冻融条件下生物结皮覆盖对土壤饱和导水率的影响
2022-01-28曾建辉郭迎香张泽宇李占斌刘晨光
曾建辉,马 波,2,郭迎香,张泽宇,李 桂,李占斌,2,*,刘晨光
1 西北农林科技大学水土保持研究所,黄土高原土壤侵蚀与旱地农业国家重点实验室 杨凌 712100 2 中国科学院水土保持研究所 杨凌 712100
土壤饱和导水率(saturated soil hydraulic conductivity,Ks)是土壤被水饱和时,在单位时间和水势梯度下通过单位面积的水量,它是土壤质地、容重、孔隙分布特征的综合反映,也是重要的土壤水力学参数之一,在进行土壤水分模拟时必须考虑[1]。土壤处于饱和状态时,孔隙全部充满水,此时导水率的数值近似于土壤稳定入渗速率[2]。在水资源匮乏的干旱半干旱地区,饱和导水率影响地表水文变化过程,对水的转化储存至关重要。我国西北地区,由于冬春降雨稀少,气温骤降且地表植被发育不良,生态环境脆弱,受到冻融作用的影响更剧烈,季节性冻土广泛分布。然而季节性冻土的消融是一个漫长的过程,土壤温度的日变化对冻融过程影响显著,对浅层土的作用尤其明显[3 ]。研究表明[4—5],冻融作用会影响土壤温度,改变土壤理化性质以及质地,伴随着三相比例变化,土壤的微观结构在不同冻融次数下也明显不同,而冻融循环过程对表层土壤的影响是最显著的。生物结皮(Biological soil crust,BSC)作为寒区旱区广泛存在的地被物,在生态系统中起着重要的作用,特别是在土壤理化性质和土壤入渗特性方面[6]。然而,冻融作用对结皮土壤导水率和强度的影响与我们预期的冻融循环对无结皮土壤的作用效果不一致[7],其影响取决于冻融循环次数,结皮类型,结皮理化性质等。