甲基丙烯酸环氧丙酯诱导16HBE细胞恶性转化过程中LncRNA表达特征分析及ceRNA调控网络预测
2022-02-15李昕苇王全凯马顺鹏乌瀚宝栎尔顾轶婷康同影许建宁
李昕苇,王全凯,马顺鹏,王 苗,乌瀚宝栎尔,顾轶婷,康同影,许建宁,*
(1.中国疾病预防控制中心职业卫生与中毒控制所,北京 100050;2.中国疾病预防控制中心化学污染与健康安全重点实验室,北京 100050)
甲基丙烯酸环氧丙酯(glycidyl methacrylate,GMA),又称甲基丙烯酸缩水甘油酯,主要在丙烯酸粉末涂料、乳胶涂料、胶粘剂的合成和加工中被广泛使用。目前国际癌症组织(International Agency for Research on Cancer,IARC)已完成对GMA的致癌性评估,根据“强有力”的致癌机制证据及“充分”的实验证据,将GMA归类为“对人类很可能致癌”(Group 2A)[1]。本课题组既往研究发现,在GMA诱导人支气管上皮细胞恶性转化过程中,细胞基因组和表观遗传学发生异常改变,过程中涉及众多细胞信号转导通路,并可能导致下游分子蛋白的一系列改变[2-4]。
长链非编码RNA(long non-coding RNA,LncRNA)是一种具有200多个核苷酸、未翻译成蛋白质的转录本,通常认为它是基因转录和染色质结构的限制调节因子,然而,随着研究的深入,LncRNA逐渐受到覆舟关注,亦发现它们具有翻译调节、信号转导等多种功能[5]。竞争性内源RNA(competing endogenous RNA,ceRNA)是LncRNA通过结合微小RNA(microRNA,miRNA)应答元件竞争性吸附miRNA,抑制miRNA与信使RNA(message RNA,mRNA)结合,从而导致靶基因沉默[6],其中,LncRNA与mRNA之间存在正调控关系,与miRNA存在负调控关系[7]。因此本次试验对GMA诱导细胞恶性转化不同时期差异表达的LncRNA进行检测分析,并以此为依据构建ceRNA调控网络,对差异表达的LncRNA功能进行预测和分析。
1 材料与方法
1.1 主要试剂与仪器
GMA(纯度≥98.5%)和二甲基亚砜(DMSO)购于Sigma公司(美国);胰蛋白酶、EDTA-Na2购于Amresco公司(美国);MEM培养基粉购于Hyclone公司(美国);标准胎牛血清购于Gibco公司(美国);100×的青、链霉素混合液购于北京索莱宝科技有限公司;TRIzol试剂购于上海碧云天生物技术有限公司,2×PCR master mix购于Arraystar公司(美国)。
本研究使用的主要仪器有:CO2培养箱(Thermo公司,美国);台式高速低温离心机(BECKMAN公司,美国);倒置显微镜(Olympus公司,日本);紫外可见分光光度计(UNICO公司,美国);7900HT Fast Real-Time PCR仪(Biosystems公司,美国);ND-1000分光光度计(NanoDrop公司,美国);ViiA 7实时定量PCR反应体系(Applied Biosystems公司,美国)。Arraystar Human LncRNA Microarray V4.0芯片技术服务和引物设计及ceRNA调控网络构建的技术支持由上海康成生物公司提供。
1.2 细胞培养
人支气管上皮细胞(human bronchial epithelial cells,16HBE)来自美国加利福尼亚大学,将冻存于液氮中的细胞复苏后,用质量浓度为0.25%的胰蛋白酶消化后,用10%标准胎牛血清(FBS)的MEM培养液,于37℃、CO2体积分数为5%的恒温培养箱中培养过夜。基于课题组既往关于GMA的细胞毒性和克隆形成率的研究结论,选择浓度为8μg/mL的GMA重复染毒诱导16HBE细胞发生恶性转化[8]。取对数生长期的细胞进行接种,24 h后向培养液中加入浓度为8μg/mL的GMA进行染毒(DMSO作为对照)。每次染毒时间72 h,间隔24 h后进行2次染毒,重复染毒3次后结束染毒,记为第1代;MEM培养液调整为5%FBS继续培养,每隔5 d传代1次。课题组既往研究[9]表明,传代培养至第30代,GMA染毒组的16HBE细胞具备恶性转化细胞的生物学特性。根据课题组既往ConA凝集试验及锚着非依赖性试验的研究结果,选择收取第10、20和30代的GMA染毒组细胞和DMSO对照组细胞,记为转化前(P10)、中(P20)和后期(P30)。
1.3 LncRNA芯片分析
