核因子E2相关因子2在椎间盘退变中的作用△
2022-02-24陈海伟刘明强张广智康学文
陈海伟,刘明强,张广智,康学文
(兰州大学第二医院骨科,甘肃 兰州 730030)
椎间盘退变(intervertebral disc degeneration,IDD)是一种临床常见的肌肉骨骼系统退行性疾病,大约70%~85%的人一生中至少会经历一次腰痛,严重降低了生活质量[1]。然而,IDD的具体发病机制尚不完全清楚。目前认为,多种因素参与IDD的发生发展,例如:炎症反应、氧化应激、线粒体功能障碍、机械负荷异常、营养剥夺和遗传因素等[2~4]。核因子E2相关因子2(nuclear factor E2 related factor 2,Nrf2)是调控细胞氧化应激反应的重要转录因子,同时也是维持细胞内稳态的中枢调节者。Nrf2是调控IDD细胞内稳态的关键调节因子,有可能作为治疗IDD的潜在靶点。本文就这方面研究进展作一综述,为后续相关研究提供参考。
1 Nrf2的结构与功能
Nrf2是亮氨酸拉链(basic leucine zipper,bZIP)转录因子,由7个Nrf2-ECH同源结构域(Neh1–7)的模块蛋白组成,每个域具有不同的功能[5]。Neh1结构域的氨基酸序列是高度保守的基本区域,可介导Nrf2与抗氧化反应元件(antioxidant response elements,ARE)序列的结合;Neh2结构域介导与Nrf2负调控因子Kelch-like ECH-associated protein 1(Keap1)和7个赖氨酸残基的相互作用,用于Nrf2的泛素化和随后的蛋白酶体降解;C端Neh3结构域具有反转录活性,并与Neh4和Neh5结构域协同激活Nrf2靶基因的转录;Neh6域是一个富含丝氨酸的区域,参与独立于Keap1的Nrf2稳定性的负调节;Neh7结构域抑制Nrf2的转录活性[6]。
Nrf2可通过协同诱导ARE驱动的细胞保护性基因转录来抵消氧化应激、炎症和线粒体功能障碍[7];其中ARE的基因包括NAD(P)H醌氧化还原酶(NQO1),血红素加氧酶-1(HO-1),超氧化物歧化酶1(SOD-1),过氧化氢酶1,谷胱甘肽过氧化物酶,谷胱甘肽还原酶,过氧化物酶等等。Nrf2的功能通过增加ARE下游基因表达来体现[8]。因此,Nrf2及其下游基因对于细胞稳态的维持至关重要。
2 椎间盘的结构及Nrf2的表达
椎间盘(intervertebral disc,IVD)是一个复杂的结构,由周围部的纤维环(anulus fibrosus,AF)、中间部的髓核(nucleus pulposus,NP)和连接上下椎体的软骨终板(cartilage endplate,CEP)三部分构成[9,10]。AF由一系列同心片层组成,主要由相互交替I型胶原纤维束构成,提供抗拉强度;NP主要由富含II型胶原蛋白、弹性蛋白和蛋白聚糖的细胞外基质(extracellular matrix,ECM)组成,其中蛋白聚糖是高负离子的物质,提供渗透特性,使NP在压缩载荷下保持高度和膨胀;CEP在结构上将椎间盘固定在相邻的椎骨上,其细胞外基质主要富含II型胶原蛋白和蛋白多糖[11]。椎间盘退变的特点是细胞外基质破坏增加,基质合成异常,椎间盘高度丧失,导致神经根受到压迫,产生下腰痛。最近研究表明,IDD患者NP组织中Nrf2的表达随着NP变性加重而逐渐降低,且Nrf2的缺乏会加剧NP细胞衰老[12]。因此,Nrf2的缺乏可能参与了IDD的发生发展。
3 Nrf2的表达受到非编码RNA调控
