稻虾鳝立体综合种养水体微生物多样性分析
2022-02-13张圆圆赵良杰朱文锦
张圆圆,赵良杰,李 泓,朱文锦
(1. 上海海洋大学 农业农村部鱼类营养与环境生态研究中心,上海 201306;2. 河南省水产科学研究院,河南 郑州 450044;3. 信阳农林学院 水产学院,河南 信阳 464000)
黄鳝(Monopterus albus)又名鳝鱼,隶属辐鳍亚纲(Actinopterygii)、合鳃鱼目(Syn-branchiformes)、合鳃鱼科(Synbranchidae)、黄鳝属(Monopterus),是我国淡水名优养殖品种。黄鳝因肉质细嫩、营养丰富及独特的药用价值越来越受消费者青睐,但受人工捕捞和水域污染等影响,野生黄鳝资源日益减少,近年来,人工网箱养殖黄鳝因占用水面少、养殖效益高等特点已成为黄鳝养殖的主要模式[1]。但常规网箱养殖黄鳝存在网箱底部容易积累大量残饵粪便、水体流动性差、水质易缺氧老化、氨氮和亚硝酸盐含量过高的情况,成为制约黄鳝网箱养殖的关键技术问题[2]。
养殖水体中微生物主要来自空气、土壤、动植物残体及分泌排泄物等,广泛参与水体中氨化、固氮、硝化及反硝化等物质循环[3],并能够反映特定系统的水质状况,是水体的重要组成部分,在养殖水环境中具有重要的意义[4]。稻渔综合种养作为我国当前生态循环农业经济的主要模式之一,在保障粮食安全、拓展渔业发展空间、促进农业增效农民增收中发挥了重要作用[5-6]。其中稻虾(克氏原螯虾,Procambarus clarkii)综合种养因其操作简单、收益较高,已成为我国最受欢迎、应用面积最大、总产量最高的稻渔综合种养模式[7-8]。但克氏原螯虾作为底栖生物,主要占据水体底层空间,稻虾综合种养模式下养殖环沟中上层环境并未得到有效利用。鉴于此,将黄鳝网箱养殖与稻虾综合种养立体结合,即在稻虾综合种养环沟内设置黄鳝养殖网箱,黄鳝产生的残饵粪便被克氏原螯虾利用,同时克氏原螯虾的活动可增加水体溶氧,促进系统物质循环转化,改善养殖环境。为明确稻虾鳝立体综合种养模式水体微生物特征,通过Illumina MiSeq 分析方法,对河南信阳罗山县稻虾鳝立体综合种养模式下稻田水体环境中微生物群落结构和多样性进行分析及菌群功能预测,旨在为科学评价稻虾鳝立体综合种养模式提供科学依据。
1 材料和方法
1.1 样品采集
试验于2020 年8—10 月在河南省信阳市罗山县商湖农业公司进行,选取2 hm²稻虾综合种养实验基地,在2 个相邻的环沟内共设置浮式联排网箱30 个,每沟15 个。每个网箱面积约1 m2,箱内水深约30 cm,网箱中栽种水花生,面积约为网箱面积的90%~95%。试验在网箱内(T1)、网箱外(T2)、邻侧环沟(T3)、对侧环沟(T4)设置4 个采样点(图1),于8月22日、9月6日、9月24日、10月11日、10月25日在4个采样点分别采集水样1 L。其中水体溶解氧(DO)、酸碱度(pH 值)用便携式水质分析仪现场检测,总磷(TP)、总氮(TN)、硝酸盐氮(NO3--N)、氨态氮(NH4+-N)、亚硝酸盐氮(NO2--N)、高锰酸盐指数(CODMn)指标通过将水样低温带回检测。