NDRG2通过调控肝癌细胞磷脂和甘油三酯代谢抑制肝细胞癌的生长:基于代谢组学分析
2022-02-03王佳媛袁依依吴有盛田红英
王佳媛,袁依依,张 坤,孙 翔,卜 歆,董 健,吴有盛,田红英,沈 岚
1延安大学医学院病原生物学教研室,陕西 延安 716000;空军军医大学2基础医学院生物化学与分子生物学教研室,3口腔医学院,陕西 西安 710032
肝细胞癌(HCC)是我国最常见的恶性肿瘤之一,临床上该肿瘤具有预后较差和病死率较高的特点。目前,肝癌的治疗方式众多,诸如外科手术切除、放疗、化疗和分子靶向治疗等治疗手段都应用于肝癌的治疗。但是,肝细胞癌患者5年生存率依旧非常低,其治疗效果仍然差强人意[1]。据世界卫生组织国际癌症研究机构数据显示,2020年我国肝癌发病例数41万例,死亡病例数39万例,死亡率极高[2]。因此,肝癌的分子靶向治疗药物逐渐成为目前的研发热点,寻找更加有效且稳定的分子靶向药物是肝癌生物治疗领域的重要内容。目前,临床常见的肝癌分子靶向药物:表皮生长因子受体EGFR抑制剂、血管内皮生长因子受体VEGFR抑制剂、PI3K/Akt/mTOR信号通路抑制剂等,都与肿瘤细胞代谢密切相关[3]。从肿瘤代谢重编程的角度探索新的肝癌分子靶向治疗药物是肝癌生物治疗领域的主要研究内容之一。
肝脏是人体最大的化工厂,细胞代谢十分旺盛。肝癌细胞也具有很显著的代谢重编程特点,即:葡萄糖有氧酵解、磷酸戊糖代谢亢进、脂肪酸合成亢进、谷氨酰胺水解代谢亢进等特点[4,5]。肿瘤细胞通过代谢重编程以满足其迅速增殖时对生物能量和生物原材料的迫切需求[6]。肿瘤代谢重编程的发生与癌基因的活化和抑癌基因的失活密切相关。其中,肿瘤细胞中重要原癌基因Myc的活化、抑癌基因p53的失活都将促进肿瘤代谢重编程[7,8]。Myc可以直接或者间接调节多个糖酵解代谢的关键酶,促进肿瘤有氧糖酵解[9];Myc与转录因子固醇调节元件结合蛋白(SREBP1)调节脂肪生成,促进肿瘤发生[10];Myc还可以通过抑制miR-23a/b促进谷氨酰胺酶表达,增强谷氨酰胺分解代谢[11]。p53转录激活多个与细胞呼吸代谢相关靶基因,参与调节肿瘤细胞糖、脂、氨基酸代谢,抑制肿瘤代谢重编程,从而抑制肿瘤的恶性增殖[12]。
NDRG2是本实验室克隆和发现的新的肿瘤抑制基因。由于该基因在结构上与已发现的N-myc下游调节基因NDRG1具有较高的同源性,故命名为NDRG2[13-15]。该基因在包括肝细胞癌在内的多种肿瘤组织和细胞中表达降低,恢复该抑癌基因的表达可以抑制肝癌细胞生长增殖、有氧酵解、侵袭转移等多种肿瘤恶性表型[16,17]。同时,该基因的表达产物还可以协同mTOR抑制剂参与抑制肿瘤的有氧酵解代谢,增强分子靶向药物依维莫司的治疗效果[18]。因此,深入研究该肿瘤抑制基因在肿瘤脂代谢重编程中的功能,以及该基因在肿瘤分子靶向治疗中的应用价值,具有潜在的临床意义。在本项研究中,我们恢复肝细胞癌中NDRG2的表达,观察其对肝细胞癌脂质代谢的调控效应。
1 材料和方法
1.1 研究资料
1.1.1 主要仪器与试剂 Triple TOF 6500+质谱仪(AB SCIEX),LC-20A超高压液相色谱仪(Shimadzu),色谱柱:Waters,ACQUITYUPLC CSH C18,1.7µm,2.1 mm×100 mm column,低温高速离心机(Eppendorf 5430R),MTBE(霍尼韦尔),超净工作台(苏州净化设备仪器厂),二氧化碳孵箱(Heraeus)。甲醇、乙腈(HPLC级,Merck)。pLenti6-POZ 质粒、pLenti6-NDRG2 质粒、psPAX2 质粒、pMD2.G 质粒为本室保存,转染试剂Lipofectamine 2000(Invitrogen),杀稻瘟菌素(Gibco)。磷脂酶联免疫分析试剂盒(上海通蔚生物科技公司),油红O染色及过碘酸-雪夫PAS染色试剂盒(上海碧云天生物技术公司)。
1.1.2 数据收集 在TNMplot网站(https://tnmplot.com/analysis/)获得抑癌基因NDRG2在肝细胞癌组织与正常肝组织表达丰度的差异[19]。从THPA网站(https://www.pro-teinatlas.org/)获得抑癌基因NDRG2在肝细胞癌患者肝癌组织的表达水平与患者生存期的相关性数据,并通过该数据库分析NDRG2在多种肿瘤细胞株的表达丰度。