最近研究表明,非编码RNA(noncoding RNAs,ncRNAs)在转录后调节基因表达,并在IDD过程中起关键作用[13]。已经证实多种miRNA参与细胞凋亡,细胞外基质的降解,炎症反应及软骨终板的退变[14]。最近研究表明,lncRNA能够通过调节Nrf2的表达,参与IDD的进程。Li等[15]用实时定量PCR分析临床组织标本,结果显示LncRNA NEAT1表达水平随IDD等级的增加而增加;在人NP细胞中,转染lncRNA NEAT1使其过表达,可导致Nrf2的表达降低。同样,Kang等[16]用实时定量PCR检测ln⁃cRNA ANPODRT的表达,结果显示ANPODRT的表达随着IDD等级的增加而减少;在NP细胞中,转染ANPODRT使其过表达,可以破坏Keap1与Nrf2缔合,从而抑制其下游靶基因的激活。这些结果表明,表观遗传学中的ncRNAs可能调控Nrf2及其下游分子的表达,减缓IDD的发展 (图1)。
4 Nrf2在IDD的作用机制
4.1 Nrf2在IDD中的抗氧化作用
氧化应激可以定义为促氧化剂和抗氧化剂之间的不平衡,导致过量的活性氧(ROS)对细胞的损伤。ROS包括超过氧化阴离子(O2-),羟基自由基(-OH)和过氧自由基(ROO-),是正常细胞内代谢产生的一种信号分子,可促进细胞生存和组织更新或抑制细胞生存基因的表达[17,18]。研究表明,随着IDD的进展,IVD中的ROS产生增加[19]。过量ROS破坏细胞内氧化还原稳态,可使NP细胞衰老和凋亡加速,且形成炎症微环境,最终导致ECM降解增加。Nrf2在抑制氧化应激中起关键作用,能够激活下游基因的转录,包括NAD(P)H醌氧化还原酶1(NQO1),血红素加氧酶1(HO-1)和超氧化物歧化酶(superoxide dismutase,SOD)等[20]。TBHP 和 H2O2是氧化应激反应的诱导剂。Tang等[12]研究发现,Nrf2的表达与人体椎间盘退变呈负相关,H2O2诱导大鼠NP细胞发生退变,导致Nrf2的表达降低。Ke⁃ap1基因沉默会抑制Keap1与Nrf2的解离,导致p62的表达增加,而其靶基因HO-1和NQO1的表达减少。Bai等[21]用免疫组化显示严重IDD的患者NP存在较高水平的ROS,其研究证实Nrf2的过表达能减少ROS的水平,减轻氧化应激的损伤。这些结果表明Nrf2基因可以调控NP细胞的退变过程。
最近的研究表明,多种化合物单体可以激活Nrf2基因的表达,保护NP细胞免受氧化应激的损伤,从而延缓IDD的进展。金线莲苷是从乌参中提取出来的一种活性化合物,具有肝保护和抗骨质疏松症等功能[22]。Wang 等[23]研究报道,金线莲苷增加Nrf2的表达,减轻由TBHP诱导NP细胞中线粒体ROS的水平,而Nrf2敲除降低了金线莲苷清除ROS的能力。金雀异黄素是豆科植物的天然类黄酮,具有抑制炎症,调节细胞凋亡的作用[24]。在氧化应激下,Wang等[25]报道了金雀异黄素可降低H2O2诱导NP细胞中ROS的水平,增加Nrf2以及下游分子HO-1和NQO1的表达;而Nrf2敲除会减弱金雀异黄素的抗氧化作用。茶多酚是绿茶中的主要多酚类化合物,对多种病理疾病(包括癌症、糖尿病和心血管疾病)具有有益作用[26]。Song 等[27]研究表明,茶多酚增加Nrf2及其下游NOX4和SOD2的表达,减少ROS的产生,其抗氧化的机制是通过Keap1/Nrf2/ARE信号通路发挥作用。淫羊藿苷是淫羊藿中含量最丰富的类黄酮成分,能够调控端粒和端粒酶的活性,延缓细胞的衰老[28]。Wang 等[29]发现淫羊藿苷可以减轻体内大鼠椎间盘退变,促进退变NP细胞增殖,并抑制NP细胞中炎性因子IL-1β、IL-6表达。另外,Hua等[30]通过DCFH-DA探针检测ROS的含量,H2O2刺激导致ROS的产生增加;而用淫羊藿苷预处理可以增加NP细胞中Nrf2的表达,并且减少ROS的产生。同样,萝卜硫烷是一种来自于葡甘露聚糖的异硫氰酸盐,能够靶向激活Nrf2,减少ROS的产生[31]。这些研究结果表明Nrf2是调控ROS水平的关键调控因子。