Assouline[8],Lichner[ 9],Wang[10],以及刘翔等[11]表示BSC能有效减少土壤入渗;而Bu[12]以及Belnap等[13]认为BSC通过粗糙表面的滞流作用和物理结构的改善促进了降雨的入渗,因此目前生物结皮对饱和导水率的作用研究结果仍有分歧。BSC的生长状态表现出很大季节变化,受土壤温度和湿度影响显著[14],而BSC厚度影响土壤中湿润锋深度,是反映土层中水分入渗的重要参数[15]。冻融土壤水分入渗特性还受粒径分布、土壤结构,土壤含水量、入渗水温以及水稳性团聚体、有机质等影响[16]。何志萍等[17]将冻融土壤导水率的降低原因归结于过水断面减少,水通路和水的粘滞性改变以及冰晶透镜体的存在等原因。在冻融条件下,结皮覆盖土壤的饱和导水率影响因素众多,变化趋势复杂且不同于裸土,从生态学角度看,冻融条件下土壤的变化影响土壤节水保肥能力,从而导致水土流失、荒漠化等[4]。目前相关研究多集中于冻融作用对裸土理化性质的影响或是冻融作用对BSC生理生化特征的改变作用,对于冻融条件下结皮土壤水文特征及水分变化研究尚且不足,因此本研究旨在揭示冻融作用对蓝藻结皮土壤饱和导水率的影响机制,以及结皮土壤在不同冻融交替次数和含水量下的变化规律,量化土壤饱和导水率的大小,为春冬季节寒区旱区土壤侵蚀评估和生态治理提供理论依据。
1 研究地区与研究方法
1.1 研究区概况
研究区域位于内蒙古鄂尔多斯市达拉特旗十大孔兑的东柳沟流域,是黄土高原北部典型的风水复合侵蚀区(39°58′2″—40°12′50″N,110°28′47″—110°31′35″E)。平均海拔995—1480m,年平均降雨量为260—390 mm,且多集中在7—8月。年平均气温为6.85℃,冬季平均气温为0℃以下,最低可达-32.5℃,每年十月下旬至次年的四月为封冻期,冻土深度达到157 cm[18]。每逢入春时期,研究区的平均气温在0℃上下波动,且受日照影响昼夜温差可达20℃,伴随融雪侵蚀形成了渗透性极弱的隔水层,使融水更易集流并具有冲刷能力[19]。
