白(HSP)在植物受到逆境胁迫后表达量变高,从而维持细胞稳态,增强植株抗逆性。Kaur等[44]研究表明OsHSP18.2在种子成熟过程中高量表达,在拟南芥背景下过量表达该基因可以减少体内ROS积累,提高拟南芥种子的生活力与耐贮性。HSFA9可以增强植株的抗旱性与氧化胁迫性,并且该基因可以诱导热激蛋白和低聚糖的积累,从而增强种子的耐贮性。Kotak等[45]通过构建拟南芥ABI3敲除株系,证实ABI3通过激活HSFA9启动子从而增强种子的耐贮性。Almoguera等[46]认为HaDREB2和向日葵热激因子A9(HaHSFA9)在向日葵胚胎中共同激活了Hahsp17.6G1启动子,有助于A9介导的种子寿命的增强,并且当HaDREB2与A9共同在种子中过表达时,会显著增强种子的寿命和耐贮性。
水稻种子贮藏过程中受环境的诱导,体内的ROS过度积累会造成生物大分子损伤,增强ROS的清除能力与修复氧化损伤可以使种子生活力与耐贮性显著增强。氧合酶(LOX)是一种属于氧化还原酶家族的非血红素铁的双加氧酶,可在脂肪酸的特定位置氧化不饱和双键产生脂肪酸过氧化物,广泛存在于动植物体中,参与植物的生长发育、成熟、衰老以及植物胁迫相关的反应,也是茉莉酸合成途径的第一个关键酶,在水稻的耐贮方面具有重要的生理功能[47-50]。Suzuki等[51]成功获取了与水稻耐贮相关的材料‘Daw Dam’,证实Lox-3缺失呈单基因隐形遗传。宋美等[52]在人工老化条件下研究了耐贮藏基因(Lox-1、Lox-2、Lox-3)在不同缺失程度下对水稻耐贮性的影响,结果表明Lox-1、Lox-2、Lox-3这3个基因全部缺失种子的耐贮性大于Lox-1、Lox-2两者缺失的种子,大于Lox-2或Lox-3基因缺失的种子,证明了种子活力受耐贮基因缺失程度的影响。吴跃进等[53]利用RNAi干涉r9-LOX1基因,获得转基因水稻植株,通过对转基因后代进行耐贮性分析,证明干涉r9-LOX1基因后的水稻种胚LOX-1酶活性显著降低,显著提高水稻耐贮特性,缓解了水稻陈化变质。刘南南等[54]在栽培稻品种中成功克隆了与种子耐贮相关的基因Lox3,Lox3在胚成熟的过程中会大量表达,在颖壳中的含量很低,主要在种子的胚与表皮中表达。汪仁等[55]成功定位并克隆了LOX缺失基因,成功克隆到编码基因OsLox1和OsLox3,并证实OsLox1所编码的蛋白是一个具有双活性的酶。Cai等[56]发现cryAb/crylAc Bt转基因水稻种子具有较好的耐贮性,在自然条件下贮藏9个月后,转Bt基因抗虫水稻内部的游离脂肪酸含量显著低于受体材料,种子发芽率也显著高于对照材料:Shin等[57]认为水稻乙醛脱氢酶7在调控水稻种子的成熟和保持水稻种子的活力方面具有重要的作用,同时突变体对低温胁迫和盐胁迫比对照更敏感:Oge等[18]研究表明蛋白修复L-异天冬氨酸-甲基转移酶I在延长种子寿命上具有重要的作用,在拟南芥中过量表达PIMT1可以减少种子蛋白组中L-isoaspartyl的积累,从而提高种子的发芽;同样Verma等[58]在鹰嘴豆中发现CaPIMT2通过修复核蛋白中异常的异天冬氨酸残基来修复种子活力与延长种子寿命;水稻存在2个L-异天冬氨酸甲基转移酶基因,2个基因通过协同作用提高水稻抗逆性,增强水稻种子的耐贮性[59];Lee等[60]报道了转铜锌SOD基因的烟草能够提高老化后种子的发芽率,证实抗氧化酶在维持种子活力过程中有重要意义;Narayana等[61]对老化后的水稻种子活力与幼苗活力进行了定量分析,发现在加速老化后的种子内部会累积糖基化和脂质过氧化化合物,如:丙二醛,甲基乙二醛,Maillard和Amadori的产物,这些代谢物质的积累导致细胞膜完整性丧失,降低能量代谢,损害RNA与蛋白质的合成,从而降低种子的活力。醛类物质也是植物生长发育代谢物质,水稻基因OsALDH7可以将丙二醛等醛类物质氧化分解为相应的羧酸,构建该基因的突变体后发现,突变体植株体内赖氨酸的降解速度变快,显著降低了种子的耐贮性[62]。金属硫蛋白(MTs)主要维持植物体内金属稳态与活性氧清除,Zhou等[63]以拟南芥为背景材料,过量表达NnMT2a和NnMT3后发现转基因植株ROS清除能力变强,得到的种子抗老化能力提高,种子活力显著的增强;Seo等[64]报道了一个与磷脂相关的基因AtDLAH,该基因在幼苗中有较高的表达量,编码的蛋白质只存在于线粒体,过量表达该基因可以降低拟南芥种子的脂质过氧化水平,提高老化后的种子活力;
