毛竹AUX1/LAX候选基因家族鉴定与表达分析
2022-02-10杨乐,张康,李龙
杨 乐,张 康,李 龙
(西北农林科技大学 林学院,陕西 杨陵 712100)
生长素是植物中发现最早且最为重要的一类激素。生长素合成于幼嫩的组织中,通过AUX1/LUX、PIN等蛋白家族转移至各个不同的部位,在植物的形态建成、细胞生长、根系发育调控以及抗逆胁迫中均发挥着重要的作用[1-2]。其中,生长素输入载体在生长素浓度梯度的调控及维持中具有重要作用[2]。在拟南芥(Arabidopsisthaliana)中发现共有4个生长素输入载体,包括AUX1(AUXINRESISTANTI)、LAX1(LIKEAUXINRESISTANT 1)、LAX2和LAX3[3]。其中,AUX1是最早被发现的生长素输入载体,氨基酸序列分析表明,AUX1与通透酶具有较高的同源性,该蛋白可以高效、特异的介导生长素从质膜外进入细胞内[4-5]。在拟南芥中,AUX1在根的表皮细胞、中柱细胞及初生韧皮部细胞的顶部中表达。aux1的突变会导致个体重力性丧失,外源施加NAA能恢复突变表型,而外施2,4-D并不能恢复突变体表型[6]。LAX1/2/3是高度保守的跨膜蛋白,广泛参与植物根的向地性、叶序发育、侧根形成等生物学过程。aux1和lax1/2/3的四突变体与野生型相比,叶序排列发生改变,并且胚根的发育极度混乱[7]。
毛竹(Phyllostachysedulis)是植物界生长最快的植物,在分类学上属于禾本科、竹亚科、刚竹属,是东亚地区重要的林业资源之一。竹材可用于造纸、家具制作、建筑等领域,竹笋可用于食用,每年可产生1 840亿美元的林业产值[8]。此外,毛竹根系发达,具有很强的固水能力,且具有很强的根鞭繁殖能力,因此毛竹在解决生态环境恶化、资源匮乏等方面具有不可替代的作用[8]。在气候适宜的春季,毛竹单日最高生长高度可达1.5 m,在短短45~60 d便可完成高达15~20 m的秆型生长[9-11],因此毛竹奇特的笋-竹生长模式一直是研究热点。
目前,在拟南芥、水稻(Orazasativa)等模式植物中虽已有AUX1/LAX的相关报道,但在毛竹中关于AUX1/LAX的研究仍是一个空白,毛竹中共有多少个AUX1/LAX候选基因,他们是否参与毛竹笋的生长发育尚不明确。本研究利用毛竹基因组信息,结合AUX1/LAX候选基因特征,在全基因组水平对毛竹AUX1/LAX候选基因进行了鉴定,并对AUX1/LAX候选基因的数量、进化关系、基因结构和不同组织,特别是不同生长发育时期竹笋中的表达模式进行了分析,为将来进一步开展PheAUX1/LAX的功能和调控机制研究奠定理论基础。
1 材料与方法
1.1 毛竹AUX1/LAX候选基因鉴定
拟南芥以及水稻的AUX1/LAX蛋白序列分别下载于The Arabidopsis Information Resource (TAIR)数据库和Rice Genome Annotation Project数据库。毛竹的基因组数据下载于Gigadb Database[12]。以拟南芥和水稻AUX1/LAX蛋白序列为种子序列,阈值设置为e-7,在构建的毛竹本地数据库中进行BLASTP同源比对,所得序列为毛竹潜在AUX1/LAX基因家族成员,之后再通过Pfam数据库验证这些序列所包含的结构域,只有包含AUX1/LAX蛋白典型结构域的序列才被认为是毛竹AUX1/LAX候选基因家族成员[13-14]。