线粒体是产生ROS的主要场所,线粒体功能障碍或线粒体损伤均会导致ROS的水平异常,对细胞产生氧化损伤。其中线粒体靶向抗氧化剂Mitoqui⁃none(MitoQ)可防止线粒体氧化损伤[32]。Kang等[33]研究发现MitoQ可降低压缩诱导下的人NP细胞中线粒体ROS的产生和Keap1蛋白表达,而增加Nrf2蛋白表达;Nrf2沉默部分消除了MitoQ对抗氧化应激对细胞的损伤作用。线粒体的抗氧化剂(Mito-TEMPO)可清除细胞内的ROS。Kang等[34]研究发现Mito-TEMPO可降低H2O2诱导终板软骨细胞中线粒体ROS的升高,增加Nrf2蛋白及其靶标SOD-2、HO-1和NQO-1的表达;而Nrf2敲除导致细胞内ROS的水平增加。这些结果表明Nrf2参与线粒体功能的维持,从而减少氧化应激的损害。此外,研究表明在软骨终板细胞中,细胞自噬会影响Nrf2的表达和核易位,从而控制抗氧化蛋白的表达,保护软骨终板免受氧化应激引起的变性[35](表1,图1)。
4.2 Nrf2在IDD中的抗炎作用
IVD细胞或巨噬细胞所分泌的促炎性细胞因子的升高与IDD的进展有关;在退化级联反应中,内部AF和NP的血管形成,使巨噬细胞迁移到IVD中,导致炎症级联反应的放大,从而加速了IDD的进程[36]。分泌的炎性介质,特别是 TNF-α、IL-1α/β、IL-6、IL-8、IL-2、IL-17、IL-10、IL-4、IFN-γ 和PGE 2,促进了细胞外基质降解、衰老和凋亡[37]。最近研究表明,Nrf2通过协调炎症细胞的募集和通过ARE调节基因表达来促进抗炎过程[38]。因此,Nrf2信号通路可能是调控炎症的重要途径。
植物来源的单体化合物,例如汉黄芩素、槲皮素、金合欢素、桑辛素及姜黄素等的抗炎作用,是通过激活Nrf2基因的表达,从而保护NP细胞免受炎症的损伤。Fang等[39]报道汉黄芩素激活大鼠NP细胞中Nrf2基因,以及SOD-2、NQO-1和HO-1基因的表达,抑制炎性介质的表达;而Nrf2的沉默进一步证实炎症介质的表达由Nrf2介导。Shao等[40]研究表明,槲皮素促进Nrf2及其下游效应蛋白HO-1的表达,抑制了IL-1β介导的NP细胞中IL-6和IL-8水平的升高。Wang等[41]发现金合欢素增加核内Nrf2蛋白以及下游HO-1、NQO1和SOD的表达,抑制THBP诱导的炎症因子iNOS和COX-2的表达。Gu等[42]用ELISA检测炎症因子的表达,结果显示LPS刺激显著增加了IL-1β、IL-6和TNF-α的水平,Nrf2和HO-1水平下调;而桑辛素处理显著增加了Nrf2和SOD的表达,减轻了炎症因子的表达;而当Nrf2基因被沉默时,桑辛素的抗炎作用被大大逆转。Cherif等[43]用免疫组化显示退化的椎间盘组织中Nrf2的表达减少;而经姜黄素预处理后,Nrf2的表达增加,ELISA显示IL-6、IL-8的表达减少。小豆蔻素通过激活Nrf2/HO-1信号轴来抑制NP细胞中IL-1β诱导的炎性因子的表达[44]。这些结果表明,Nrf2以及其下游的分子参与NP细胞炎症反应的调节。因此,Nrf2可能是参与调控NP细胞炎症反应的关键调控因子 (表1,图1)。
