转胶蛋白沉默影响三阴性乳腺癌细胞凋亡和自噬的作用机制
2022-02-02杨家磊廖芮婷加晓芳杨文理
杨家磊,唐 朝,廖芮婷,加晓芳,何 涛,杨文理
0 引 言
乳腺癌是女性最常见的恶性肿瘤,已成为全球第一大癌症[1]。根据生物学行为、临床病理特征和分子特征的表达不同,乳腺癌可分为多种亚型。其中,三阴性乳腺癌(triple-negative breast cancer, TNBC)是一类雌激素受体(estrogen receptor, ER)、孕激素受体( progesterone receptor, PR)、和人表皮生长因子受体2( human epidermal growth factor receptor, HER-2)表达均为阴性的乳腺癌[2]。TNBC的恶性程度高、侵袭性强、复发转移率高、死亡率高。因此,寻找三阴性乳腺癌发生发展的相关基因,并阐明其可能的分子机制,对三阴性乳腺癌的防治具有重要意义。
转胶蛋白(transgelin, TAGLN)是钙调节蛋白家族成员之一,由单一N-末端钙蛋白同源结构域(CH)和C-末端钙调蛋白样模序(CLIK)组成[3]。TAGLN在除骨骼肌、红细胞和神经元外的所有细胞和组织中都表达,在平滑肌细胞和成纤维细胞中含量最高。TAGLN与肌动蛋白结合参与细胞骨架相关的细胞增殖、分化、上皮间质转换(epithelial-mesenchymal transition, EMT)等生命活动,与肿瘤的发生发展密切相关。研究表明,TAGLN在不同类型的癌症中的表达不同且作用是相互矛盾的,在乳腺癌[4]、结直肠癌[5]、食管鳞癌[6]和前列腺癌[7]中表达降低,而在晚期结直肠癌[8]、肺癌[9]、胃癌[10]和胰腺癌[11]中表达增强。肿瘤的发生发展是一个多因素、多基因参与的复杂过程,其中细胞凋亡及自噬在这一过程发挥着重要作用[12]。ERK1/2信号通路作为肿瘤研究中的经典信号通路在调节细胞增殖、迁移、凋亡和自噬等多种生物学功能中发挥重要作用[13]。
因此,本研究首先构建TAGLN慢病毒干扰质粒,用慢病毒感染三阴性乳腺癌细胞Hs578T,建立TAGLN敲低的Hs578T sh1、Hs578T sh2稳定细胞系和对照Hs578TCtrl稳定细胞系;然后,分析TAGLN对三阴性乳腺癌细胞凋亡、自噬的影响及作用机制,为三阴性乳腺癌的防治提供新的实验依据。
1 材料与方法
1.1 实验材料三阴性乳腺癌细胞Hs578T、人胚肾HEK293T细胞、大肠杆菌DH5α感受态细胞、shRNA慢病毒质粒pLKO.1-puro为本实验室保存。抗体:TAGLN(Abcam,ab14106)购自成都万道生物科技发展公司,GAPDH(CST, 5174S)、ERK1/2(CST, 4695S)、p-ERK1/2(CST, 4370S)、PARP(CST, 9532S),Caspase3(CST, 9662S),Cleaved-Caspase3(CST, 9664S),LC3A/B(CST, 12741S)购自优宁维生物科技有限公司,Bax(Proteintech, 50599-2-Ig)、Bcl-2(Proteintech, 12789-1-AP)、P62(Proteintech, 18420-1-AP)购自四川生工科技有限公司。