供试土壤为典型的风沙土,主要的构成颗粒为细砂,是干旱半干旱地区砂性母质上形成的幼年土。颗粒间非毛管孔隙大量分布,土壤粘结性较差,受冻融作用影响使该地区土壤性质发展不同于其它地区。研究区内地表植被稀疏,自然单元内生物结皮主要以藻结皮和苔藓结皮居多,并伴有少量地衣,生物结皮呈现混合生长状态。藻结皮的优势种为小席藻(Phormidiumtenue)、具鞘微鞘藻(Microcoleusvaginatus)和爪哇伪枝藻(Scytonemajavanicum)。地表植被优势群落包括猪毛菜(SalsolacollinaPall.),小蓬草(Conyzacanadensis),狗尾草(Setariaviridis),艾蒿(ArtemisiaargyiH),马唐(Digitariasanguinalis),少花米口袋(Gueldenstaedtiaverna)以及地锦草(EuphorbiahumifusaWilld)。
1.2 研究方法
1.2.1样品采集
选取研究区域内自然撂荒地上生物结皮发育良好且具有代表性的4—6个退耕地为研究样地进行野外调查,用100 cm3环刀在样区内采集用于测定饱和导水率的藻结皮土壤和裸土样品。采集生物结皮层及结皮层下0—5 cm的土样,分层混合后带回室内用于基本理化性质测定。有机质采用重铬酸钾容量法测定,全氮通过凯氏定氮法测定,粘结力通过土壤剪力测量仪(15.10型袖珍剪切仪CL101)分别测定保留结皮层粘结力和去除生物结皮后下层土壤的粘结力,测量的结果除去最大和最小值后取平均,结果见表1。根据结皮盖度调查结果,样地内藻结皮盖度普遍在40%—68%左右。
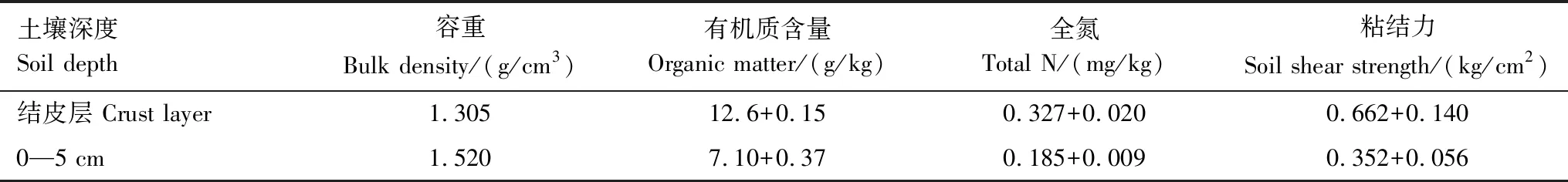
表1 生物结皮土壤样品理化性质
1.2.2试验方法