种子的休眠与耐贮性呈正相关,种子的休眠性越强,表现出的耐贮性就越强。一些与激素合成相关基因的克隆进一步揭示了种子耐贮的调控机制。ABA调控种子成熟、脱水耐性与诱导种子的休眠。ABA羟基化是植物体内ABA分解的主要途径,植物体内有3种不同的ABA羟基化途径(C-7′,C-8′,C-9′),其中C-8′被认为是ABA分解代谢的主要途径[65]。ABA 8′-羟化酶是一种与膜结合的细胞色素P450(CYP450)单加氧酶,Kushiro等[66]证实cyp707a2突变体的拟南芥种子表现出高度的休眠,内源ABA的积累是野生型的6倍,能显著提高种子的耐贮性。OsDET1是拟南芥DET1的直系同源基因,Zang等[67]以水稻为背景材料,OsDET1的RNAi植株ABA生物合成受到明显抑制,而在过表达OsEDT1的植株中内源ABA的含量明显提高,种子的休眠性得到明显的增强,也提高了种子的耐贮性。DOG1调控拟南芥种子的休眠,促进种子内源ABA的积累,抑制种子的萌发;Bentsink等[68]以拟南芥为背景材料,通过构建了DOG1敲除突变体,发现该突变体种子的耐贮性显著降低。AtPER1是种子内部所特有的过氧化酶,与种子的休眠存在一定的关系,Chen等[69]发现atper1-1与atper1-2突变体的种子初级休眠受到抑制,并且伴随着内源激素ABA的降低以及GA含量的增加。Quettier等[70]为了了解蛋白酪氨酸磷酸酶(PTP)在ABA信号传导中的作用,在拟南芥获取纯合的T-DNA插入系,证实了编码双特异性PTP的PHS1对ABA的信号传导起负调控的作用。Bueso等[71]认为拟南芥ATHB25(Homeobox25/Zinc finger protein domain)正调控GA3ox2,促进与GA相关物质的合成以及种皮细胞细胞壁的形成,且能显著增强种子的耐贮性。DOF1.5与DOF2.3是拟南芥DOF类转录因子,在成熟的种子中特异表达,具有重要的调控作用,Bueso等[72]发现DOF1.5与DOF2.3可增强种子耐贮性。生长素代谢相关基因也参与种子成熟发育的过程,调控种子的耐贮性。Liu等[73]发现,ABI4通过介导GA与ABA的平衡会对种子的初级休眠存在一定的调控作用,并且负向调控子叶变绿。ABI3作为ABA的下游合成基因,是抑制种子发芽、调节种子休眠的关键基因,同时证实生长素也能通过激活ABI3加深种子的休眠,从而增强种子耐贮性。
4 展望
种子是植物的遗传物质载体,也是农业生产的基本物质资料和主要产出形式。收获后的种子由于贮藏环境的影响,不可避免地会发生老化劣变,并且对种子的发芽活力造成很大的影响。由于农时与政策的需要,每年都必须有一定数量的种子储备与贮藏,因此获取强耐贮性的水稻品种更为迫切。当前对水稻耐贮性的研究已有一定的研究报道,但挑战与机遇并存,培育优良耐贮性的水稻品种仍迫在眉睫,具体的问题与笔者的应对建议做如下总结。
(1)加强基础领域研究,前人主要是通过利用栽培稻品种构建人工群体进行种子耐贮性QTL研究,也检测到了大量控制水稻种子耐贮性QTL,甚至对部分主效QTL进行了精细定位和图位克隆,然而传统的QTL定位效率不高,周期较长,且人工群体遗传变异较少,因此在一定程度上阻碍了种子耐贮性的研究。笔者认为可以从3个方面进行改进:第一,着重利用自然群体发掘种子耐贮性位点,在更广泛的遗传背景下更有利于开展种子耐贮性研究;第二,着重从野生稻种质资源中发掘新基因、新位点,栽培稻经过人工驯化后,丢失了许多重要的基因,利用野生稻资源可以找到丢失的种子耐贮性位点。第三,更新技术手段,可以从转录组、蛋白组、代谢组水平上对水稻耐贮机理进行探讨。
(2)加快分子标记辅助选择育种的进程,在当前的分子辅助选择育种过程中,育种家们的育种目标一直停留在高产、抗虫、抗病、优质等方面,忽视了种子活力相关性状。需知优良品种推广的前提是具有优良的种子,因此加强种子相关性状的分子标记辅助选择尤为重要,特别是加强水稻种子耐贮性的分子标记辅助选择育种,培育耐贮性较强的品种。不仅能有效的保障用种安全,而且可以有效地降低中国粮食在贮藏过程中的损失,为中国的粮食安全提供保障。
(3)育种实践证明:耐贮性较强的品种也同时具有较强的休眠特性。因此在育种过程当中休眠性强的品系,不可避免地被育种家们淘汰掉,也就间接淘汰了耐贮性强的品系。笔者研究发现水稻种子的休眠性和耐贮性呈正相关,但同时也发现不少休眠性弱而耐贮性强的种质材料,因此可以在基数庞大的水稻种质群体材料中遴选出休眠性适中或较弱但具有耐贮性强的种质资源,并利用这些材料作为供体材料,进而选育耐贮性强的水稻新品种。