DMEM(H)培养基(HyClone)、胎牛血清(FBS)(HyClone)购自成都金线科技有限公司,Western及IP细胞裂解液(凯基生物)购自成都豪乙科技有限公司,PrimeScriptTMRT reagent kit with gDNA Eraser(TaKaRa)购自成都微克生物技术有限公司。
1.2细胞培养Hs578T、HEK293T细胞用含10%FBS的DMEM(H)培养基培养,TAGLN敲低的稳定细胞系(Hs578T sh1、Hs578T sh2)及其对照稳定细胞系(Hs578T Ctrl)在培养时另加1 μg/mL Puromycin,所有细胞常规培养传代。
1.3靶向TAGLN基因shRNA慢病毒载体的构建与鉴定
1.3.1 靶向TAGLN基因shRNA的设计、合成和退火以TAGLN的mRNA序列 ( NM_001001522.2)为目标序列,根据shRNA设计原则,设计、合成2对靶向TAGLN的shRNA干扰序列见表1。靶向Luciferase的阴性对照质粒(ctrl)为实验室保存。将2对寡核苷酸单链片段分别溶于灭菌DEPC H2O,使其浓度为100 pmol/μL。将配对的寡核苷酸片段各取4 μL等量混合,加入8 μL10×M buffer(TaKaRa),总体积16 μL。PCR仪上完成退火程序:96 ℃加热4 min,70 ℃加热10 min,缓慢冷却至4 ℃,-20 ℃保存。

表 1 靶向干扰TAGLN的shRNA序列
1.3.2靶向TAGLN基因shRNA质粒的构建和鉴定将退火后的寡核苷酸双链片段和AgeⅠ/EcoRⅠ酶切、纯化后的pLKO.1-puro shRNA表达质粒按摩尔比3~10:1,取pLKO.1-puro质粒 2 μL(55 ng)、退火寡核苷酸双链片段1 μL、ddH2O 2 μL、TakaRa ligation kit 2.1 solution(Ⅰ) 5 μL,总体积共10 μL。16 ℃连接过夜;再将连接产物转化至大肠杆菌DH5α感受态细胞;涂板、37℃倒置培养过夜;挑取单克隆培养,提取质粒并用AgeⅠ/EcoRⅠ酶切鉴定,体系为:质粒DNA 1 μg, 10×H buffer 2 μL,EcoRⅠ(12 Unit/μL) 0.5 μL,NdeⅠ(12 Unit/μL) 0.5μL加ddH2O 补足20 μL,37 ℃孵育2 h。经1g/L琼脂糖凝胶电泳酶切产物,阴性质粒仅有75 bp插入条带,阳性质粒可见134 bp插入条带,20%甘油保种。阳性克隆菌送生工生物工程(上海)股份有限公司测序,分别将测序正确的克隆命名为sh1和sh2。