选取冻融初始含水量、冻融交替次数为试验因子,采用析因实验设计,考虑结皮容重、结皮厚度、粒径分布等土壤物理性质以及有无结皮覆盖对结果的影响。研究区内解冻期降水少而蒸发量大,根据土壤实测含水量,其最低含水量为2.31%,根据相关文献表明,表层5 cm土壤在较长时间尺度上的冻融期内,所达到的最高和最低含水量差异的均值为15%[20],因此,将冻融模拟初始含水量设定为:4%,6%,8%,15%。土壤经过15次冻融交替后,土壤结构性质基本趋于稳定[5],为明确持续冻融作用在结构稳定后对土壤渗透性能的影响,因此将冻融交替次数设定为:0,1,3,5,7,10,15,20次,共进行37组试验,每组重复5次。供试土壤在阴凉处风干后,按照既定水分梯度进行预处理,为了最大程度模拟自然条件下土层温度的传递过程,在环刀外围包裹隔绝温度的气凝胶垫,热传导系数为0.018 w m-1k-1,将处理好的土样用保鲜膜覆盖防止水分蒸发散失,并在室温下静置12 h,待水分分布均匀后放入制冷机中进行冻融交替循环。根据研究区域的气温变化和土层昼融夜冻(昼夜各12 h)的自然状态将冻结温度设定为-10—-15℃,并在10—15℃的室温条件下进行融解,冷冻和解冻时长各12 h。
在冻融交替结束后测定土壤的湿重和饱和导水率,再从试验样品中采样测定结皮厚度,结皮层容重,结皮下层土壤容重以及对土壤颗粒组成进行分析。结皮厚度用游标卡尺测量,结皮容重采用涂膜法[21],结皮下层土壤容重采用环刀法测定。通过Mastersizer 2000型激光粒度分析仪进行土壤颗粒分析,土壤质地的划分依照美国制土壤分级标准,通过恒定水头法测定土壤的饱和导水率。
1.3 数据处理
土壤结构指数可以用来定量反映土壤受到外力作用而引起土壤结构发生变化的程度,而土壤结构对土壤饱和导水率具有重要影响[22]。根据土壤颗粒分析、土壤容重和含水率分析的结果得到土壤三相比,计算广义土壤结构指数[23]:
GSSI=[(Xs-25)XLXG]0.4769
式中,Xs为土壤固相体积百分比,Xs> 25;XL为土壤液相体积百分比,XG为土壤气相体积百分比。
土壤颗粒体积分形维数计算公式为[24]:
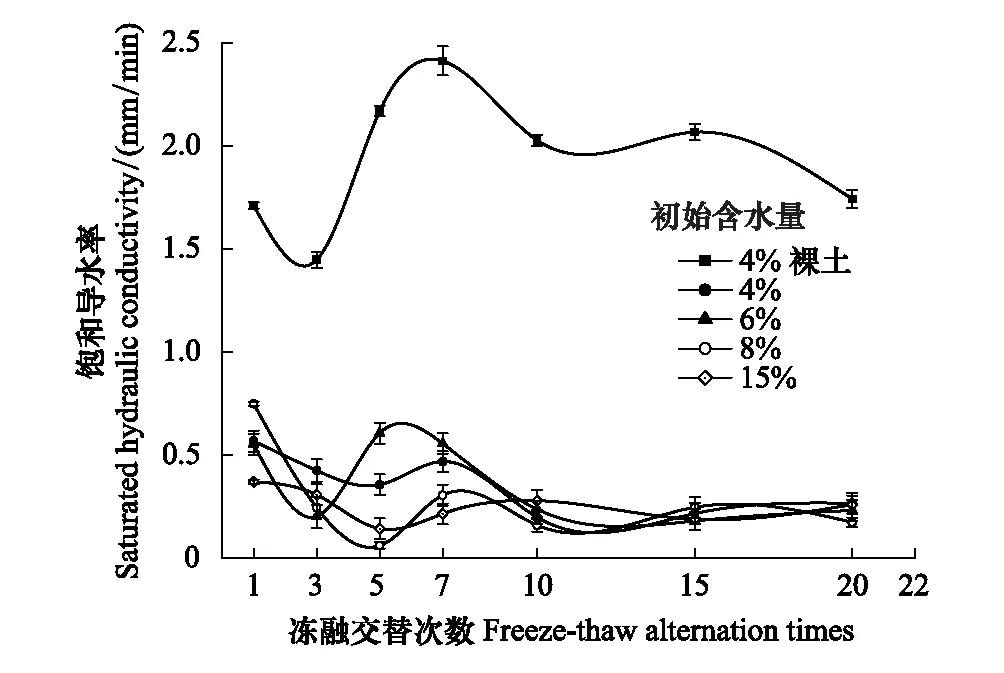
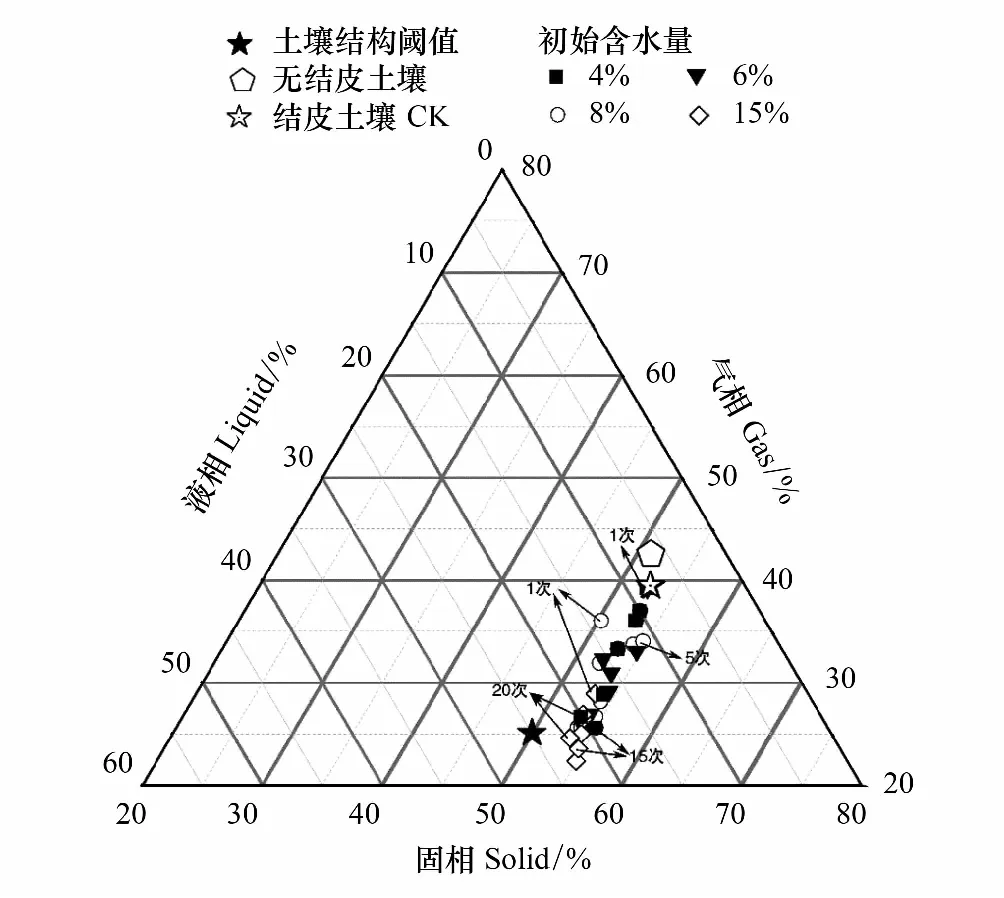
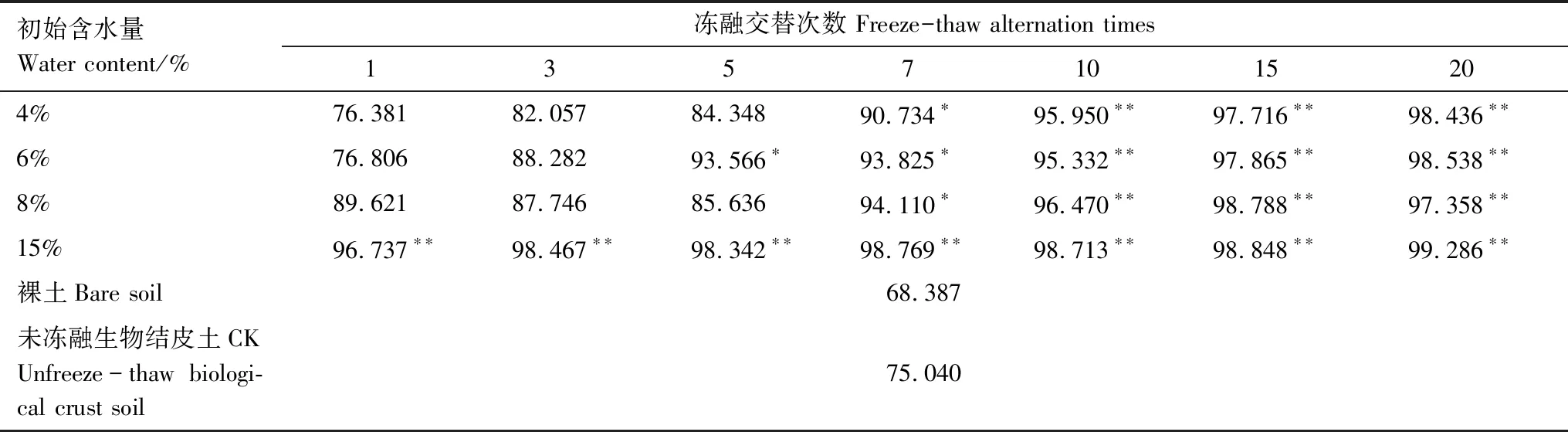
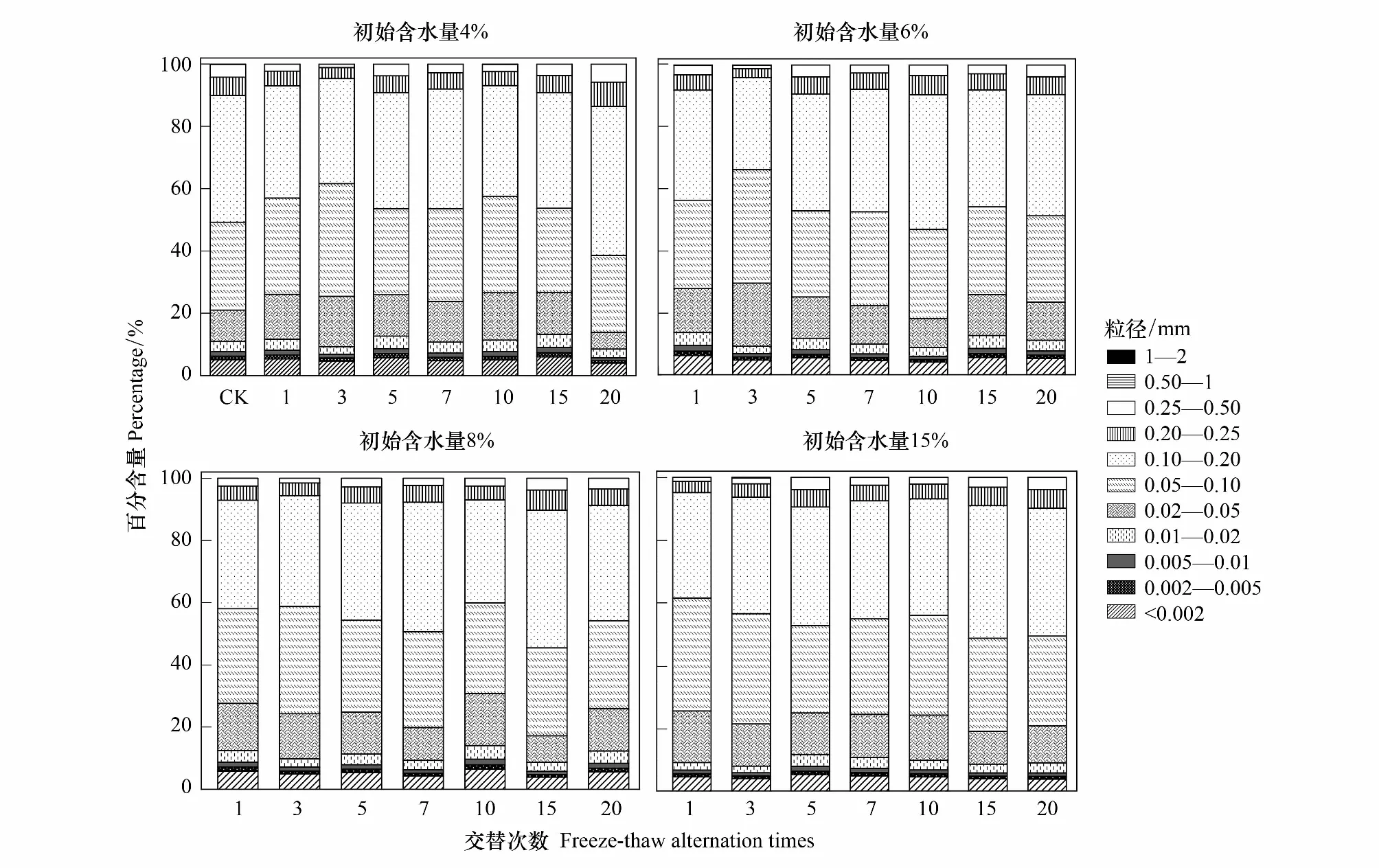
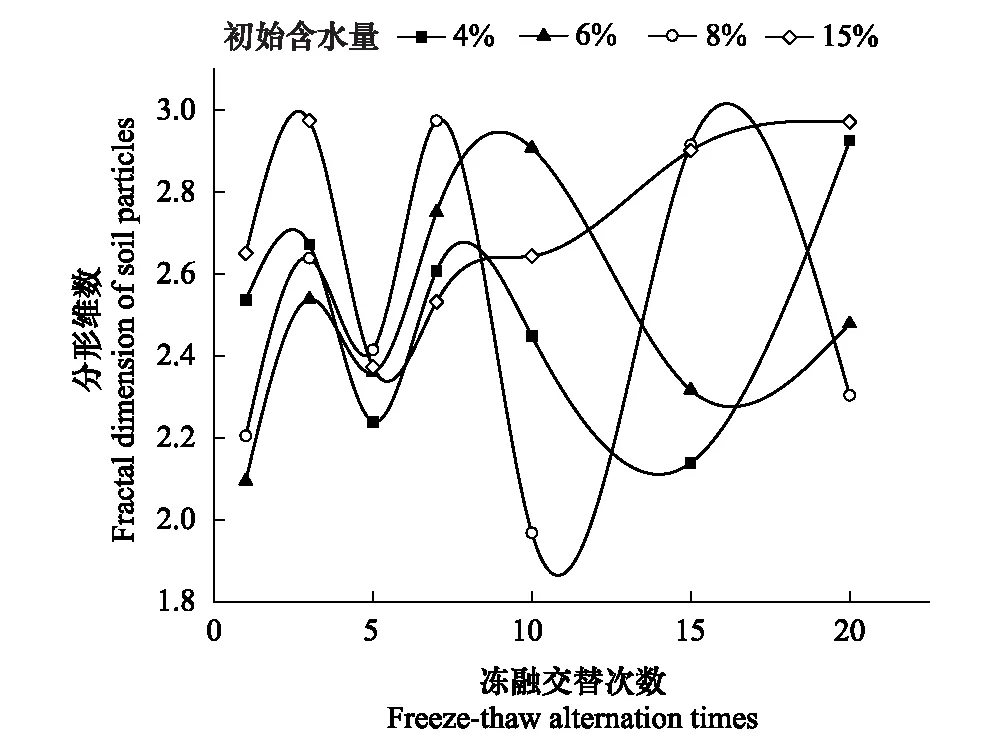
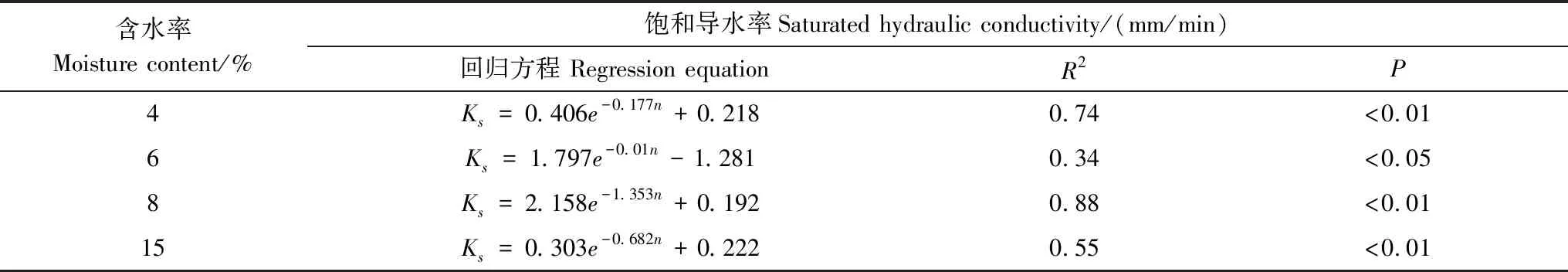
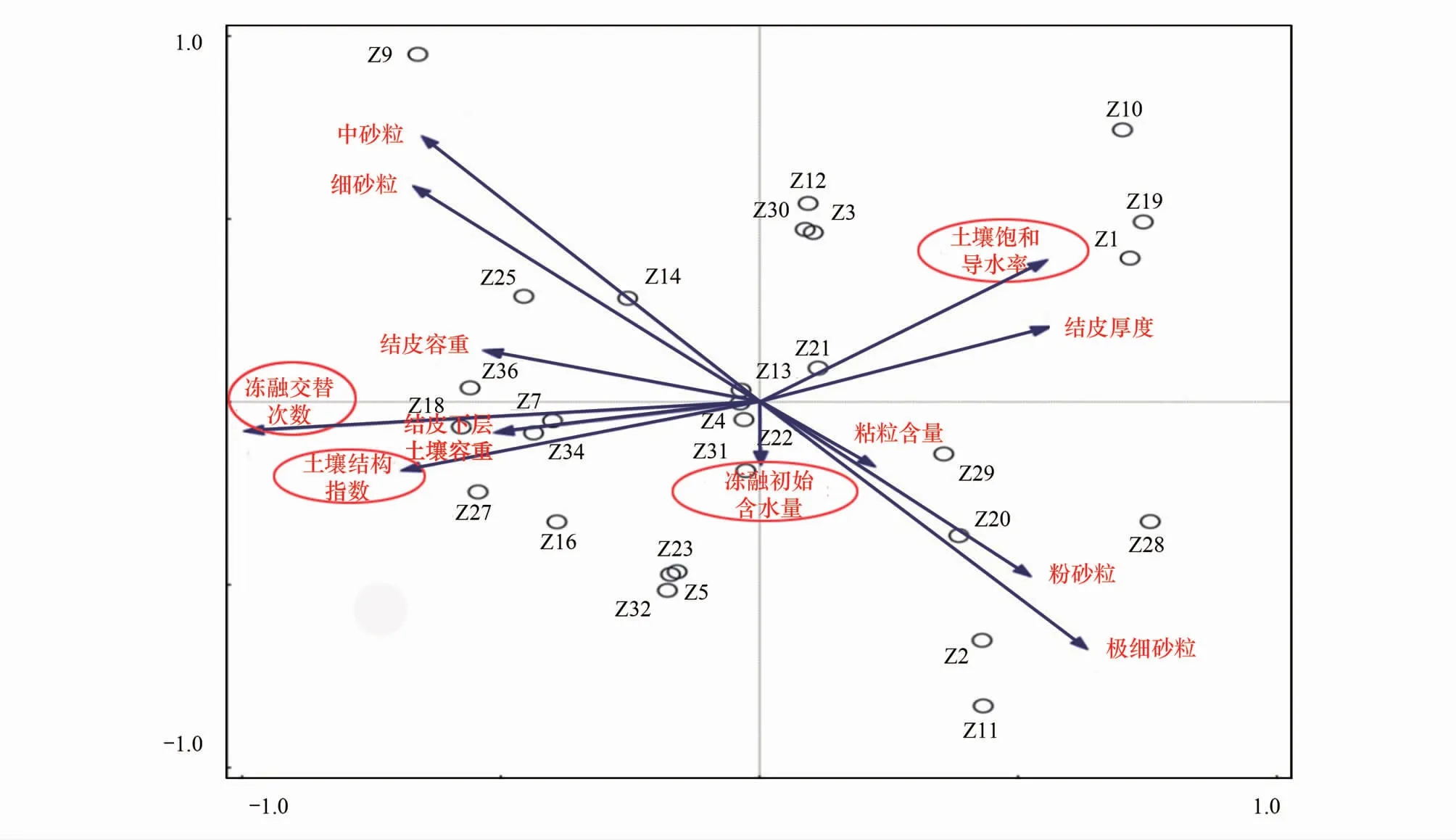
式中,V(r 土壤饱和导水率的计算公式为[25]: 式中,Kt为温度为t℃时的土壤饱和导水率(mm/min),Q/T为单位时间内的出水量(mL/min),L为环刀中的土层的厚度(mm),A为环刀横截面的面积(cm2),h为水头高度(cm),由于不同温度下水的粘滞系数不同,因此为了方便比较,将其换算成10℃下的饱和导水率Ks(mm/min)。 式中,ηt为温度为t℃下的水粘度(Pa·S),η10为10℃下的水粘度(Pa·S)。 本文采用Excel 2010和 SPSS 23.0进行数据分析,应用单因素方差分析(One-way ANOVA)和LSD多重比较对结皮土壤饱和导水率、环境指标进行差异显著性分析以及交互作用分析。使用Canoco 5.0进行主成分分析(Principal Component Analysis,PCA),并借助Pearson相关系数得出饱和导水率与试验因子的相关性以及表征程度排序。 