1.2 系统进化树构建
利用ClustalW对AUX1/LAX氨基酸序列进行多序列比对分析,比对结果通过IQ-Tree构建系统进化树。执行参数为最大似然法,Bootstrap 重复设置为1 000次,采用最适合建树模式(best fit tree model),经筛选发现JTT+R3为最适合建树模式[15-16]。
1.3 基因结构和基序分析
利用在线软件MEME对毛竹Aux1/LAX蛋白保守基序进行分析,检索的最大motif 数设置为6,其他参数选择默认参数[17]。在毛竹基因组数据库下载GFF文件,将GFF文件输入TBTools分析候选Aux1/LAX的基因结构[18]。
1.4 毛竹Aux1/LAX候选基因染色体定位和共线性分析
根据毛竹基因组GFF注释文件获取AUX/LAX候选基因家族成员的染色体位置信息;利用MCScanX软件计算并获取共线性区块及基因的串联重复信息[19];利用Circos软件对毛竹AUX1/LAX候选基因的染色体位置信息、共线性关系进行可视化[20]。
1.5 Aux1/LAX候选基因转录组表达数据分析
在NCBI下载毛竹不同生长茎秆的RNA-seq数据(GSE90517,PRJNA604634),包括1.5 cm和8 cm高的毛竹幼苗茎秆、冬笋(15 cm高)、春笋(1.5 m高)、竹鞭、跳鞭。从中提取PheAux1/LAX家族候选基因的FPKM 值(Fragments Per Kilobase of transcript per Million fragments mapped)[21],利用Morpheus进行热图绘制。
1.6 实时定量PCR
根据毛竹不同的发育阶段,选取7个不同高度的竹笋笋尖代表不同的竹笋生长发育时期,分别为冬笋,50、100、300、600、900、1 200 cm高的春笋,命名为S1,S2,S3,S4,S5,S6和S7。采集时间从2020年1月15日(冬笋)开始直至笋快速生长基本停止。取得的笋尖部分在剖去箨壳,液氮速冻后,置于-80 ℃冰箱中保存。利用TRizol法提取不同笋组织的mRNA,采用Prime Script TM RT Reagent Kit (TaKaRa,Japan)反转录试剂盒,将mRNA样品反转录成cDNA。参照SYBR® Green Realtime PCR Master Mix (Roche,Mannheim,Germany)试剂盒的说明书对基因在不同笋组织中的表达展开分析[22]。具体反应体系为:Water,PCR-grade,7.2 μL;Primer F,0.4 μL;Primer R,0.4 μL;SYBR Master Mix,10 μL;cDNA(20 ng·μL-1),2 μL。PCR程序设置为:预热温度设置成95 ℃,5 min。扩增一共设置45个循环,具体步骤为95 ℃,10 s;55 ℃,10 s;72 ℃,20 s。qRT-PCR引物见表1。

表1 qRT-PCR引物Table 1 The primers used for qRT-PCR analysis
2 结果与分析
2.1 毛竹AUX1/LAX候选基因的鉴定及理化特征