1.4确定Hs578T细胞的Puromycin最佳筛选浓度接种2×105个/(0.5 mL·孔)的Hs578T细胞至24孔板,当细胞70%~80%融合时,分别加入不同浓度的嘌呤霉素浓度(0、0.5、1、2、4、6、8 μg/mL);继续培养,每隔3天换液,7 d后,选择Hs578T细胞全部死亡的最低嘌呤霉素浓度(1 μg/mL)作为最佳筛选浓度。
1.5构建TAGLN沉默的Hs578T稳定细胞系大量提取测序正确的2种TAGLN shRNA质粒(sh1和sh2)和阴性对照质粒(ctrl),待HEK293T细胞融合到70%~80%时进行慢病毒包装。根据Lenti-PacTMHIV慢病毒包装试剂盒(GeneCopoeia)说明书,分别制备质粒DNA-转染试剂复合物,更换细胞培养基为无血清无双抗的DMEM(H);将复合物分别滴加入细胞培养基中,8 h后更换新鲜全培养基;再继续培养48 h,分别收集24 h和48 h的细胞培养基(含病毒颗粒),0.45 μm的无菌滤器过滤去除细胞碎片,获得慢病毒悬液,-80 ℃保存备用。待Hs578T细胞融合到70%~80%时,换液,加8 μg/mL的Polybrene,用慢病毒悬液进行感染。24 h后,用 1 μg/mL 的Puromycin对细胞进行筛选,2~4周后获得TAGLN敲低的稳定细胞系(Hs578T sh1、Hs578T sh2以及对照稳定的细胞系(Hs578T Ctlr)。
1.6qRT-PCR和Western blot用Trizol分别提取稳定转染细胞的总RNA,并根据PrimeScriptTMRT reagent kit with gDNA Eraser说明书,去除基因组DNA后进行逆转录。qPCR引物见表2,由生工生物工程(上海)股份有限公司合成;qPCR体系为:TB Green Premix Ex TaqⅡ12.5 μL、PCR Forward Primer(10 mΜ) 1.0 μL、PCR Reverse Primer(10Μm)1.0 μL、RT反应液(cDNA溶液)2 μL、灭菌水、8.5 μL,总体积25 μL;qPCR反应条件为95 ℃ 30sec→95 ℃ 5sec→60 ℃30~60sec,后面两个步骤重复40个循环,采用2-△△CT法进行定量分析。

表 2 qPCR引物序列
按细胞总蛋白提取试剂盒(南京凯基)说明书,分别提取稳定转染细胞的总蛋白,留取小部分进行BCA(碧云天)浓度测定;其余总蛋白按比例加入5×上样缓冲液,沸水浴10 min变性,分装、-80 ℃保存备用;蛋白样品进行12% SDS-PAGE电泳,转膜,5%BSA-0.1%TBST封闭1h;分别加封闭液稀释的一抗TAGLN(1∶500)、GAPDH(CST,1∶1000)、ERK1/2(1∶1000)、p-ERK1/2(1∶1000)、Bax(1∶500)、Bcl2(1∶500)、Caspase3(1∶1000)、Cleaved-Caspase3(1∶500)、LC3I/II(1∶1000)、P62(1∶500)4 ℃、过夜孵育;1×TBST漂洗5 min×3次,室温孵育相应的近红外荧光二抗1 h,1×TBST漂洗5 min×3;Odessey近红外成像系统进行图像采集及灰度分析。