其中TP、TN 含量测定参照GB/T 11894—1989、GB/T 11893—1989 中的方法,NO3--N、NO2--N 含量测定参照HJ/T 346—2007、GB/T 7493—1987 中的方法,NH4+-N 含量测定参照GB/T 7479—1987中的方法,CODMn测定参照GB 11892—1989 中的方法。水化学指标检测后,从剩余水样取500 mL 用0.22 μm 硝酸纤维素滤膜进行过滤,滤液保存于-20 ℃冰箱,待所有样本采集完成后统一进行微生物多样性分析。第1次采样编号为T1_1、T2_1、T3_1、T4_1,共采样5 次,编号以此类推。

图1 稻虾鳝立体综合种养试验田采样点Fig.1 Sampling points in test field of rice-crayfish-eel coculture system
1.2 测序方法
对16S rRNA 基因的V4—V5 区进行扩增,上下游引物分别为515F(5'-GTGCCAGCMGCCGCGG-3')、907R(5'-CCGTCAATTCMTTTRAGTTT-3'),扩增 体 系 共20 μL:4 μL 5×FastPfu Buffer,2 μL 2.5 mmol/L dNTPs,5 μmol/L 上下游引物各0.8 μL,0.4 μL FastPfu 聚合酶,0.2 μL BSA,10 ng DNA 模板(10 ng/μL),ddH2O 补至20 μL。PCR 反应条件:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,进行27 个循环;72 ℃终延伸10 min。利用2%琼脂糖凝胶电泳对PCR 产物进行检测和回收,采用Illumina MiSeq测序平台进行双端测序[9]。
1.3 数据处理和分析
对原始测序序列使用Trimmomatic 软件进行质量控制,通过FLASH 软件进行拼接,使用UPARSE软件根据97%的相似度对序列进行操作分类单元(Operational taxonomic units,OTU)聚 类,使 用UCHIME软件剔除嵌合体,获得样本有效序列,利用RDPclassifier 对每条序列进行物种分类注释,在门、纲、属等不同分类水平上对各样本的群落组成进行统计分析,与Silva 数据库(SSU128)进行比对,设置比对阈值为70%。群落结构采用柱状图描述,表示各样品中不同菌群操作分类单元所占百分比。
采用香农(Shannon)指数、Ace 指数、辛普森(Simpson)指数、Chao 指数及覆盖度(Coverage)评价不同采样点水体微生物群落的Alpha 多样性;使用菌群代谢功能预测工具PICRUST 对OTU 丰度进行标准化,在直系同源基因簇(COG)中对比每个OTU对应的基因名称,获得OTU 对应的COG 家族信息,计算各COG 的丰度,从直系同源蛋白分组比对(eggNOG)数据库解析各COG 的描述信息和功能信息,得到功能丰度谱。利用SPSS 19.0软件对多样性指数进行非参数检验和T检验,分析不同采样点的微生物多样性指数差异。利用SPSS 19.0 软件对水体化学指标和门水平群落结构进行冗余分析(RDA),得到影响群落结构的主要环境因子。
2 结果与分析
2.1 稻虾鳝立体综合种养水体测序结果分析