根据拟南芥和水稻AUX1/LAX的氨基酸序列在毛竹基因组数据库中进行BLAST搜索、筛选和验证,共鉴定到8个毛竹AUX1/LAX候选基因(表2)。在PheAUX1/LAX候选基因编码的蛋白质中,氨基酸的数目476~534个。分子量为53 155.22~59 132.68 u,理论等电点变化范围为8.27~9.08。亚细胞定位预测除PH02Gene05025.t1定位在细胞膜、液泡;PH02Gene07716.t1定位在细胞膜、细胞核、内质网以外,其余6个PheAUX1/LAX候选成员均定位在细胞膜、液泡和内质网上。
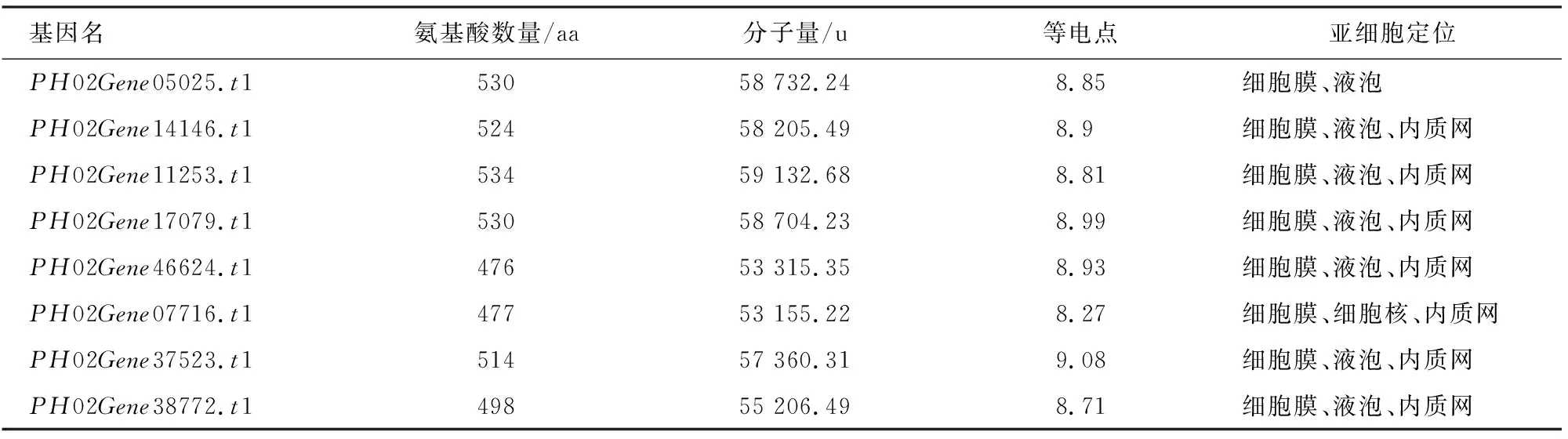
表2 PheAUX1/LAX候选基因的理化性质Table 2 Physicochemical properties of putative PheAUX1/LAX gene family
2.2 PheAUX1/LAX候选基因的系统进化及基因序列分析
使用IQ-Tree软件对拟南芥、水稻和毛竹AUX1/LAX氨基酸序列构建系统进化树(图1a)。根据系统发育树,AUX1/LAX候选基因分为2个亚组:ClassⅠ和ClassⅡ。其中ClassⅠ包括2 个拟南芥、3个水稻、5个候选毛竹成员;ClassⅡ包括2个拟南芥、3个水稻、3个候选毛竹成员。
基序分析表明,候选AUX1/LAX的氨基酸序列在物种内和物种间均非常保守,通过MEME软件分析发现所有水稻、拟南芥、毛竹候选AUX1/LAX的成员均包含6个保守基序。基因结构分析表明,classⅠ的基因包含6~7个外显子,而ClassⅡ的基因包含8~10个外显子(图2)。
2.3 毛竹AUX1/LAX候选基因的染色体定位和共线性分析
染色体定位发现,5、7、14、15、16、21号染色体上分别有1个AUX1/LAX基因,13号染色体上有2个AUX1/LAX候选基因。共线性分析发现,有3对AUX1/LAX候选基因位于共线性区段,对这3对基因所在的染色体进行共线性分析发现,3对染色体均存在大量的共线性区域,表明毛竹进化史上的染色体复制事件(7~12百万年前)在候选PheAUX1/LAX基因家族成员扩增中发挥着重要的作用(图3)。
2.4 PheAUX1/LAX候选基因在不同组织中的表达模式
为了探讨PheAUX1/LAX候选基因在不同组织中的表达模式,基于先前发表的转录组数据对8个基因在1.5 cm高幼茎、8 cm高幼茎、冬笋、春笋、跳鞭、竹鞭、鞭芽中的表达量进行了分析,发现毛竹PheAUX1/LAX候选基因在不同的组织或器官中的表达模式有明显的差异(图4)。其中,4个PheAUX1/LAX候选基因在冬笋中表达丰度最高,且这4个基因均属于classⅠ。PH02Gene46624.t1在1.5 cm高幼茎中表达丰度最高,PH02Gene38722.t1在跳鞭中表达丰度较高,而PH02Gene37523.t1和PH02Gene07716.t1在7个样品组织中均呈现出较低的表达丰度。