1.7流式细胞术检测细胞的凋亡分别收集TAGLN敲低的稳定细胞系Hs578T sh1、Hs578T sh2及对照稳定细胞系Hs578T Ctrl,各个细胞系分别分组:空白组(不加FITC-Annexin V、PI组)、FITC-Annexin V组(只加FITC-Annexin V)、PI组(只加PI组)、FITC-Annexin V+PI组(加FITC-Annexin V及PI)。按Annexin V-FITC凋亡检测试剂盒(凯基生物)说明书,分别加入100 μL Binding Buffer重悬细胞;每组分别不加或加入5 μL FITC-Annexin V和(或)5 μL 50 μg/mL PI,室温避光孵育15~25 min,再加入400 μL的Binding Buffer终止染色,避光、1 h内完成流式细胞仪上机检测。

2 结 果
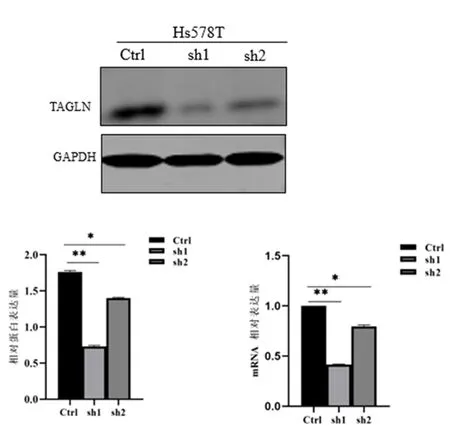
2.1 TAGLN敲低Hs578T稳定细胞系的构建及鉴定NdeⅠ/EcoRⅠ双酶切结果显示,获得插入TAGLN shRNA序列的阳性质粒,见图1。测序结果显示,成功建立了TAGLN慢病毒shRNA质粒(sh1及sh2),见图2。包装慢病毒并感染Hs578T细胞,经1 μg/mL puromycin筛选2~4周后,获得TAGLN敲低的稳定细胞系(Hs578T sh1、Hs578T sh2)及对照细胞系(Hs578T Ctrl)。Western blot和qPCR结果显示,与对照细胞Hs578T Ctrl相比,TAGLN敲低细胞Hs578T sh1、Hs578T sh2的TAGLN蛋白表达分别减少58%、21%(P<0.01)、mRNA表达分别降低32%、48%(P<0.01),见图3。提示TAGLN敲低的三阴性乳腺癌稳定细胞系Hs578T sh1、Hs578T sh2及其对照稳定细胞系Hs578T Ctrl构建成功。
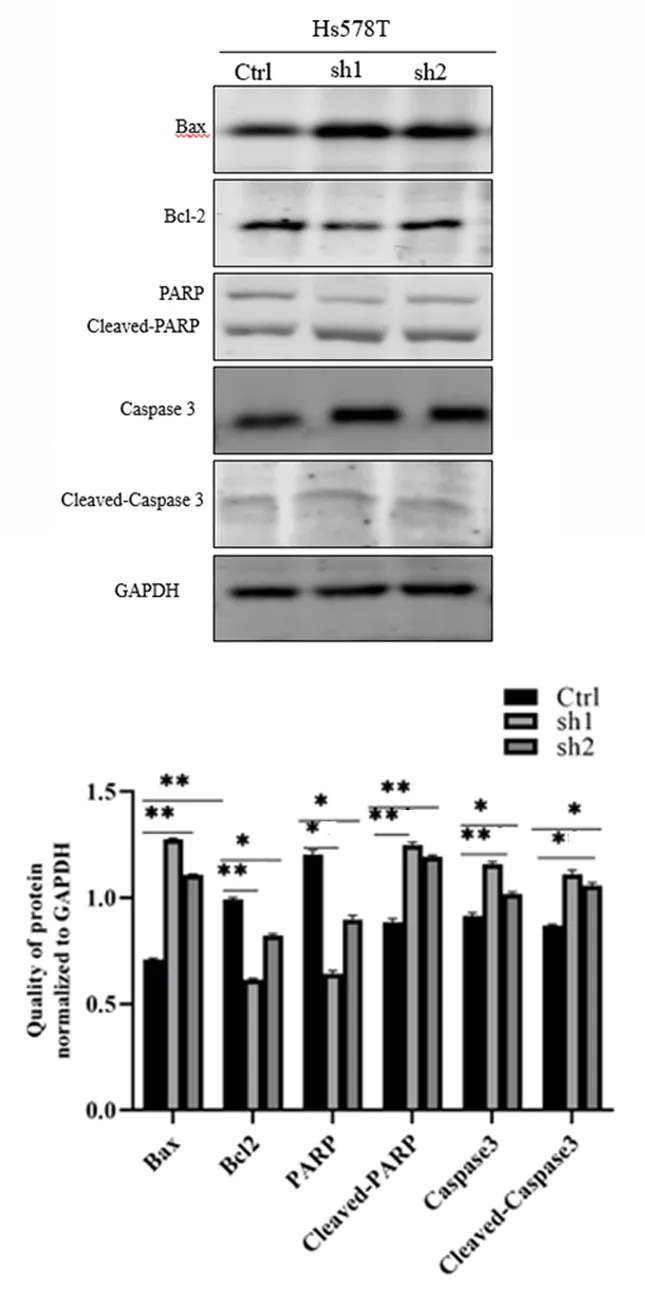
2.2敲低TAGLN促进Hs578T细胞的凋亡流式细胞术检测细胞凋亡结果显示,敲低TAGLN的三阴性乳癌Hs578T sh1、Hs578T sh2稳定细胞较Hs578T Ctrl细胞的总凋亡率分别增加100.7%、74.4%(P<0.01),早期凋亡率分别增加126.4%、85.7%(P<0.01),晚期凋亡率分别增加49.2%、51.9%,见图4,表3。Western blot结果显示,敲低TAGLN的Hs578T sh1、Hs578T sh2稳定细胞较Hs578T Ctrl细胞的抗凋亡蛋白Bcl-2表达均减少,促凋亡蛋白Bax表达均增多,Bax/Bcl-2的分别增加148%、115%,Cleaved-PARP蛋白分别增加41%、34%,Capase3蛋白分别增加26%、11%,Cleaved-Caspase3 蛋白增加27%、21%(P<0.01),见图5。表明敲低TAGLN促进了三阴性乳腺癌Hs578T细胞的凋亡。