裸土在不同冻融交替次数下的土壤饱和导水率如图1所示,未冻融的裸土(CK)和结皮土壤的Ks分别为2.583 mm/min和0.669 mm/min。4%含水量条件下,裸土的Ks随冻融次数增加而极显著变化(P<0.01),于3次时达到最小值1.447 mm/min,较CK下降44.0%;冻融7次时达到较大值,此时土壤的微观结构发生剧烈改变,水的冻结膨胀使土粒间的孔隙扩张连接导致土壤垂向渗透性能大幅提高[26],并在10次后逐渐稳定,Ks逐渐趋近于1.945 mm/min。而结皮土壤的Ks最小值较未冻融的结皮土壤下降70.1%,降幅显著大于裸土。冻融3次前,裸土较CK的Ks下降幅度大于结皮土壤,但随着冻融交替次数增加,结皮土壤较CK的降幅大于裸土。裸土的Ks未经冻融时,其饱和导水率是结皮土壤的3.86倍,这是由于生物结皮的存在,使土壤表面的导水孔隙封闭,随冻融次数变化,两者的差异呈波动上升,并在10次时裸土的Ks为结皮覆盖土壤的10.13倍,差异达到最大,表明结皮覆盖作用使土壤的饱和导水率显著下降,冻融作用使渗透能力进一步降低。结皮土壤在4%、6%、8%、15%含水量下随着冻融次数增加至10以上,Ks分别趋近于0.228 mm/min、0.216 mm/min、0.194 mm/min、0.242 mm/min,存在极显著差异,说明土壤的初始含水量会显著影响经过多次冻融后的饱和导水率的渐进值。 图1 不同冻融交替次数下结皮覆盖土壤饱和导水率变化Fig.1 Change of saturated hydraulic conductivity of crust-covered soil under different freeze-thaw cycles 在不同的土壤初始含水量条件下,随冻融交替次数增加,Ks差异明显,总体呈现平滑曲线式下降并最终趋于稳定。