8个PheAUX1/LAX候选基因中有4个在冬笋中表达丰度最高,表明PheAUX1/LAX在笋生长发育过程中发挥着重要作用,因此,通过qRT-PCR技术对8个PheAUX1/LAX候选基因在不同生长发育时期笋中的表达量进行分析(图5),发现除了PH02Gene07716.t1和PH02Gene37523.t1以外的其余6个基因与成熟茎秆(CK)相比,在不同笋生长时期均呈现出不同程度的上调表达,而PH02Gene07716.t1和PH02Gene37523.t1均属于ClassⅡ成员。PH02Gene05025.t1、PH02Gene14146.t1、PH02Gene11253.t1和PH02Gene17079.t1随着笋高度增加呈现出逐渐降低的表达趋势。
3 结论与讨论
3.1 结论
AUX1/LAX基因作为生长素输入载体广泛参与植物生长的生长发育,而目前尚未有关于AUX1/LAX在毛竹中的研究报道。分析表明毛竹PheAUX1/LAX候选基因分为Class Ⅰ和Class Ⅱ 2个亚组,其中class Ⅰ的候选基因包含6~7个外显子,而ClassⅡ的候选基因包含8~10个外显子。表达量分析表明多数Class Ⅰ成员在毛竹冬笋中高丰度表达,而Class Ⅱ中的PH02Gene38722.t在跳鞭中高丰度表达。通过qRT-PCR对AUX1/LAX候选基因在不同生长发育时期笋中的表达量分析表明,有6个基因与对照相比呈现出不同程度的上调表达,表明PheAUX1/LAX候选基因广泛参与毛竹笋的生长发育。
3.2 讨论
对不同物种AUX1/LAX的数量统计发现,在拟南芥中共有4个AUX1/LAX成员[23],在水稻中共有6个AUX1/LAX成员24],而在毛竹中共鉴定出8个AUX1/LAX候选成员,多于水稻和拟南芥中的数量。共线性分析发现,有3对AUX1/LAX候选基因位于共线性区段,对这3对基因所在的染色体进行共线性分析,发现每对染色体之间都有大量的共线性区域存在,表明毛竹进化史上的染色体复制事件(7~12 mya)在PheAUX1/LAX候选基因家族成员扩增中发挥着重要的作用。
在毛竹笋生长发育过程中,有6个AUX/LAX候选基因在所有的7个生长发育时期呈现出不同程度的上调表达。有研究表明,AtAUX1主要在拟南芥根尖和侧根根冠原生木质部中表达并在生长素运输中发挥着重要功能[23];AtLAX3能增强细胞壁重塑酶活性,从而增强根原基细胞的分裂和分化能力[25];杨树PtAUX1基因的超表达会改变植株再生芽分生组织中IAA的流向并影响植株对光照的敏感性[26-27];将樱桃PaLAX1转入拟南芥和水稻发现过表达株系中内源IAA的含量增加[28]。而在本研究中,Class Ⅰ基因中的PH02Gene14146.t1、PH02Gene05025.t1、PH02Gene11253.t1、PH02Gene17079.t1在毛竹冬笋中的表达量是最高。qRT-PCR分析发现,8个AUX1/LAX候选基因中的6个在毛竹快速生长过程中呈现不同程度的上调
表达,并多数随着笋的生长发育呈现出逐渐降低的表达趋势。之前的研究表明,笋组织的细胞分裂和细胞伸长生长共同促进了笋的快速生长,其中在快速生长前期以细胞分裂为主,后期以细胞伸长生长为主[29]。而AUX1/LAX候选基因在冬笋期和快速生长前期的高丰度表达,表明这些基因在竹笋分生组织中通过增强细胞壁重塑酶活性,从而增强笋分生组织细胞的分化和分裂能力。本研究将为毛竹AUX1/LAX基因家族各基因的具体功能研究提供一定的理论依据和数据支撑。
