不同输入电压等离子体诱变对双对栅藻生长、光合性能及油脂积累的影响
2022-02-12位文倩李鹏飞
黄 峰,孙 昕,位文倩,李鹏飞
(西安建筑科技大学 环境与市政工程学院,西北水资源与环境生态教育部重点实验室,西安 710055)
微藻是一种生长速度快、生存环境要求低、光合效率高、含有较高油脂的单细胞生物,在其生长过程中也能固定二氧化碳,从而减轻全球气候变暖问题[1-2]。近年来,微藻作为一种极具有潜力的生物燃料原料被广泛讨论,微藻生物质能源的发展对于我国在2060年实现碳中和的目标具有重要意义。
尽管微藻作为生物柴油原料具有很多的优势,但是其商业化生产仍然存在着一些障碍,比如低脂质产率和高运营成本的问题,导致其发展水平远远低于预期[3]。因此,获得高生物量和高油脂含量的藻株对降低微藻生物柴油的生产成本至关重要[4]。目前已经开发了多种微藻培养策略来增强微藻的油脂含量,例如化学胁迫,常见的氮或磷胁迫就是一种简单经济的提高微藻油脂含量的方法[5-6],但是营养元素的缺乏会抑制藻细胞生长,降低生物量,显著降低油脂产率[7]。微藻诱变育种技术已经被广泛应用于许多不同藻类,以获得能稳定遗传的高脂质产率突变株[8]。常压室温等离子体诱变作为一种新型的物理诱变技术,其与紫外线、X射线、碳离子束等传统诱变技术相比,操作更加简单且安全,等离子体产生的活性粒子可以透过细胞膜与胞内脂质、蛋白质等发生反应从而引起DNA突变,突变效率也远远高于其他诱变[9-10]。曹旭鹏等[11]在输入电流1.4 A、诱变时间30 s的条件下,获得的等鞭金藻突变株油脂含量提高了11%。Fang等[12]在输入功率100 W、作用时间60 s条件下,诱变所得的突变株碳水化合物较原始株提高了22%。在不同条件下,等离子体设备可能会产生不同活性和浓度的等离子体,而等离子体对不同的微藻有其独特的作用机制,因此有必要寻找针对不同微藻的适宜诱变条件。
目前等离子体诱变应用于双对栅藻的研究较少,本研究通过在不同输入电压条件下产生等离子体,对双对栅藻进行诱变,确定了最适的诱变条件,并通过测定突变株生物量、光合性能、生化组分等指标,分析了等离子体诱变促进双对栅藻油脂积累的可能途径,为后续的高脂质产率微藻的定向基因改造提供基础。
1 材料与方法
1.1 实验材料
1.1.1 藻种及培养基
双对栅藻,购于中国科学院水生生物研究所淡水藻种库。
液体培养基为BG11培养基,在BG11培养基中加入1.5%琼脂粉用作固体培养基,每次配制完成后调节pH为7,121℃灭菌30 min。
1.1.2 仪器与设备
CTP-2000K等离子体诱变仪,UV-6000紫外分光光度计,F-7000荧光分光光度计(日本日立),IMAGING-PAM调制叶绿素荧光成像系统(Heinz Walz GmbH公司)。
1.2 实验方法
1.2.1 双对栅藻的培养
将含有100 mL双对栅藻藻液的250 mL培养瓶用无菌透气封口膜封口,置于恒温光照培养箱(温度25℃)培养,平均光照强度80 μmol/(m2·s),光照周期12 L/12 D,每天随机调换培养瓶的位置并摇动3~4次使藻液分布均匀。
1.2.2 等离子体诱变及筛选