1.2 研究方法
1.2.1 细胞培养 人肝癌细胞系HepG2和HEK293T细胞购自购自中国科学院典型培养物保藏委员会细胞库。细胞培养于含10%FBS的完全培养基,在37 ℃、5%CO2、完全饱和湿度条件下常规培养,培养过程中视细胞的生长状态进行换液,并在细胞达到对数生长期、融合度为85%~95%时进行消化传代。消化过程中,先将原培养基吸除,用PBS洗涤3次去除残余培养基,然后加入适量含0.25%胰蛋白酶/EDTA对细胞进行消化,待细胞消化完全后,加入完全培养基终止消化,用移液器轻轻吹打均匀,将细胞悬液置于离心管中,1000 r/min离心5 min,弃上清后加入完全培养基重悬,按照一定比例进行传代,或者计数后进行铺板。
1.2.2 慢病毒包装 接种HEK293T细胞于100 mm细胞培养皿中,培养过夜,待密度达到80%左右进行转染。包装体系包括:10 μg 载体质粒(pLenti-POZ 质粒、pLenti-NDRG2质粒),7.5 μg包装质粒(psPAX2质粒),2.5 μg 包膜质粒(pMD2.G 质粒)以及40 μL 转染试剂Lipofectamine 2000。将上述体系混匀,按照转染试剂说明书进行转染。转染结束后将细胞培养皿放入5%CO2培养箱继续培养,24 h后更换新的培养液,48 h后用0.45 μm滤器过滤收集含有病毒颗粒的培养液。标记并保存于-80 ℃冰箱,用于感染细胞。
1.2.3 慢病毒感染 进行感染前1 d,将HepG2细胞以低密度接种于6孔细胞培养板中,分别置于含5%胎牛血清的McCoy's5A,DMEM培养基中培养。将收集的病毒液融化离心后和新鲜的细胞培养液按1∶1混合加入培养板中。慢病毒组分为Lenti-POZ,Lenti-NDRG2两组,其中Lenti-POZ,Lenti-NDRG2慢病毒带有杀稻瘟菌素抗性基因。病毒感染24 h后,分别更换含有杀稻瘟菌素的新鲜培养液,隔天换液,经过2周左右的压力筛选获得稳定感染的细胞株。
1.2.4 细胞系鉴定与筛选 两组慢病毒Lenti-POZ,Lenti-NDRG分别感染后HepG2细胞后,通过实时定量PCR实验检测分析NDRG2 mRNA含量,每单组样本设3复孔。TRIzol 法提取各组HepG2细胞总RNA,用微量分光光度计检测总RNA的纯度和浓度,并用反转录试剂盒反转录为cDNA,反应条件为37 ℃15 min,85 ℃5 s,4 ℃1 min。采用TB Green Premix II预混液于荧光定量PCR 仪上进行测定,反应条件为95 ℃30 s;95 ℃5 s,60 ℃30 s,40个循环。β-actin基因作为内参,mRNA相对含量通过公式2-△△Ct计算。所有qRT-PCR引物由北京擎科生物科技有限公司设计并合成。其中实时定量PCR实验使用引物序列如下:NDRG2正向引物序列为5'-CAGGACAAACACCCGAGA-3',反向引物序列为5'-AGCCATAAGGTGTCTCCACAG-3'。β-actin 正向引物序列为5'-CGCGAGAAGATGACCCAGAT-3';反向引物序列为5'-GTACGGCCAGAGGCGTACAG-3';同时,通过Western blot实验检测分析NDRG2 蛋白含量。使用RIPA裂解液与PMSF蛋白酶抑制剂的混合溶液从各组HepG2细胞中提取总蛋白质,BCA法测定蛋白浓度,并用5×上样缓冲液配平,置于100 ℃充分变性10 min,蛋白样品上样后,经电泳、转膜至PVDF 膜上;5%脱脂奶粉封闭1 h;4℃冷藏室摇床过夜孵育相应一抗(NDRG2 抗体稀释比例为1∶1000);TBST 洗膜3次,10 min/次;室温摇床上孵育二抗1 h(β-actin抗体稀释比例1∶2000);TBST洗膜3次,10 min/次;超敏ECL试剂显影。
1.2.5 样本处理 收集培养好的各组细胞去除完全培养基,使用4 ℃预冷的PBS 洗涤2 遍,然后加入预冷的生理盐水1 mL 制备细胞悬液并进行细胞计数,确保各样本含有相同的细胞数且不少于1×107/mL。取等量细胞悬液离心,细胞沉淀最后加入1 mL 甲醇:乙腈:水(2∶2∶1,V/V)混合溶液,吹悬细胞置于1.5 mL离心管,保存于-80℃冰箱,用于质谱分析。