收集GMA处理组和DMSO对照组的P10、P20和P30细胞,TRIzol提取总RNA,使用NanoDrop-1000分光光度计检测RNA浓度和纯度,于-80℃冰箱中保存备用。使用Arraystar RNA Flash标记试剂盒标记总RNA样品,再使用Agilent SureHyb进行杂交实验,完成杂交后洗涤芯片,采用Agilent DNA Microarray Scanner扫描芯片的信号值,标准化处理后得到不同时期细胞LncRNA表达谱。
最后,通过人工检索GMA重复染毒诱导16HBE细胞恶性转化过程中P10、P20和P30代细胞LncRNA表达谱的差异性表达结果,以3个时期差异倍数的对数绝对值|log2FC|≥2.0且P<0.05为标准筛选得到在不同时期表达水平均上调的LncRNA,确定GMA诱导16HBE细胞恶性转化过程及其不同时期涉及的相关LncRNA。
1.4 ceRNA调控网络构建
ceRNA调控网络根据miRNA建立调控关系,因此以表达水平具有上调趋势的LncRNA为中心,通过TargetScan和miRanda数据库进行差异性表达LncRNA靶基因的预测[10-11],根据超几何检验来验证LncRNA与mRNA之间是否存在ceRNA关系及其显著性差异,公式如下:

经过筛选后得到P<0.05的ceRNA对用于构建ceRNA调控网络。在此基础上,利用网络工具Cytoscape构建GMA诱导16HBE细胞恶性转化的ceRNA调控网络。
1.5 实时荧光定量PCR验证LncRNA G013234、CTA-384D8.35、G087116的表达水平
使用逆转录试剂盒(Invitrogen)将已提取的P10、P20和P30代细胞的总RNA样品反转录为cDNA,再以cDNA为模板进行实时荧光定量PCR(quantitative realtime PCR,qPCR)反应,引物序列见表1。qPCR体系(10μL):2×Master Mix 5μL、上下游引物各0.5 μL、cDNA模板2μL及无核酸酶水2μL;扩增条件为:95℃,预变性10 min;95℃,变性10 s;60℃,退火60 s,40个PCR循环。以管家基因βactin为内参,同期DMSO对照组基因的平均值为基准,以GMA组基因表达量除以样品内参基因表达量,得到的比值即所验证的LncRNA的相对表达量。
1.6 统计学分析
采用SPSS 25.0软件进行统计分析,LncRNA相对表达量的比较采用单因素方差分析,组间两两比较采用SNK检验,检验水准α=0.05。
2 结果
2.1 GMA诱导16HBE细胞恶性转化过程中LncRNA芯片的筛选结果
对GMA重复染毒诱导16HBE细胞恶性转化过程中P10、P20和P30代细胞LncRNA芯片结果进行分析,筛选得到在不同时期表达水平均上调的LncRNA共16个,见图1,差异性表达LncRNA变化情况如表2所示。其中恶性转化的P20细胞中AP000619.5、CTA-384D8.35及G071314表现尤为明显,与同代龄DMSO溶剂对照组相比上调超过10倍。

图1 GMA诱导16HBE细胞恶性转化过程中表达水平一致性上调的LncRNA

表1 qPCR测定的引物序列

表2 第10、20、30代染毒组细胞中表达一致性上调的LncRNA
2.2 ceRNA调控网络的构建和功能富集分析
通过TargetScan和miRanda数据库预测GMA诱导16HBE细胞恶性转化过程中差异表达的16个LncRNA的目标靶向mRNA和miRNA,二者结果取交集后,进行超几何分析并计算ceRNA对之间表达量的Pearson相关系数>0.9,最终筛选出3个关键的差异表达LncRNA,32个mRNA和204个miRNA,并以此为基础构建GMA诱导16HBE细胞恶性转化过程相关的ceRNA调控网络,见图2。构建的ceRNA调控网络结合hub基因评分,发现LncRNA G013234、CTA-384D8.35、G087116可能通过LncRNA G013234-hsamiR-378b-TMEM129、LncRNA CTA-384D8.35-hsamiR-486-3p-LYPD、LncRNA G087116-miR-29b-3p-NAV1调控轴在GMA诱导16HBE细胞恶性转化过程中发挥作用。
2.3 LncRNA G013234、CTA-384D8.35、G087116相对表达量的检测结果

绿色圆形:LncRNA;蓝色圆形:mRNA;红色圆形:miRNA.