将藻液浓度调整至约1×106mL-1,取100 μL藻液置于灭菌的培养皿,在发射器与样品的距离为2 mm、电流1 A、空气流量5 L/min、诱变时间60 s条件下,分别调节电压为0、20、40、60、80、100、120、140、160、180、200 V,对双对栅藻进行诱变,诱变后立即测定叶绿素荧光参数最大光化学量子产量(Fv/Fm)。将诱变后的藻液稀释涂布于固体平板上培养,以存活率低于10%为指标初选藻株进行孔板培养,以培养3 d和8 d的藻液在680 nm处的吸光度计算比生长速率,测定培养8 d藻液的甘油三酯含量。经两步筛选即第一步筛选出比生长速率和相对荧光值均高于原始株的突变株,第二步选择比生长速率或者相对荧光值特别突出的突变株,再将筛选出的突变株重复上述孔板培养过程,验证稳定性,最终筛选出遗传稳定的优势突变株[13]。筛选的突变株按1.2.1方法培养35 d。
1.2.3 微藻生物量的测定
生物量即单位体积藻液内的藻细胞干重,生物质产率即单位时间内生物量的增长量。将稳定期双对栅藻藻液分别稀释成20 mL不同质量浓度梯度的藻液,测定每个质量浓度下的藻液在680 nm处的吸光度(A)和经离心干燥恒重后的生物量(Y),绘制生物量与吸光度之间的校准曲线,得到曲线方程为Y=310.4A-19.7(R2=0.999)。通过测定不同培养时间下藻液的吸光度,代入曲线方程中计算生物量。
1.2.4 叶绿素荧光参数及色素含量测定
采用调制叶绿素荧光成像系统测定双对栅藻的最大光化学量子产量(Fv/Fm)、实际量子产量(Y(II))[14]。
取5 mL藻液于装有5 mL 95%乙醇溶液的离心管中,在75℃水浴15 min后,于5 000 r/min离心10 min,取上清液,测定其在665、649、470 nm处的吸光度,再测定其经盐酸化后相应波长处的吸光度,以酸化前后吸光度差计算色素含量,具体方法参照文献[15]。
1.2.5 甘油三酯产量和总脂含量测定
采用优化的尼罗红法[16]测定双对栅藻生长过程中甘油三酯产量,采用改进的Bligh-Dyer法测定其总脂含量[17]。
1.2.6 碳水化合物含量和蛋白质含量测定
采用苯酚-硫酸法[18]测定双对栅藻碳水化合物含量(以葡萄糖为标准溶液)。采用考马斯亮蓝法[19]测定双对栅藻蛋白质含量(以牛血清蛋白为标准溶液)。
2 结果与讨论
2.1 不同输入电压对双对栅藻生长的影响
叶绿素荧光可以作为光合突变体的筛选指标[20],Fv/Fm反映植物的潜在光合能力,常用来反映微藻生长中的光合活性变化以及所受的外界环境胁迫程度[21]。不同输入电压下诱变对双对栅藻的Fv/Fm和存活率的影响如图1所示。

图1 不同输入电压下诱变双对栅藻的Fv/Fm、存活率
从图1可以看出,Fv/Fm随着输入电压的升高呈现不断下降的趋势,说明在短时间内双对栅藻所受到的胁迫程度逐渐加深。当输入电压为140 V时,Fv/Fm较初始值下降73.2%;当输入电压高于160 V时,Fv/Fm已接近0,这说明藻细胞受到较大的损害可能无法继续生长。从存活率曲线可以看出,当输入电压从60 V升至140 V时,存活率显著下降,从83.9%下降至5.4%,160 V时存活率仅为2.5%,180 V以上时几乎没有存活藻株。图1所示结果说明,随着输入电压增大,藻细胞所受损害的程度加剧,可能是等离子体系统所产生的活性粒子浓度增加导致。根据诱变育种理论,通常存活率小于10%时可获得更高的正突变率[22]。在本实验中,为了确保藻细胞光系统不受致命损害,平板上有足够的可供挑选的藻株,选择输入电压140 V条件下存活的藻株进行两步筛选,最终筛选出比生长速率和甘油三酯含量均高于原始株的突变株T-2、T-5和仅甘油三酯含量显著提高的突变株T-6进行扩大培养,并进一步分析原始株与突变株之间的差异。