1.2.6 LC/MS分析 LC-MS分析使用Triple TOF 6500+质谱仪与LC-20A 超高压液相色谱仪。色谱柱选用Waters ACQUITY CSHC18(2.5 μm,100×2.1 mm)进行分析。流动相:A-0.1%甲酸溶液,B-乙腈(0.1%甲酸);流速:0.4 mL/min;Post Time:5 min;进样量:3 μL。优化的色谱梯度:0~2 min,5%B;2~13 min,5%~95%B;13~15 min,95%B。Post time 设为5 min,用于平衡系统。质谱使用正离子模式结合负离子模式。
1.2.7 信息分析 基于质谱检测得到原始数据文件,首先将原始数据文件导入ScienxOS软件中,进行数据预处理,然后对数据进行质控,保证数据结果的准确度和可靠性对脂质进行多元统计分析,分析方法包括主成分分析(PCA)、偏最小二乘法判别分析(PLSDA)等,用以揭示不同组别脂质的差异。利用层次聚类(HCA)和脂质相关性分析,揭示了脂质和样本之间的关系。通过脂代谢通路等功能分析发现脂质相关的生物学意义。
1.2.8 代谢物含量的检测 分别利用多种磷脂即神经酰胆碱PC、磷脂酰甘油PG、磷脂酰乙醇胺PE、鞘磷脂酰丝氨酸SM、和神经酰胺Cer 5种酶联免疫分析(ELISA)试剂盒检测两组HepG2细胞内相应种类磷脂的含量。每组设立7 个复孔,利用酶标仪检测450 nm 处吸光值A450nm,测定含量并计算相应种类磷脂的含量。
1.2.9 HepG2 细胞油红染色(1)去掉细胞培养基,用PBS洗涤2遍;(2)4%多聚甲醛固定10 min,水洗;(3)加油红O工作液1~2 mL,室温避光染色30 min,密封;(3)60%异丙醇冲洗30 s至背景透明,水洗;(4)苏木素染液复染细胞核30 s;(5)流水冲洗后,甘油明胶封片,室温晾干,于显微镜下观察各组HepG2细胞内脂滴沉积情况并拍照。
1.2.10 HepG2细胞PAS染色(1)去掉细胞培养基,用PBS洗涤2遍;(2)4%多聚甲醛固定10 min,水洗后70%酒精洗涤;(3)0.5%高碘酸酒精溶液氧化10 min后70%酒精洗涤;(4)加入还原液1 min后70%酒精洗涤;(5)加入无色盐基性品红溶液1~1.5 h后流水洗涤10 min;(6)苏木素染液复染细胞核3 min,1%盐酸酒精分化;(7)流水冲洗,脱水,透明,封固后于显微镜下观察各组HepG2细胞内多糖沉积情况并拍照。
1.3 统计学分析
应用SPSS10.0与GraphPad Prism 8软件进行统计分析,对符合正态分布的的计量资料采用t检验或方差分析比较。对于不符合正态分布的计数资料以百分位数法进行比较。P<0.05表示差异有统计学意义。
2 结果
2.1 数据筛选分析
通过TNMplot 数据库获得379 例正常肝组织及809例肝细胞癌组织中NDRG2的表达含量,统计分析后得到正常人肝组织与肝细胞癌患者NDRG2表达差异箱式图,进行对比可发现:相对正常肝组织,肝细胞癌患者的肝癌组织中NDRG2基因表达量明显降低(图1,P<0.05)。
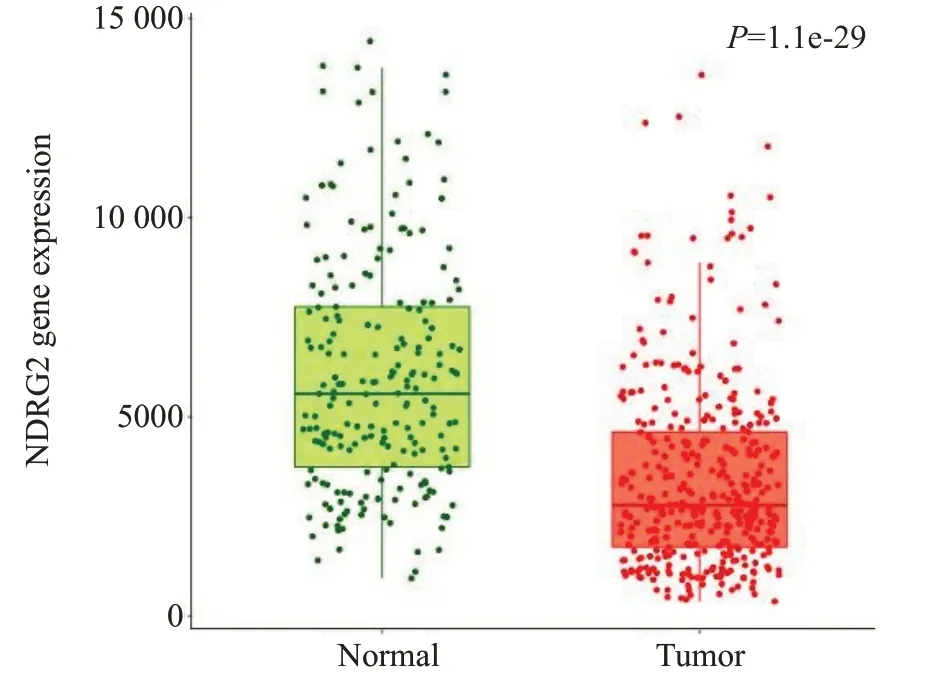
图1 NDRG2基因在正常肝组织及肝细胞癌组织中的表达Fig.1 Expression of NDRG2 gene in normal liver and hepatocellular carcinoma tissues.
2.2 NDRG2表达与HCC预后的关系
通过THPA数据库分析得到NDRG2表达量与肝细胞癌患者预后生存期的关系,结果显示:NDRG2表达量愈高,患者生存时间越长(图2)。在此基础上应用THPA数据库分析NDRG2基因在各种类型肿瘤细胞中RNA的表达丰度,结果显示:肝细胞癌HepG2细胞系中NDRG2基因的表达丰度最高(图3)。
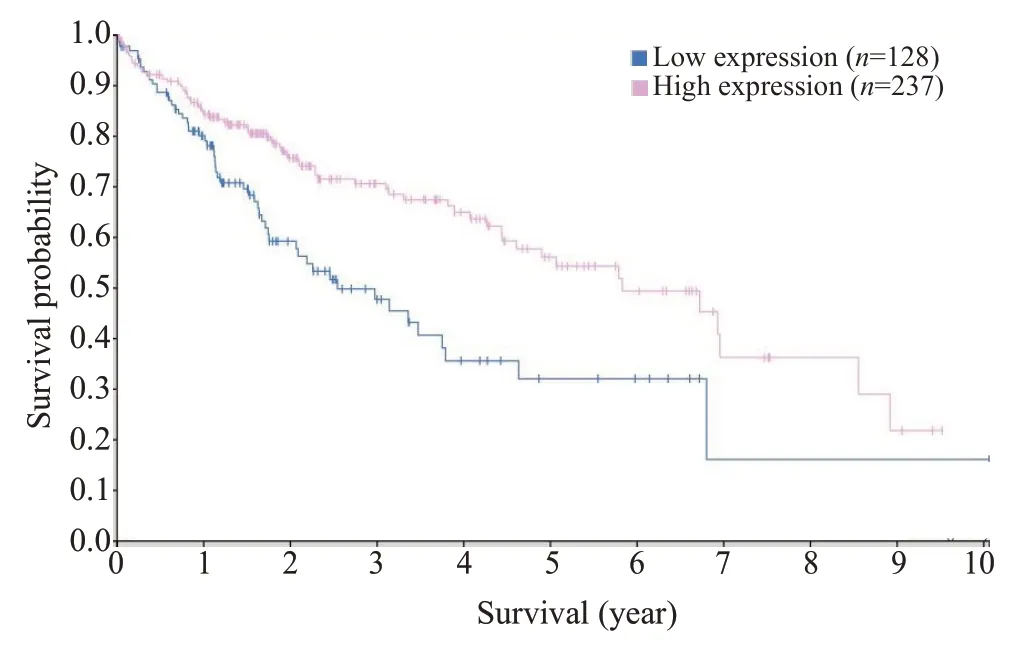
图2 NDRG2表达与HCC病人生存曲线的相关性Fig.2 Correlation between NDRG2 expression and survival of HCC patients.