通过Illumina MiSeq 测序平台对T1、T2、T3、T4共4 个采样点20 个样本的16S rRNA 基因V4—V5区进行测序,原始序列数据经过质控和过滤后共得到有效序列1 130 370 条,按97%相似度水平聚类,得到3 737 个OTU,共鉴定出47 门、137 纲、309 目、509 科、929 属、1 697 种。各样品的OTU 数、序列数及Alpha多样性指数见表1。
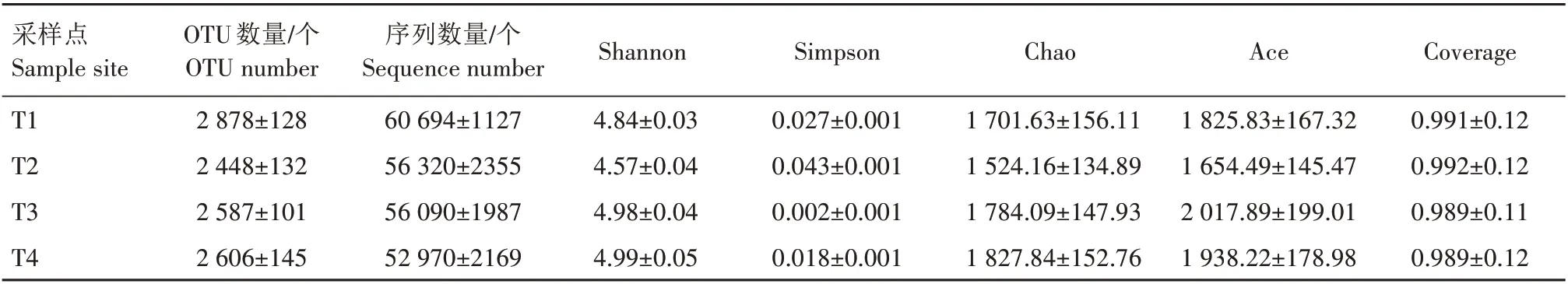
表1 稻虾鳝立体综合种养水体测序情况和Alpha多样性统计Tab.1 Sequencing and alpha-diversity of samples in rice-crayfish-eel coculture system
Alpha 多样性主要对单样品的微生物组成进行分析,可以反映样本的群落结构和丰富度,常用的度量指标包括Shannon 指数、Chao 指数、Ace 指数、Simpson 指 数 和Coverage 值。其 中,Shannon 指 数、Chao 指数、Ace 指数越高表明菌群的多样性和丰度越高,Simpson 指数值越低表明菌群的多样性越高,Coverage 值越接近于1 表明测序深度越合理[9]。从表1 可以看出,各样本Coverage 值均高于0.989,表明测序样本覆盖率较好,序列未被测出的概率较低。T3、T4 采样点的Shannon 指数、Chao 指数、Ace指数高于T1、T2 采样点,Simpson 指数和Coverage 值低于T1、T2 采样点,配对样本的非参数检验和T检验均表明,各个采样点间的多样性指数均未见显著性差异。
从图2 可以看出,各样本共有OTU 为157 个,T3_3 独有的OTU 最少,为0 个,T1_5 独有的OTU 最多,为211 个。从采样时间来看,第5 次采样的样本较前期样本具有较高的独有OTU 数量,表明随着养殖过程的进行,采样点间逐渐表现出较大的微生物群落组成差异。

图2 稻虾鳝立体综合种养水体OTU的Venn分析Fig.2 Venn analysis of OTU in samples of rice-crayfish-eel coculture system
2.2 稻虾鳝立体综合种养水体微生物组成分析
2.2.1 基于门水平的水体微生物组成分析 测序结果显示,采样点微生物在门水平上主要包括变形菌门(Proteobacteria)、蓝细菌门(Cyanobacteria)、放线菌门(Actinobacteriota)、厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidota)、浮霉菌门(Planctomycetota)、绿弯菌门(Chloroflexi)和Armatimonadota 共8 个菌门。除Armatimonadota之外,其余7种在T1、T2、T3采样点丰度均超过1%;T4 采样点所有菌门丰度均超过1%。T1、T4 采样点优势菌门(丰度>10%)包括变形菌门、蓝细菌门、放线菌门、厚壁菌门,T2、T3 采样点优势菌门(丰度>10%)包括变形菌门、蓝细菌门、放线菌门。其中,T1采样点变形菌门丰度(50.25%)高于T2(41.55%)(P<0.05)、T3(37.38%)(P<0.05)、T4 采样点(38.24%)(P<0.05),T3 采 样 点 蓝 细 菌 门 丰 度(31.29%)高 于T1(14.23%)(P<0.05)、T2(30.83%)(P>0.05)、T4采样点(24.11%)(P<0.05),T4采样点放线菌门丰度(15.82%)高于T1(11.60%)(P<0.05)、T2(10.53%)(P<0.05)、T3 采 样 点(12.30%)(P<0.05)(图3)。