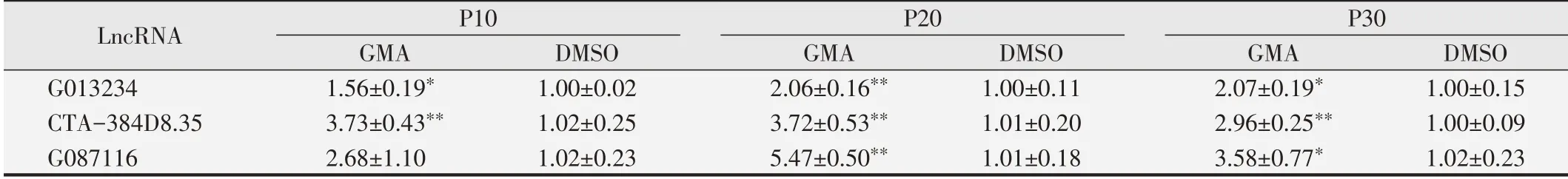
表3 qPCR检测不同时期16HBE细胞LncRNA相对表达水平
对构建ceRNA调控网络的关键差异表达LncRNA进行qPCR检测,结果见表3。与同代龄DMSO对照组相比,GMA组细胞LncRNA G013234、CTA-384D8.35和G087116相对表达量在P10、P20和P30代细胞中均上调。除G087116在P10代细胞中表达量差异无统计学意义(P>0.05)外,其余LncRNA的相对表达量在两组间差异均具有统计学意义(P<0.05)。qPCR检测结果显示细胞不同时期LncRNA相对表达水平的变化趋势与芯片检测结果一致。
3 讨论
LncRNA是一类进化保守的非编码RNA,参与细胞表观遗传调控、周期调控、分化调控等过程,同时LncRNA可能包含未被发现的细胞恶性转化驱动因子。LncRNA可以通过RNA-蛋白质,RNA-RNA和RNA-脂质相互作用,在细胞恶性转化过程中发挥关键的信号转导介质作用[12]。本研究主要研究在GMA诱导16HBE细胞发生恶性转化过程中相关差异表达LncRNA的筛选及表达改变,并以此为基础构建ceRNA调控网络,探讨LncRNA在16HBE细胞恶性转化过程中可能存在的生物学作用。
本研究采用GMA重复染毒诱导16HBE细胞恶性转化,根据P10、P20和P30代细胞的芯片检测结果,筛选在细胞恶性转化不同时期差异表达的LncRNA,共发现16个LncRNA在不同时期表达均发生上调,以差异性表达上调的LncRNA为基础构建ceRNA调控网络,构建的ceRNA调控网络包含3个关键的差异表达LncRNA即G013234、CTA-384D8.35及G087116,32个mRNA和204个miRNA。在PubMed、万方、中国知网等文献数据库中检索上述关键差异表达LncRNA的相关文献,并在NCBI、Ensemble等数据库中查询这些LncRNA相关注释信息,发现LncRNA G013234、CTA-384D8.35及G087116可以检索到参与肿瘤发生发展的研究及相关文献。后经qPCR检测ceRNA调控网络中LncRNA在细胞恶性转化不同时期的相对表达量,检测结果与芯片结果一致,进一步说明上述LncRNA可能在GMA诱导16HBE细胞恶性转化过程中发挥重要作用。
CeRNA调控网络的构建为LncRNA和mRNA之间更复杂精细的调控提供了假说,也为研究LncRNA在细胞内的功能及与其他编码、非编码RNA的相互作用提供了新的思路[13]。在本研究构建的ceRNA调控网络中,LncRNA G013234-hsa-miR-378b-TMEM129,LncRNA CTA-384D8.35-hsa-miR-486-3p-LYPD,LncRNA G087116-miR-29b-3p-NAV1这3个调控轴是与GMA诱导16HBE细胞恶性转化过程关联较为密切的ceRNA三元组(P<0.05)。Li等[14]发现,胰腺导管腺癌(pancreatic ductal adenocarcinoma,PDAC)组织中CTA-384D8.35表达与IL-6呈正相关,同时CTA-384D8.35直接结合IL-6启动子,激活IL-6转录,并上调IL-6表达和分泌以激活PDAC细胞中STAT3信号通路;且胰腺导管腺癌中PanIN-3病变部位miRNA hsa-miR-486-3p发生异常过表达[15]。