2.3敲低TAGLN抑制Hs578T细胞的自噬通量Western blot结果显示,敲低TAGLN的Hs578T sh1、Hs578T sh2稳定细胞较对照Hs578T Ctrl细胞的LC3-Ⅱ蛋白表达减少,而P62蛋白表达升高(P<0.01),说明敲低TAGLN抑制了三阴性乳腺癌Hs578T细胞的自噬通量。见图6。

1、3、5: 分别是ctrl、 sh1、 sh2未进行酶切; 2、4、6: 分别是酶切后的ctrl、sh1、 sh2

a:TAGLN sh1质粒的测序结果;b: TAGLN sh2质粒的测序结果
*P<0.01、**P<0.001
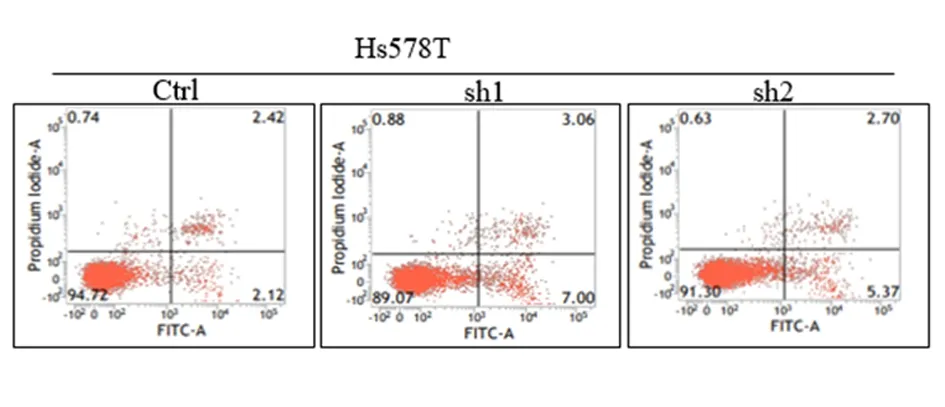
图 4 敲低TAGLN增加Hs578T细胞的凋亡率
表 3 Hs578T Ctrl、Hs578T sh1、Hs578T sh2细胞的凋亡率比较
*P<0.01、**P<0.001
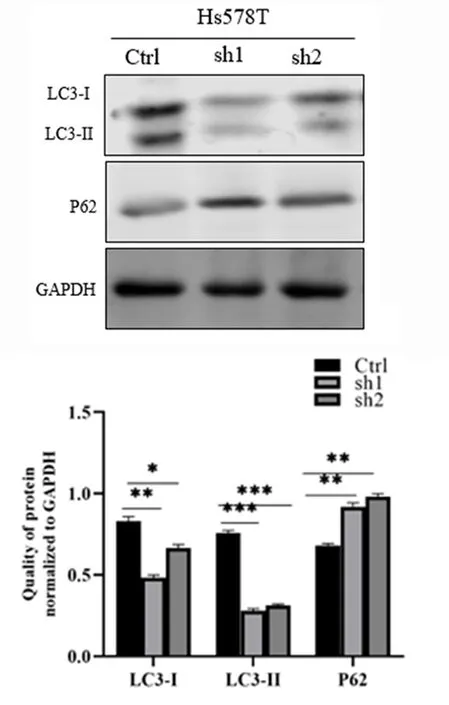
*P<0.05、**P<0.01、***P<0.001
2.4敲低TAGLN通过ERK1/2信号通路影响Hs578T细胞的凋亡和自噬Western blot结果显示,敲低TAGLN的Hs578T sh1、Hs578T sh2稳定细胞较对照组Hs578T Ctrl细胞的ERK1/2、p-ERK1/2蛋白表达减少(P<0.01),提示敲低TAGLN可能通过抑制ERK1/2的磷酸化水平,进而促进三阴性乳腺癌细胞的凋亡及抑制细胞的自噬通量。见图7。
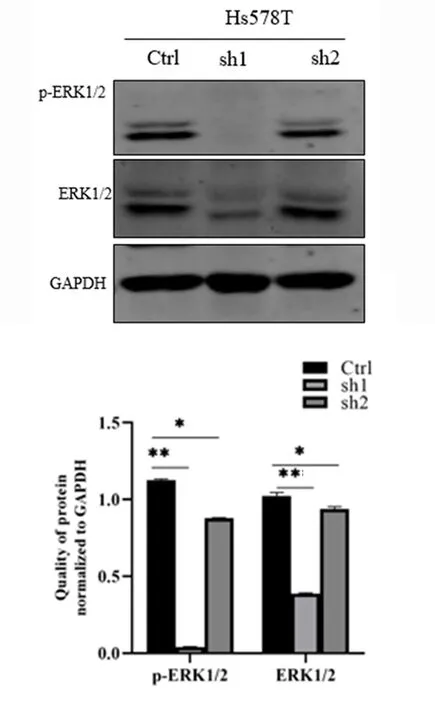
*P<0.01、**P<0.001
3 讨 论
TAGLN是一种细胞骨架相关蛋白,该基因及其蛋白质序列具有种属间的高度同源性及进化保守性。TAGLN主要通过稳定肌动蛋白丝,参与细胞骨架的重塑,进而参与肿瘤的发生发展,但在不同类型肿瘤中的生物学功能又不尽相同甚至相反的。既往研究报道TAGLN基因的缺失对肿瘤早期进展很重要,可作为乳腺癌和结直肠癌的诊断标记物之一;可促进乳腺癌血管正常化并抑制血管出芽和肿瘤转移;过表达TAGLN抑制人乳腺癌细胞系MDA-MB-231的体外迁移和侵袭;TAGLN启动子甲基化下调其表达并与乳腺癌患者无复发生存率呈负相关[4,14-16]。本研究通过慢病毒shRNA沉默TAGLN,建立TAGLN敲低的Hs578T sh1、Hs578T sh2稳定细胞系和对照Hs578T Ctrl稳定细胞系,探讨敲低TAGLN是否影响三阴性乳腺癌Hs578T细胞凋亡及自噬。