其中,当初始含水率为4%,冻融10次时Ks达到最小值(0.200 mm/min),相较CK下降了70.09%,而含水量为6%时,Ks的最小值出现在15次(0.181 mm/min),较CK下降73%,含水量为8%和15%时,Ks最小值均出现在第5次(0.061 mm/min、0.143 mm/min)较CK分别下降了90.87%、78.6%;试验条件下,冻融7次时,试验中各含水量条件下的饱和导水率才与CK相比均有显著差异(P<0.05)。 冻融过程中土壤Ks处于动态变化过程,不同土壤初始含水量条件下所能达到的Ks峰值大小为6%>4%>8%>15%,较高初始含水量下,Ks达到峰值的时间则相对滞后,且波动幅度减小。通过比较不同含水率和交替次数下Ks的变异系数发现,随着含水率由4%增加至15%,Ks变异系数由0.354增加至0.628后降低并在8%时达到最大。总体上看,试验条件下的所有变异均属于中等变异,随冻融交替次数增加,变异系数降低,冻融作用对Ks的影响逐渐减弱。冻融7次时,不同含水量下Ks逐渐接近,并在10次时差异显著减小,此时Ks的平均值为0.219 mm/min,变异系数为0.20。机械组成和土壤结构的差异在经过冻融作用后愈加明显,影响程度及变化趋势还需要深入分析。 土壤中不同形态和大小的土壤颗粒按照一定的排列组合方式形成土壤结构,土壤结构的存在使土壤中水、气得以储存和运输,从而影响着土壤中重要的物理和生物过程,也同样影响着土壤的饱和导水率。广义土壤结构指数(GSSI)是以固、液、气三相为研究对象,借助柯布-道格拉斯生产函数构建的用以定量有效表达土壤结构的指标[23],而GSSI越高,土壤越接近结构阈值,从而表现出土壤结构改变的临界状态。试验条件下,对土样初始含水量的处理在一定程度上改变了土壤的三相比。