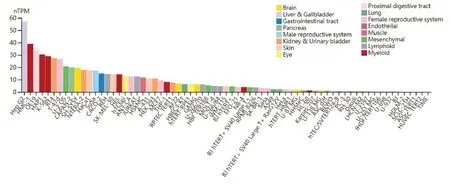
图3 NDRG2基因在各种肿瘤细胞系中的表达丰度Fig.3 Expression of NDRG2 in different tumor cell lines.
2.3 NDRG2在肝癌细胞内的表达含量分析
将携带NDRG2 cDNA及其对照组POZ的慢病毒感染肝癌细胞系HepG2后,通过实时定量PCR实验检测HepG2细胞内NDRG2 mRNA含量,结果显示:与对照组相比,NDRG2的表达明显增强,约为对照组2.6倍(P<0.001,图4A)。同时Western blot实验检测2组细胞内NDRG2蛋白含量,结果显示:与对照组相比,携带NDRG2 cDNA的慢病毒感染后HepG2细胞内NDRG2明显增强(P<0.001,图4B),与实时定量PCR实验结果一致。
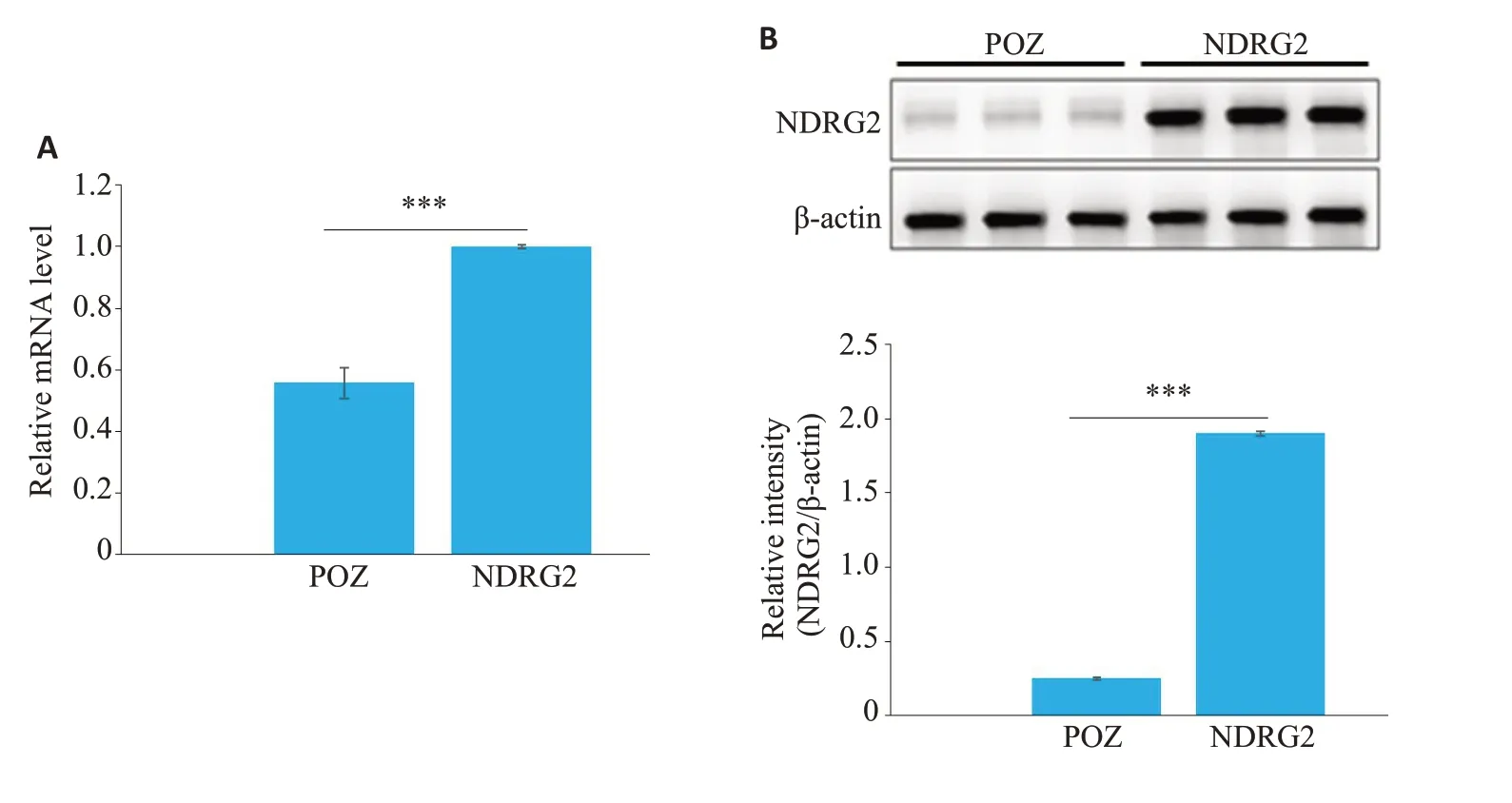
图4 慢病毒感染后HepG2细胞中NDRG2含量Fig.4 NDRG2 expression in HepG2 cells infected with lentivirus containing NDRG2 or control POZ.A: Real-time qPCR for detecting NDRG2 mRNA levels in the infected HepG2 cells.B:Western blotting for detecting NDRG2 protein levels in the infected HepG2 cells.***P<0.001.