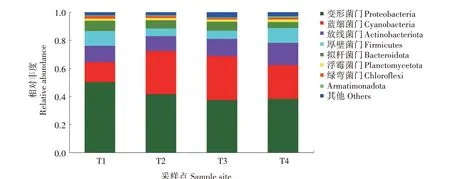
图3 稻虾鳝立体综合种养水体微生物在门水平分布情况Fig.3 Distribution of microflora at the phylum level in rice-crayfish-eel coculture system
2.2.2 基于纲水平的水体微生物组成分析 如图4所示,在纲水平上,采样点微生物组成主要包括γ-变 形 菌 纲(Gammaproteobacteria)、蓝 细 菌 纲(Cyanobacteriia)、α-变形菌纲(Alphaproteobacteria)、放线菌纲(Actinobacteria)、拟杆菌纲(Bacteroidia)、梭状芽孢杆菌纲(Clostridia)、普朗克菌纲(Bacilli)、嗜 酸 菌 纲(Acidimicrobiia) 、浮 霉 菌 纲(Planctomycetes)、绿弯菌纲(Chloroflexia)、嗜热菌纲(Thermoleophilia)共11 个菌纲,T1 和T4 采样点丰度超过1%的菌纲有10种,T2和T3采样点丰度超过1%的菌纲有9 种。4 个采样点相对丰度最高的3 个菌纲均为γ-变形菌纲、蓝细菌纲、α-变形菌纲,T1、T2 和T4 采样点的第一优势菌纲为γ-变形菌纲,T3的第一优势菌纲为蓝细菌纲。γ-变形菌纲在各采样点的相对丰度分别为T1(36.50%)>T2(31.20%)>T3(27.03%)>T4(24.07%),蓝细菌纲在各采样点的相对丰度分别为T3(31.20%)>T2(30.80%)>T4(24.05%)>T1(14.20%),a-变形菌纲在各采样点的相对丰度分别为T4(14.10%)>T1(13.60%)>T2(10.33%)>T3(10.30%)(图4)。

图4 稻虾鳝立体综合种养水体微生物在纲水平分布情况Fig.4 Distribution of microflora at the class level in rice-crayfish-eel coculture system
2.3 稻虾鳝立体综合种养水体微生物群落结构分析
NMDS非度量多维尺度分析对不同物种微生物群落间物种多样性进行组间比较分析,样本以点显示,点之间的距离表明相互关系的远近,由此判断各样本群落组成的相似性[10]。如图5 所示,从空间的角度看,各采样点菌群均能较好地自成区域,但也有部分重叠。从时间的角度看,随着养殖进行,同次采样各个点的离散程度越来越大,表明养殖后期,4个采样点的微生物群落结构发生了较大改变。可见,黄鳝养殖网箱的设置显著改变了其周围区域的小环境和微生物群落结构。
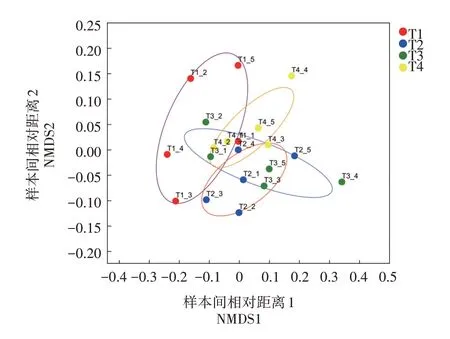