2.2 诱变对双对栅藻生物量积累的影响(见图2)
从图2可以看出:在开始培养期间,原始株与突变株均以较高的速率增长;培养7 d左右,突变株T-2、T-5生长优势逐渐显现,直至培养35 d,突变株T-2、T-5的最高生物量分别达到788.9、764.1 mg/L,分别比原始株(672.5 mg/L)提高17.3%、13.6%。在培养前期,突变株T-6生物量积累慢于原始株,但T-6在培养29 d之后仍保持缓慢生长,而原始株此时已达到稳定期,在培养结束时T-6生物量达到699.5 mg/L。实验表明,在适宜的输入电压下产生等离子体,可以诱变微藻,促进微藻生物量积累。原因可能是在合适的电压下,产生了含有适宜浓度的活性氧或活性粒子的等离子体。一方面,等离子体诱变使突变株的对数生长期延长,从而提高生长末期的生物量;另一方面,等离子体在损伤细胞的同时,也激发了更大的细胞自身修复作用,从而刺激藻细胞增殖,这与Rana等[23]的研究结果相似。等离子体诱变应用于其他微藻中也有不同的效果,如:Cao等[24]利用等离子体诱变获得的小球藻突变株平均生物量较原始株增加了7.75%;Fang等[12]通过等离子体诱变螺旋微藻获得1株突变株,生长速率提高了10.3%。
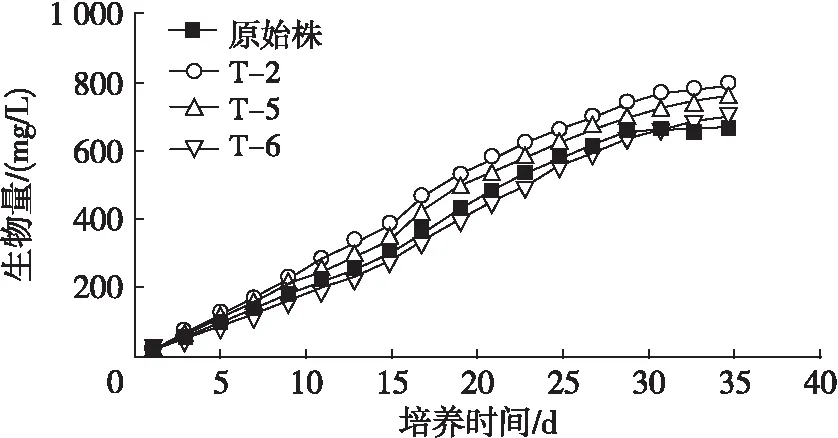
图2 突变株与原始株的生物量积累曲线
2.3 诱变对双对栅藻色素含量及光合性能的影响
为了进一步研究突变株与原始株在生物量增长上的差异,对突变株与原始株的色素含量和光合性能参数进行了测定。光合色素是微藻在光合作用中捕获光能的色素,对微藻生长起重要作用[25]。培养35 d的突变株与原始株的色素含量见图3。
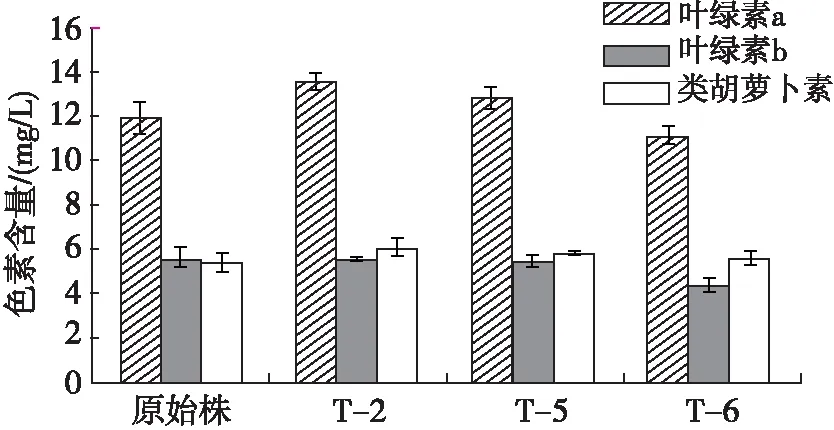
图3 培养35 d的突变株与原始株色素含量
从图3可以看出:在培养35 d时,突变株T-2、T-5的叶绿素a含量分别为13.5、12.8 mg/L,略高于原始株的11.8 mg/L,叶绿素b含量无明显差异;突变株T-2、T-5、T-6的类胡萝卜素含量分别为6.1、5.8、5.6 mg/L,均较原始株的5.3 mg/L有所增加,这可能是突变株对等离子体诱变所做出的抵抗应激反应的结果。因此,可能是光合色素含量增加使藻细胞光合活性提高,从而提高了突变株生物量的积累。
突变株与原始株实际量子产量(Y(Ⅱ))与最大光化学量子产量(Fv/Fm)变化趋势如图4所示。

从图4可以看出:在生长前期,Fv/Fm与Y(Ⅱ)均有短暂的下降,随着藻细胞增殖,细胞密度不断增大,捕获光能的能力也逐渐加强,二者呈现出上升趋势并在培养22 d达到最大值;在培养后期,二者均有小幅度的下降,可能是后期藻细胞大量增殖,藻类之间发生遮蔽效应,导致光照不均,或是后期营养物质减少,引起光合效率下降。在整个培养过程中,突变株T-2的Y(Ⅱ)均高于原始株,突变株T-6的Y(Ⅱ)均小于原始株,与图2所示生物量变化趋势类似。在培养后期,4个藻株的Fv/Fm均保持在0.5~0.7正常范围内[26],说明藻株基本适应了等离子体辐射产生的胁迫。
2.4 诱变对双对栅藻脂质积累的影响
突变株与原始株的甘油三酯积累曲线如图5所示。

图5 突变株与原始株甘油三酯积累曲线