2.4 细胞LC/MS分析总样本PCA分析
将QC样本UHPLC-Q-TOF MS总离子流图,进行谱图重叠比较,结果表明各色谱峰的响应强度和保留时间基本重叠,说明在整个实验过程中仪器误差引起的变异较小(图5A)。将所有实验样本和QC样本提取得到的峰,标准化后经Pareto-scaling处理后进行PCA分析(图5B)。观察样本之间的分布趋势,经7-fold crossvalidation(7次循环交互验证)得到PCA的模型参数,找出可能存在的离散点(图5C)。分别建立实验组与对照组的PLS-DA 模型(图5D)和OPLS-DA 模型(图5E),分析NDRG2 过表达的实验组HepG2 细胞与对照组HepG2细胞脂质代谢物的差异。
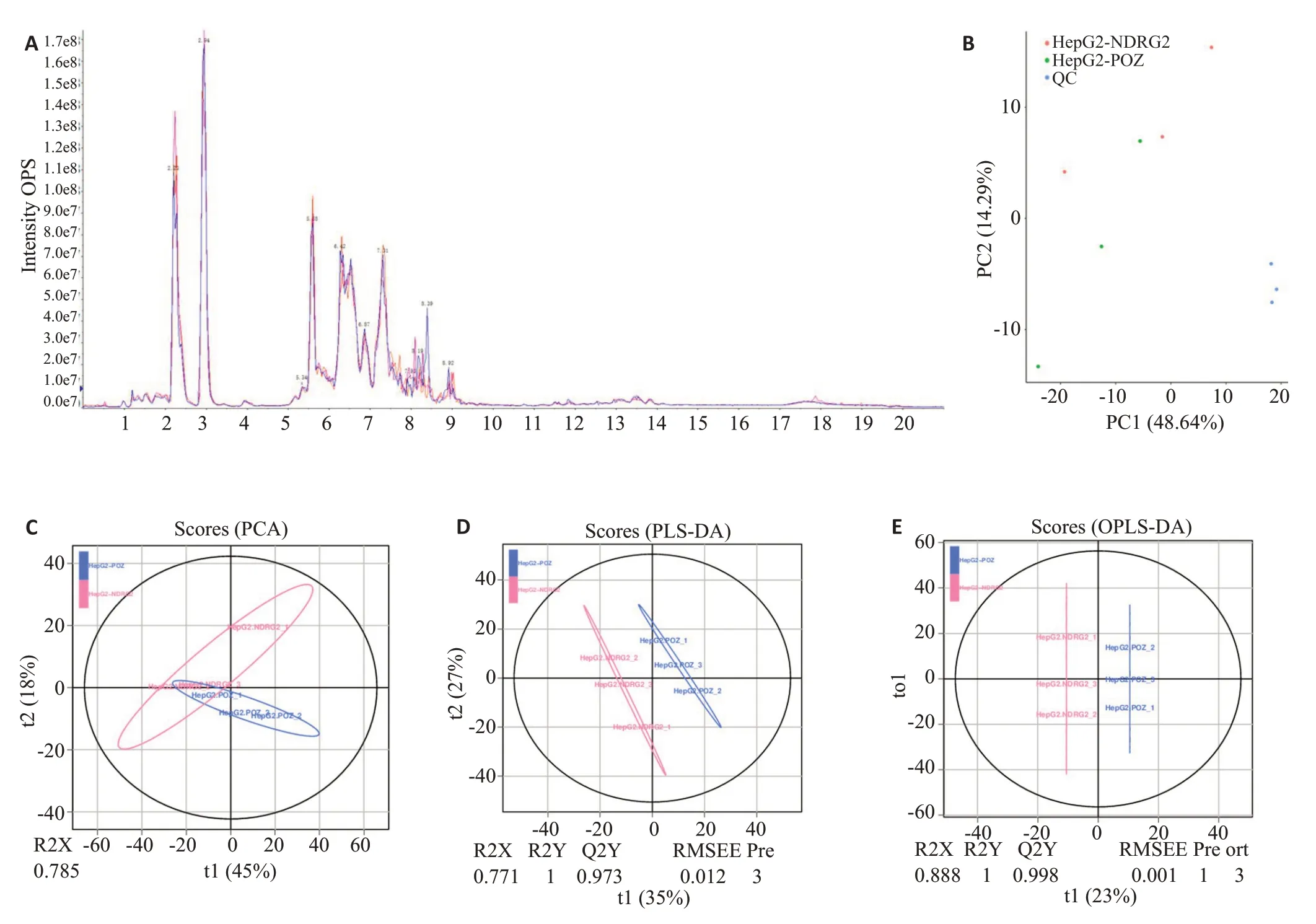
图5 细胞样本的质控分析与代谢轮廓分析Fig.5 Quality control analysis and metabolic profile analysis of HepG2 cells infected with lentivirus containing NDRG2 or control.A:Total ion chromatogram(TIC)of QCs samples.B:PCAanalysis of the total samples.C-E:PCAplots,PLS-DAplots and OPLS-DA plots of NDRG2 overexpression HepG2 cells and control HepG2 cells.