细胞凋亡是由多种基因调控的细胞程序性死亡,主要有线粒体途径、死亡受体途径、内质网途径及颗粒酶B途径[17]。在线粒体途径中,Bcl-2家族是最为重要的调节因子之一,包含促凋亡因子(Bax, Bad, Bid)和抗凋亡因子(Bcl-2, Bcl-W, Mcl-1)两类。Bcl-2和Bax形成异源二聚体调节转录活性,通过细胞色素C的释放调节细胞凋亡,Caspase-3在凋亡级联反应的下游发挥重要作用,通过降解细胞内相应底物诱导细胞发生凋亡[18]。本研究中流式细胞术结果显示,与对照Hs578T Ctrl稳定细胞系相比,敲低TAGLN的Hs578T sh1、Hs578T sh2稳定细胞系的凋亡率增加,Bcl-2表达下调,Bax、Cleaved-PARP、Caspase3、Cleaved-Caspase3表达上调,提示敲低TAGLN通过激活Caspase3从而促进了三阴性乳腺癌细胞Hs578T的凋亡。
自噬是一种保守的分解代谢过程,在维持细胞内环境稳态、物质循环再利用等方面发挥重要作用[19]。自噬发生过程中涉及一系列信号转导途径的参与,在机体营养不充足时,mTOR失活、MAPK活化催化ULK1复合物的形成,从而引发自噬[20]。自噬发生后,LC3-Ⅰ在共价连接酶ATG3的作用下转变为膜结合形式的LC3-Ⅱ,参与自噬小体的结合;P62是一种泛素结合的蛋白,在自噬发生时P62可与LC3-Ⅱ蛋白结合形成复合物,然后在自噬溶酶体中降解[21-22]。本研究中,TAGLN敲低Hs578T sh1、Hs578T sh2稳定细胞系中LC3-Ⅱ的表达减少,P62表达增加,表明敲低TAGLN通过减少LC3-Ⅱ而抑制了Hs578T细胞的自噬通量。
丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPK)是细胞内的一类丝氨酸/苏氨酸蛋白激酶,参与细胞生长、分化和程序性死亡等一系列生物学过程。在哺乳动物细胞中,与ERK1/2相关的细胞内信号途径被认为是经典MAPK信号转导途径;ERK1/2参与多种生物学调节过程,包括细胞周期、迁移、凋亡及自噬等。研究表明ERK1/2通过下调Bcl-2、MMP-9等蛋白,上调Bax、Bad等蛋白促进细胞凋亡,而TAGLN过表达导致RAS和ERK1/2的激活增加,其下调也导致激活的RAS和ERK1/2减少[23-24]。在HeLa细胞中,氧化应激诱导剂VK3诱导细胞自噬,激活ERK1/2,ERK1/2抑制剂U0126可以阻断该过程LC3-Ⅱ的表达,说明ERK1/2参与细胞自噬的形成[25]。在本研究中,敲低TAGLN的Hs578T sh1、Hs578T sh2稳定细胞系的ERK1/2、p-ERK1/2明显下调,说明TAGLN以依赖ERK1/2途径促进三阴性乳腺癌细胞Hs578T的凋亡及抑制自噬通量。
综上所述,本研究成功构建了TAGLN低表达及对照的三阴性乳腺癌Hs578T稳定细胞系,TAGLN沉默促进三阴性乳腺癌细胞凋亡并抑制自噬通量,说明TAGLN在三阴性乳腺癌Hs578T细胞中扮演癌基因的角色,可能促进肿瘤的发生发展,为深入研究TAGLN作为三阴性乳腺癌防治的潜在分子靶点提供了实验依据。