图5 稻虾鳝立体综合种养水体微生物NMDS非度量多维尺度分析(胁强系数:0.093)Fig.5 NMDS multidimensional scaling analysis of microflora in rice-crayfish-eel coculture system(stress:0.093)
2.4 稻虾鳝立体综合种养水体COG功能预测
通过PICRUST 工具对样本微生物群落进行COG 功能预测和分类统计,结果表明,4个采样点菌群均具有22个功能分类簇,且均在氨基酸转运和代谢、能量产生和转化及翻译、核糖体结构和生物发生3个功能类群的相对丰度较高,在细胞运动、细胞周期控制和细胞分裂、染色体分配功能菌群相对丰度较低,不同采样点之间功能分类均无显著差异(图6)。
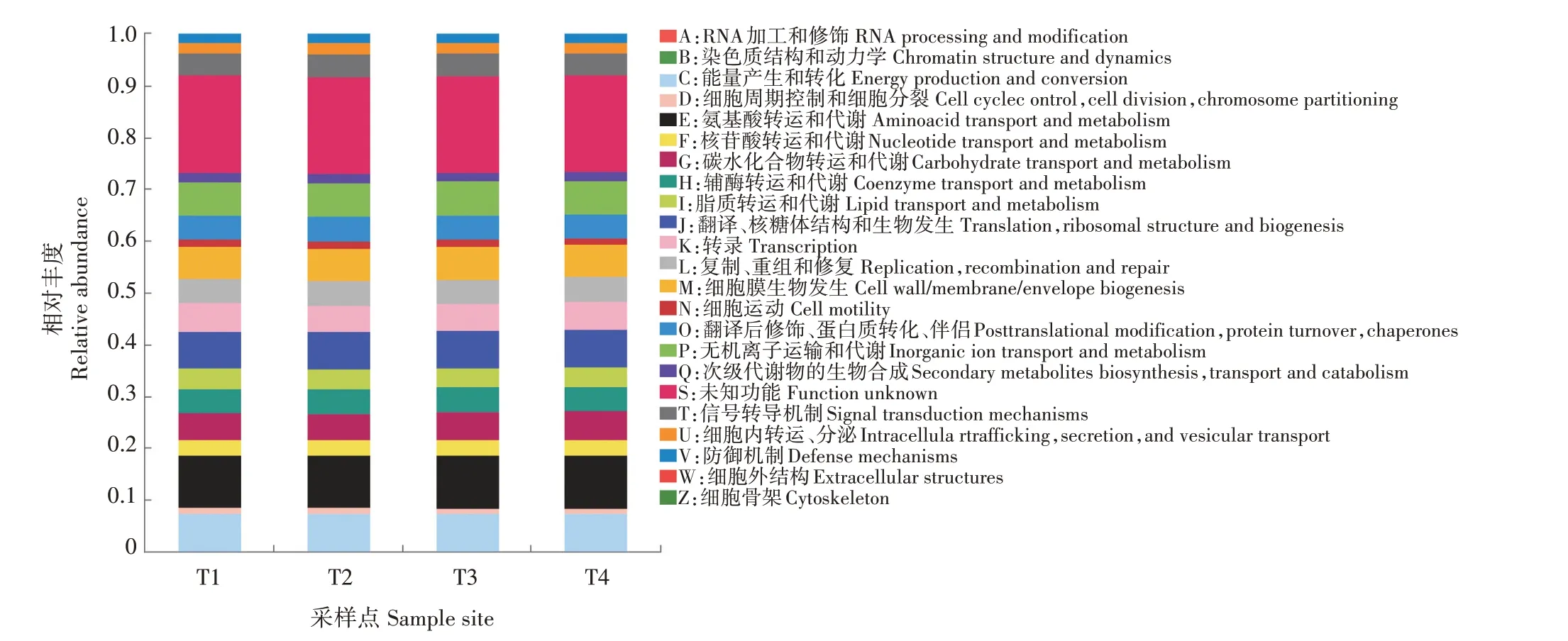
图6 稻虾鳝立体综合种养水体微生物群落COG功能统计Fig.6 The COG function classification of microflora in rice-crayfish-eel coculture system
2.5 稻虾鳝立体综合种养水体环境因子和微生物群落的RDA分析
对水体环境因子和门水平微生物群落进行RDA 分析,结果表明,前2 个排序轴的特征值分别为35.71%、8.33%,NO2--N 含量对水体微生物群落影响最大,其次是TN 和NH4+-N 含量,CODMn对水体微生物群落影响最小。其中,NO2--N 含量与TN、NH4+-N 含量呈负相关,与TP、NO3--N 含量和CODMn呈正相关;变形菌门丰度与TN、NH4+-N 含量呈正相关,与TP、NO3--N、NO2--N 含量和CODMn呈负相关;蓝细菌门丰度与TP、NO3--N、NO2--N 含量和CODMn呈正相关,与TN、NH4+-N 含量呈负相关;放线菌门丰度与TN、TP 含量和CODMn呈正相关,与NO3--N、NH4+-N含量呈负相关(图7)。

图7 稻虾鳝立体综合种养水体环境因子与微生物菌群RDA分析Fig.7 RDA of environmental factors and microbial flora in rice-crayfish-eel coculture system
3 结论与讨论
微生物在养殖环境的物质循环和能量流动过程中发挥着重要作用,微生物群落组成和多样性与环境异质性密切相关,常可作为环境变化的指示因子[11]。微生物群落结构的多样性会影响水体理化环境因子和养殖效果[12]。对养殖环境中微生物多样性进行研究,不仅可以为养殖过程中病害防控提供参考,还能对养殖环境微生态调控提供理论依据[13-14]。本研究将网箱养殖黄鳝和稻虾综合种养立体结合,通过Illumination MiSeq测序技术对不同采样点水体的微生物群落多样性指数进行配对样本的非参数检验和T检验,结果表明,各个采样点间并未表现出微生物多样性指数的显著差异,表明网箱设置并未显著改变各个区域的微生物多样性指数。Venn 分析结果显示,随着养殖进行,不同采样点拥有更多的独有OTU 数,即微生物菌群组成在各采样点表现出较为明显的差异。