2.5 显著性差异脂质
根据OPLS-DA模型得到的变量权重值(VIP)来衡量各脂质的表达模式对各组样本分类判别的影响强度和解释能力,挖掘具有生物学意义的差异脂质。本实验以VIP>1为筛选标准,初步筛选出各组间的差异物。进一步采用单变量统计分析,验证差异脂质是否具有显著性。选择同时具有多维统计分析VIP>1和单变量统计分析P<0.05的脂质,作为具有显著性差异的脂质;而VIP>1且0.05<P<0.1则作为差异脂质。
从脂质代谢组学的研究结果可以看出NDRG2表达含量增加,导致肝癌细胞中下列脂类物质出现明显变化,其中:溶血磷脂酰胆碱LPC 18:4_1和鞘磷脂SM 40:0出现增加;同时:卵磷脂PC 40:3、磷脂酰甘油PG 32:1、磷脂酰乙醇胺PE 42:2、甘油二酯DAG 38:2_7、磷脂酰乙醇胺PE 44:6、神经酰胺Cer 42:0_2、磷脂酰丝氨酸PS 40:0、和甘油二酯DAG 38:4_7等大量脂类物质出现明显降低(图6)。这些NDRG2表达增强后发生显著变化的脂质代谢物主要是磷脂类代谢物(表1)。
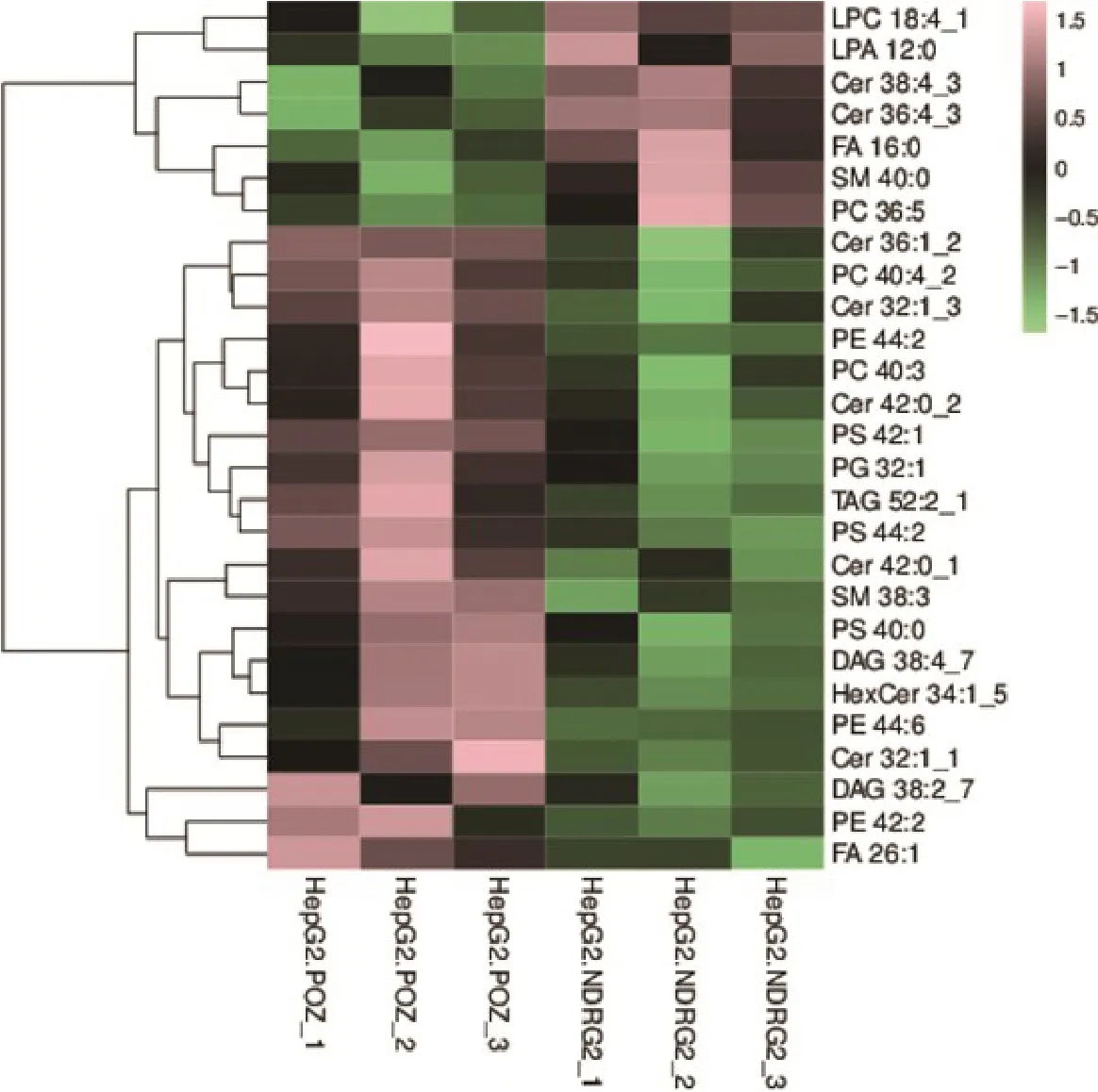
图6 NDRG2过表达与对照组肝癌细胞差异脂质代谢物层次聚类分析Fig.6 Hierarchical clustering analysis of differential metabolites between NDRG2-overexpressing HepG2 cells and control cells.