目前有研究表明,hsa-miR-486-3p在类风湿性关节炎患者的血液中异常表达且富集到靶基因产物STAT3[16]。而Xu等[17]发现LYPD可通过抑制IL-6和TNF-α的分泌来调控结直肠癌细胞的增殖和迁移,并且TNF-α和IL-6的分泌与STAT3和P65的磷酸化呈正相关。因此,LncRNA CTA-384D8.35可能通过与hsa-miR-486-3p的相互作用调节细胞因子IL-6的表达,进而影响STAT3信号通路的传导,导致LYPD在GMA诱导16HBE恶性转化细胞中的分泌改变,从而影响细胞的增殖和迁移等活动。
有研究表明has-miR-378在胆管癌中可作为癌基因发挥作用,促进胆管癌细胞增殖、迁移和侵袭[18],且has-miR-378参与骨肉瘤的发生及其肺转移的生物学过程[19]。TMEM129是非糖基化蛋白,包含一个非剪切信号锚定序列,可用于催化分泌蛋白细胞质降解所需的泛素化反应[20];而TMEM129所涉及的DNA甲基化过程也与黄种人和白种人的早期肺腺癌的预后有关[21]。上述研究表明,LncRNA G01323可能调节TMEM129所涉及的DNA甲基化过程,从而激活原癌基因has-miR-378b,在GMA诱导16HBE细胞发生恶性转化。
Zhang等[22]的研究表明,miR-29b-3p通过下调TRAF3促进MDA-MB-231三阴性乳腺癌细胞的进展。NAV1是一种主要在神经元中发现的蛋白,为电压门控钠离子通道,据报道在微管发育中发挥关键作用[23]。亦有文献报道其在乳腺癌、前列腺癌等肿瘤细胞中出现异常表达,如Luo等[24]研究显示,Nav1以电压门控钠通道的形式在转移性乳腺癌细胞中异常激活和表达,从而增强乳腺癌细胞在体内外的转移和侵袭性。Papadopoulos等[25]的研究表明,在单侧输尿管梗阻的体外试验中,NAV1遵循经典的调控模式,即miR-29b-3p的上调导致靶标NAV1的下调,证实NAV1可能是miR-29b-3p的直接靶点;同时有报道指出,根据时间和条件的改变,miRNA可能与诱导其靶标mRNA的调控方向相同,这亦为miR-29b-3p调控NAV1可能的间接机制提供了解释[26]。因此,LncRNA G087116可能通过调节miR-29b-3p的表达导致NAV1的表达改变,从而参与GMA诱导16HBE细胞恶性转化过程。
本研究通过LncRNA芯片分析和qPCR验证发现,在GMA诱导16HBE细胞恶性转化不同时期细胞中LncRNA G013234、CTA-384D8.35、G087116发生表达上调。以上述差异表达LncRNA为基础构建的ceRNA调控网络及预测得到GMA诱导细胞恶性转化相关的关键mRNA,筛选出LncRNA G013234-hsa-miR-378b-TMEM129、LncRNA CTA-384D8.35-hsa-miR-486-3p-LYPD、LncRNA G087116-miR-29b-3p-NAV1调控轴,并且这些调控轴可能在GMA诱导16HBE细胞恶性转化的过程中发挥重要作用。