从图5可以看出,甘油三酯积累与生物量积累保持着一致性,从培养5 d开始,甘油三酯积累速率明显加快,但是突变株与原始株在甘油三酯积累上的差异也逐渐加大,突变株T-6在甘油三酯积累上的优势最为突出。培养30 d后,藻株生长逐渐进入稳定期,甘油三酯的积累量也趋于稳定,突变株T-2、T-5、T-6 最高甘油三酯产量分别达到229.7、263.0、283.6 mg/L,较原始株的165.5 mg/L分别提高了38.8%、58.9%、71.3%。
甘油三酯产量反映藻细胞内中性脂含量,还需结合总脂含量和总脂产率作为评估微藻生产生物柴油潜力的重要指标[27]。培养35 d的突变株与原始株生物量、生物质产率、总脂含量和总脂产率如图6所示。
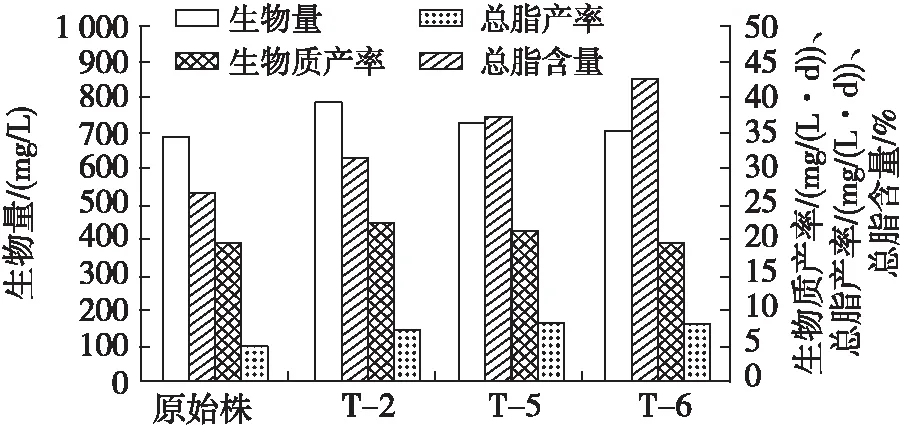
图6 培养35 d的突变株与原始株生物量、生物质产率、总脂含量、总脂产率
由图6可以看出,原始株和突变株T-2、T-5、T-6的总脂含量分别为25.9%、32.4%、38.2%、42.7%。突变株T-2、T-5、T-6的总脂产率分别达到7.3、8.3、8.5 mg/(L·d),较原始株的5.0 mg/(L·d)分别提高了46%、66%、70%。说明在此条件下的等离子体诱变明显促进了双对栅藻的总脂积累。
2.5 诱变对双对栅藻碳水化合物和蛋白质积累的影响(见图7)
微藻的生化组分主要包括脂质、蛋白质、碳水化合物,三者之间保持着动态的平衡,脂质含量的提升必然会导致其他成分的降低[28]。从图7(a)可以看出,随着培养时间的延长,碳水化合物不断积累,原始株和突变株T-2、T-5、T-6在生长末期碳水化合物含量分别为79.6、89.3、83.6、81.5 mg/L,分别占生物量干重的11.8%、11.3%、10.9%、11.7%,说明诱变并没有明显影响双对栅藻的碳水化合物合成。从图7(b)可以看出,突变株蛋白质含量呈现先上升后下降的趋势,突变株T-2、T-5、T-6蛋白质含量在培养21 d时达到最大,分别为175.7、170.3、166.1 mg/L,之后3株突变株蛋白质含量发生了不同程度的下降,待培养结束时,突变株T-2、T-5、T-6蛋白质含量较原始株分别下降了15.2%、24.4%、31.8%。这说明诱变可能影响了双对栅藻的蛋白质代谢途径,在培养后期突变株可能是以消耗蛋白质为代价来促进油脂的积累。Williams等[29]通过监测在不同氮磷比的培养基中生长的微拟球藻发现,在低氮磷比的情况下,蛋白质含量较正常时下降了30.3%,而总脂质是对照组的2倍。藻细胞内不同组分之间的相互变化,可能是由于等离子体所产生的活性粒子对藻细胞内调控相关组分代谢的部分基因产生突变,导致油脂快速积累,后续还需对其进行转录组分析进一步确定差异基因表达,以便更好地阐明诱变对双对栅藻代谢途径的影响。

3 结 论
对双对栅藻进行不同输入电压条件下的等离子体诱变,确定了最适诱变条件为140 V,筛选出突变株T-2、T-5、T-6,分析了它们与原始株之间的生物量积累、光合性能以及油脂积累的差异。在培养至35 d时,突变株T-2、T-5生物量分别达到788.9、764.1 mg/L,较原始株(672.5 mg/L)分别提高了17.3%、13.6%,突变株T-6的生物量为699.5 mg/L,较原始株略有提升,诱变并未对突变株的叶绿素荧光特性参数产生较大影响,突变株均保持着良好的光合性能。突变株T-2、T-5、T-6培养35 d的总脂产率较原始株分别提高了46%、66%、70%,蛋白质含量较原始株都有不同程度的下降。突变株T-5的总脂产率与T-6相差不大,但其生物量比T-6高9.24%,因此突变株T-5更具产业化潜力。