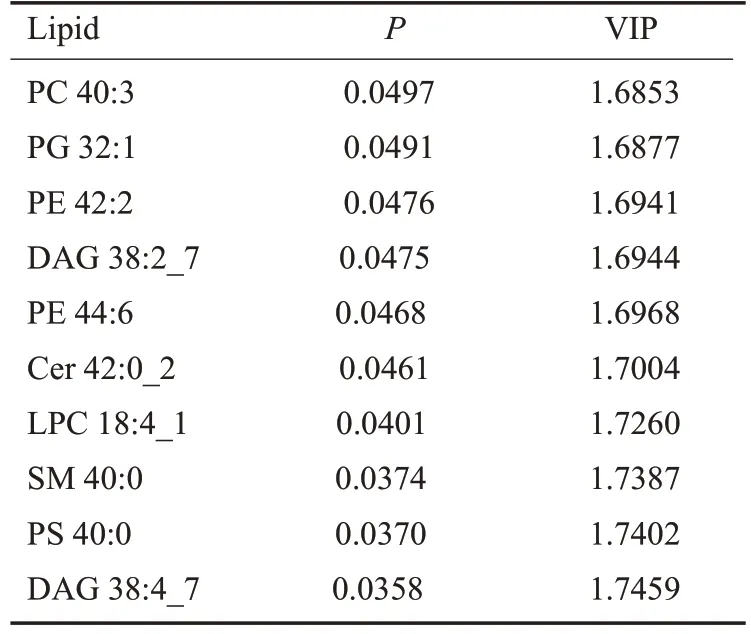
表1 前10种NDRG2表达增强后发生显著变化的脂质代谢物Table 1 Top 10 lipid metabolites that show significant changes in NDRG2-overexpressing HepG2 cells compared with the control cells
2.6 NDRG2过表达对肝癌细胞磷脂含量的影响
ELISA的检测结果显示:与POZ对照组相比,NDRG2过表达的HepG2细胞内磷脂酰丝氨酸PS(155±2.41vs145±2.3,P<0.05)、磷脂酰甘油PG(113±1.73vs106±1.3,P<0.05)、磷脂酰乙醇胺PE(14.55±0.55vs13.11±0.31,P<0.05)、磷脂酰胆碱PC(443.5±1.55vs386.25±0.35,P<0.05)和神经酰胺Cer(261±1.85vs245±2.15,P<0.05)含量减少。NDRG2抑制肝癌细胞内多种磷脂的含量(图7)。
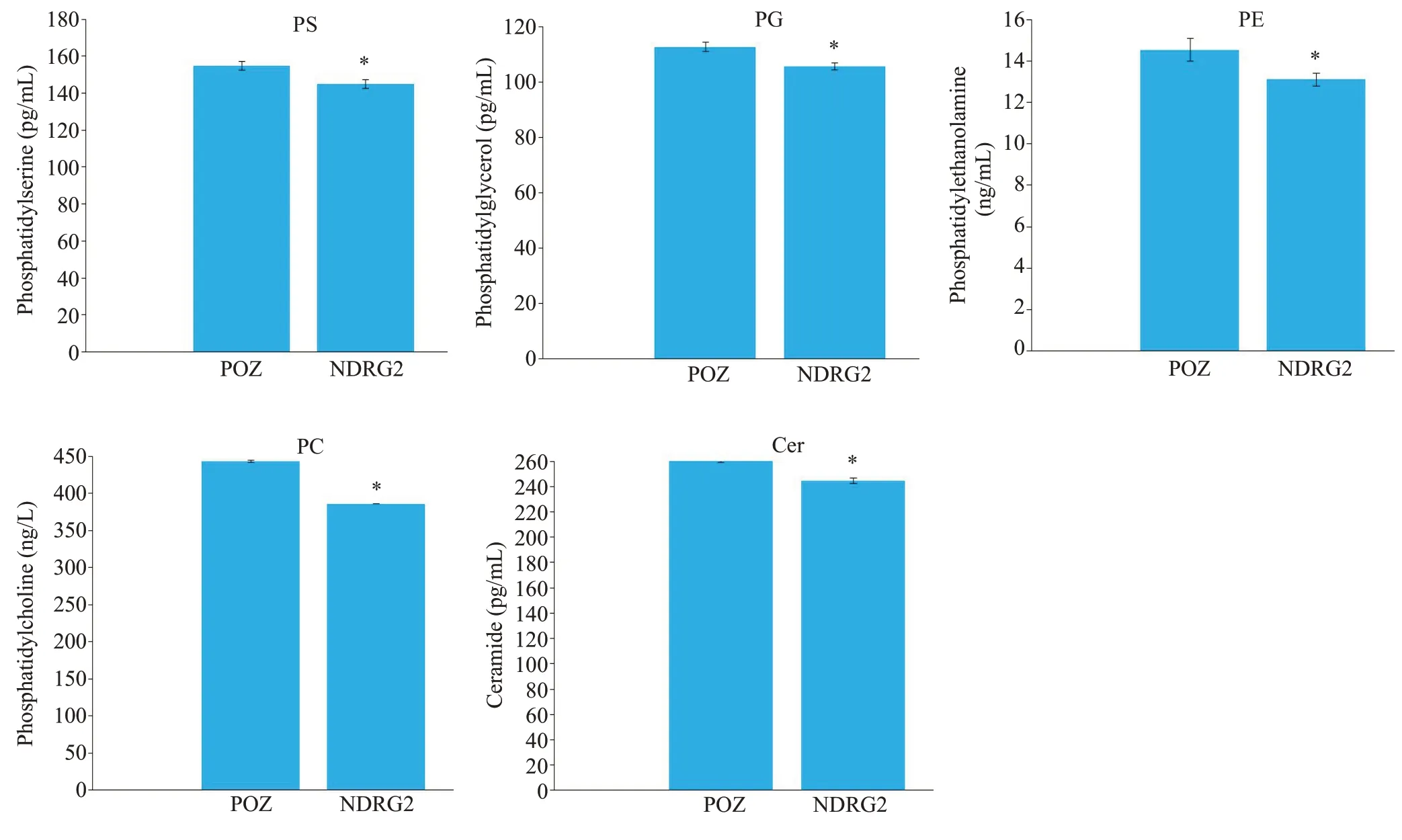
图7 酶联免疫分析NDRG2过表达与对照组肝癌细胞内磷脂代谢物含量Fig.7 ELISAanalysis of phospholipids content in NDRG2-overexpressing HepG2.*P<0.05 vs control.