4.3 Nrf2改善IDD中的线粒体功能障碍
线粒体是IVD细胞中能量供应的主要来源,其通过氧化磷酸化过程产生三磷酸腺苷(Adenosine tri⁃phosphate,ATP)从而保证细胞的正常功能活动,并且也是ROS产生的贡献者[45]。线粒体自噬会清除过多的ROS以维持线粒体稳态;ROS水平过高导致线粒体功能障碍,触发程序性细胞死亡,加速IDD的进展[37-46]。Drp1从细胞质迁移到线粒体是线粒体裂变的先决条件。Kang等[33]研究发现压缩增强Drp1向线粒体的易位;用线粒体裂变阻滞剂Mdivi-1处理NP细胞,Mdivi-1显著抑制了Drp1向线粒体的易位。因此,线粒体结构的完整对于线粒体稳态和NP细胞存活至关重要。
研究证实,植物来源的单体化合物可以激活Nrf2基因,参与维持线粒体的结构与功能。线粒体中的促凋亡蛋白Cytochrome C(Cyto C)释放到细胞质中,能够促进细胞的凋亡。金线莲苷减少TBHP诱导的线粒体膜电位(mitochondrial membrane potential,MMP)的丢失,并抑制了Cyto C的释放;而Nrf2的沉默减弱金线莲苷对线粒体的保护作用[23]。Mito⁃Tracker Green是一种绿色荧光染料,能够显示活细胞中线粒体膜电位的高低。Wang等[47]用MitoTracker Green染色显示白藜芦醇可以减轻TNF-α诱导的NP细胞荧光强度的损失和ATP生成减少,并增加核Nrf2和HO-1的水平;Nrf2敲低显示更弱的Mito⁃Tracker荧光强度,提示Nrf2在线粒体内稳态中起保护作用。Hua等[30]用JC-1染色显示线粒体的跨膜电位(ΔΨm)的强度,淫羊藿苷可以增加NP细胞中Nrf2的表达,并且降低H2O2刺激NP细胞中ΔΨm的产生,维持线粒体膜电位的稳定。
除上述研究之外,线粒体的抗氧化剂(Mito-TEMPO) 通过增加Nrf2的表达降低H2O2处理引起终板软骨细胞的ΔΨm产生,并且增加ATP的生成,减轻线粒体功能障碍[34]。这些结果表明,Nrf2可能通过改善线粒体的结构与功能进而延缓IDD的进展(表1,图1)。

图1 Nrf2作用IDD可能的机制图 多种不利因素刺激导致IVD细胞发生炎症、氧化应激和线粒体功能障碍,加速IDD的进展。Nrf2的激活剂和非编码RNA能够调控Nrf2基因及其下游分子(NQO1、SOD、HO-1和NOX4)的表达、调节自噬,发挥抗氧化、抗炎、改善线粒体功能障碍,从而延缓IDD的发展
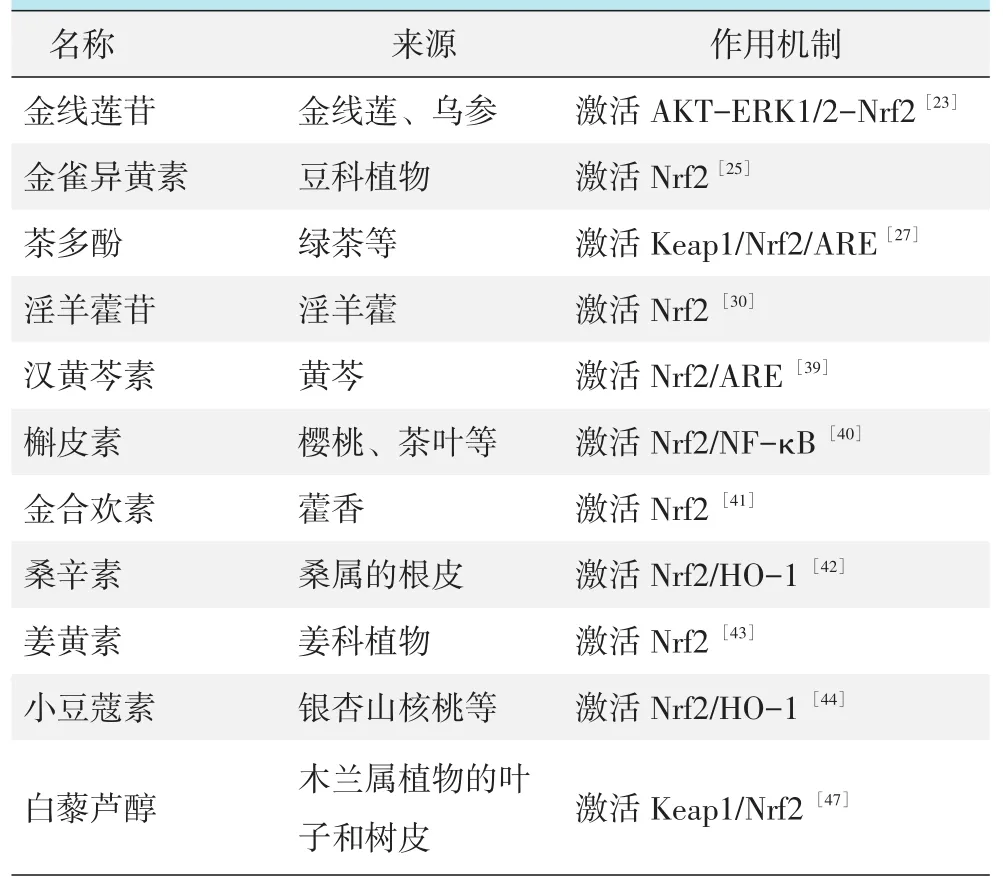
表1 Nrf2的激活剂
5 小结
Nrf2的表达与IVD的变性程度呈负相关,并且Nrf2的表达还受到表观遗传学的调控,因此,Nrf2基因与IDD的进展密切相关。Nrf2除有上述抗氧化、抗炎、改善线粒体功能障碍的作用外,Nrf2还可通过诱导自噬蛋白p62和其他自噬相关基因的表达来促进自噬,调控氧化应激对髓核细胞的损害;很多从天然产物提取的化合物如金线莲苷、金雀异黄素、茶多酚、淫羊藿苷、汉黄芩素、槲皮素和白藜芦醇等可有效激活Nrf2,减轻氧化应激、炎症对IVD细胞的损伤,并维持线粒体的结构与功能。另外,激活Nrf2对骨关节炎、慢性肾病、糖尿病等也有一定的改善作用。目前,有关Nrf2的研究大多是通过细胞和动物实验进行的,缺乏相应的临床依据。随着对Nrf2基因不断地深入研究,有望研制出靶向Nrf2的临床药物,为IDD的预防和治疗提供新的方法和思路。