对优势物种的分析显示,门水平上,变形菌门、蓝细菌门、放线菌门均为各采样点的优势菌门。变形菌门是细菌中最大的门,包含多种代谢种类细菌,广泛参与环境中的物质代谢[15]。前人研究表明,水环境中多数参与有机物降解和生物脱氮除磷过程的细菌属于变形菌门,如硝化细菌、反硝化细菌等[16]。本研究中,T1 采样点变形菌门的相对丰度高达50.25%,显著高于其他3 个采样点,推测网箱内因代谢物质积累较多造成变形菌门积累。蓝细菌门和放线菌门的丰度,均与水体中N、P 含量密切相关,水体中TP含量的增加会促进蓝细菌和放线菌的生长繁殖[4,17]。聂志娟等[18]的研究结果表明,稻鲤共生系统和单稻模式水体中优势菌门均为变形菌门、放线菌门等,与本研究结果一致。纲水平上,γ-变形菌纲、蓝细菌纲和α-变形菌纲为不同采样点的共同优势菌纲,与门水平分析结果相吻合。γ-变形菌是海洋中普遍存在的细菌,多样性高,多数为兼性异养菌,以有机物作为碳源,是系统中降解高锰酸盐的主要参与者[4,13]。α-变形菌纲是淡水细菌群落中典型的优势类群,包括能与植物共生的固氮细菌,可为水体提供更强的固氮能力[3]。推测在稻虾鳝养殖系统内,细菌能够较好地进行水体自净,并广泛参与N 循环过程,对维持系统健康具有重要的作用。
微生物群落组成对环境差异有着较强的敏感性,菌群结构与环境因素密切相关且相互影响[19-20]。本研究中,T1、T2、T4采样点具有较明显的群落组成差异,尤其T1 和T2 采样点的群落组成几乎未有重叠,表明网箱养殖黄鳝对水环境中微生物的群落组成影响明显,与本研究中Alpha 多样性分析结果一致。对水体环境因子和微生物群落进行RDA 分析结果表明,NO2--N、TN、NH4+-N 含量等指标与微生物群落呈现出较强的相关性,CODMn相关性最小,这与陈玲等[20]的研究结果一致,表明氮元素对水体微生物群代谢功能具有重要意义。同时,α-变形菌纲在4个采样点中均为优势菌纲,推测与系统氮循环相关的菌群如硝化菌群、反硝化菌群多样性和丰度均较丰富,下一步需对N 循环相关菌群进行深入研究。
闫法军等[21]的研究结果表明,微生物群落的代谢功能特征能够反映环境的健康状况。基于16S rDNA 进行COG 功能预测的比较分析可以观测不同组别样品之间微生物群落的功能差异,是研究群落样本与环境适应性变化的有效手段[10]。对稻虾鳝立体综合种养系统中COG 功能预测分析表明,系统水体共有22个功能簇,其中与氨基酸转运和代谢有关的菌群丰度最高,其次为与能量产生和转化有关的菌群,与细胞周期控制、细胞分裂、染色体分配有关的菌群丰度最低。表明水体细菌代谢活动十分活跃,但细菌的更新迭代较慢,水体易老化,实际生产中应注意加强水体的更换,促进系统的物质循环。
本研究通过在稻虾共生系统中加入黄鳝网箱养殖,进一步利用水体空间,丰富了系统的生物多样性,但因网箱内代谢废物积累较多,物质循环减弱,造成特定菌群富集,降低了网箱内微生物多样性,形成了不同区域内的微生物组成空间异质性。同时,由于网箱内外水体交换能力较差,网箱内较高的营养盐积累不能得到网箱外水体的有效缓冲,造成网箱内变形菌门丰度较高,进一步需采取间隔设置网箱或加大网布孔径等措施加强网箱内外水体交换,同时,需对N 循环相关的菌群如硝化菌群、反硝化菌群的丰度和优势种进行测定分析,完善稻虾鳝综合种养系统内部微生物评价体系,为传统稻渔综合种养模式创新、提质增效提供参考。