2.7 NDRG2过表达对肝癌细胞多糖和甘油三酯蓄积的影响
PAS染色显示:与对照组相比,NDRG2过表达的HepG2细胞多糖堆积明显减少,推测糖脂含量减少。油红O 染色显示:与对照组相比,NDRG2 过表达的HepG2细胞脂质堆积明显减少(图8)。NDRG2表达增强后显著抑制了肝癌细胞内甘油三酯和多糖的含量与聚集。
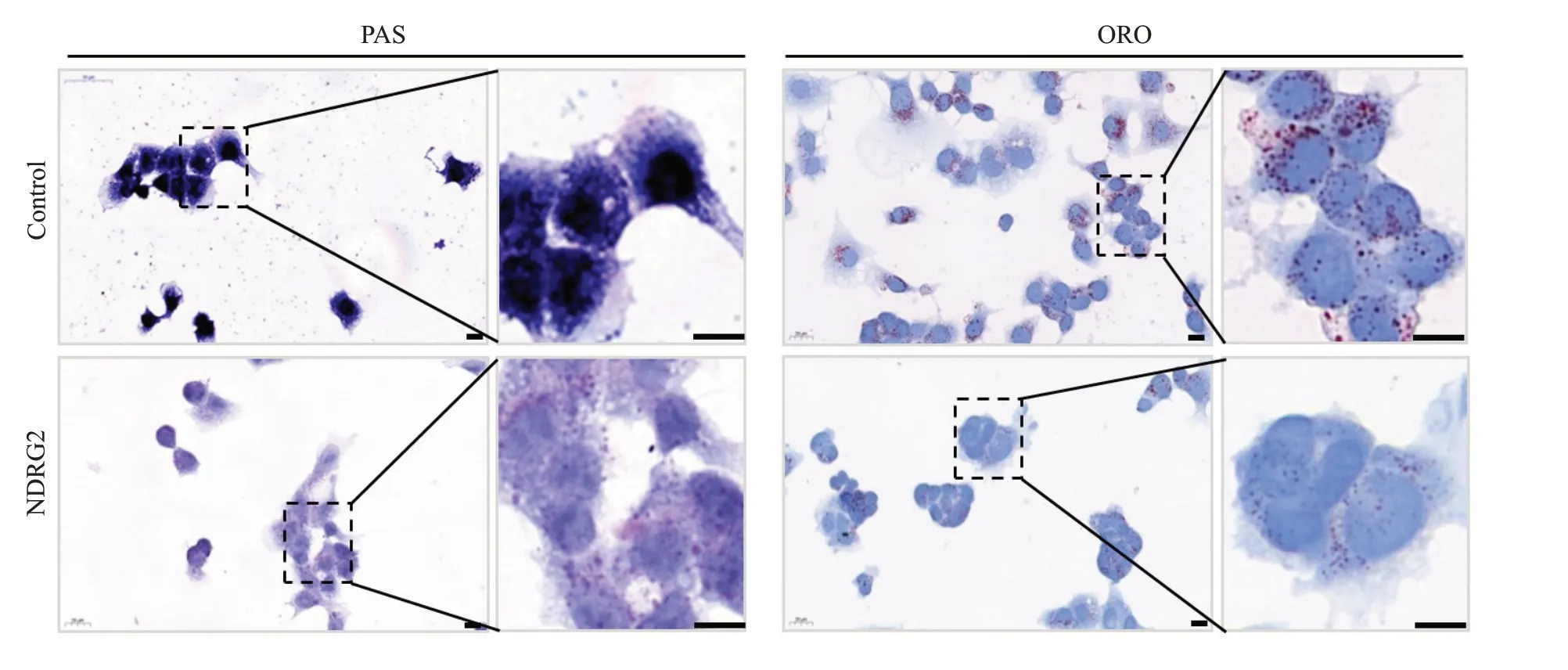
图8 PAS染色和油红O染色分析NDRG2过表达与对照组肝癌细胞内多糖和甘油三酯含量Fig.8 PAS staining and Oil Red O staining for detecting intracellular glycogen and triglyceride in NDRG2-overexpressing HepG2 cells and control cells(Scale bar=50µm).
3 讨论
代谢组学研究目前在基础和临床研究各个领域广泛开展。在肿瘤代谢的研究领域中,代谢组学可以在较广范围内灵敏观察到代谢物的变化,从而聚焦相应代谢途径及其调控模式[20]。本项研究主要关注肿瘤抑制基因NDRG2对肝细胞癌脂质代谢重编程的调控效应。目前,我们的研究结果显示NDRG2主要参与调控肝细胞癌HepG2细胞系中的磷脂和甘油三酯代谢。由于磷脂和甘油三酯合成代谢与游离脂肪酸密切相关,推测NDRG2可能参与脂肪酸的合成代谢过程。后续拟从多株肝癌细胞模型及NDRG2基因敲除小鼠肝脏组织中深入开展表型及机制研究。
磷脂是细胞膜的重要成分,其在细胞物质和能量代谢中发挥重要作用。肝癌细胞磷脂代谢在肿瘤发生早期就出现了异常:在肝细胞癌早期患者的肝组织中多种甘油磷脂含量较高,包括磷脂酰胆碱、磷脂酰乙醇胺以及甘油磷酸胆碱等[21];在肝细胞癌代谢组学研究中常常观察到溶血磷脂酰胆碱的含量有显著变化[22];在包括肝细胞癌在内的多种肿瘤组织中,重要的鞘磷脂分子1-神经鞘氨醇的含量都增加,大量神经鞘氨醇的储存可能与肝细胞癌肿瘤微环境相关[23]。因此,在肝细胞癌发生发展过程中,磷脂代谢重编程既满足了肿瘤生长的能量与物质需求,也为肿瘤生长微环境提供支持。
肝细胞磷脂代谢调控过程中,mTORC2发挥重要功能[24]在肝细胞癌发生发展过程中,mTORC2可以促进心磷脂和鞘脂的生物合成[25]。心磷脂主要位于细胞线粒体中,促进细胞线粒体能量代谢[26]。鞘脂主要参与细胞膜的结构组成,为新生的肿瘤细胞提供原材料[27]。因此,有效抑制mTORC2的活性是肝细胞癌分子靶向治疗的潜在靶点[28,29]。
本项研究中,通过代谢组学的技术方法观察到抑癌基因NDRG2参与调控肝细胞癌的磷脂代谢,其调控磷脂代谢的具体分子机制是什么?其是否影响mTORC2的活性以及如何调控mTORC2的活性都值得进一步分析和探讨。同时,也有研究报道:NDRG2的表达产物可以和PTEN相互作用,从而促进PTEN磷脂酰肌醇磷酸酶的活性,抑制PI3K/Akt信号途径[30]。那么,NDRG2对肝细胞癌磷脂代谢的调控是否依赖于PI3K/Akt/mTOR信号途径也值得进一步研究。
综上所述,本项研究通过生物信息学分析显示肿瘤抑制基因NDRG2在肝细胞癌中具有重要的生物学功能,是肝癌显著的独立预后指标。通过代谢组学和酶联免疫分析证实其参与调控肝细胞癌磷脂代谢重编程。该研究不仅为肝细胞癌磷脂代谢重编程的分子机制研究提供潜在靶点,而且为肝细胞癌的分子靶向治疗提供新的思路。