经不同次数的冻融交替作用之后,土壤三项比结果如图2所示。一般情况下,旱作土壤三相结构的临界值是固相50%,气相和液相各25%[27]。生物结皮土壤表层具有极复杂的结构演替特征[28],砂质土壤松散,植物假根和菌丝的穿插作用导致其剖面土壤孔隙网状结构连续拓展,土体孔径弯曲度不断增加,结皮土壤孔隙度降低,相较裸土其土壤中水的通路增加,水分在土层中滞留时间延长。当冻融1次时,由于土壤初始状态和物质组成的差异导致不同含水量下的土壤结构差异最明显,而随着冻融次数增加至15次以上时,不同含水量间的土壤结构逐渐稳定,且均向临界值趋近,在图中表现为向结构阈值中心点聚拢。相同含水量条件下,频繁的冻融交替促使土壤结皮层与外界进行更多的热量和物质交换,使土壤水分增加并不断压缩土壤空气,土壤气相显著减小。土壤结构随冻融次数改变具有极显著差异(P<0.01)(图2),当含水量为8%时,土壤结构的分散程度较大,冻融作用对结构的影响较明显,15%含水量下土壤结构的改变较小。在解冻期,生物结皮能够增加土层未冻水含量[29],水的较高比热容使土壤在高含水量条件下冻结和融化过程滞缓,相同时间内土层三相相对稳定,因此随着含水量增加冻融交替对土壤结构的影响减弱,土壤在不同含水量条件下随着冻融次数增加其结构均向阈值发展。 图2 不同冻融次数下结皮土壤三相变化 Fig.2 Three-phase changes of crust soil under different freeze-thaw time 试验中不同土壤初始含水量和冻融交替次数下的土壤结构指数如表2所示,各含水量条件下,冻融7次后的土壤结构与CK相比均有显著差异,试验条件下,15%含水量在经过20次冻融后GSSI逼近于结构阈值,与CK相差24.25,两者液相与气相的比值相差0.761。因此,冻结过程中初始含水量与冻融交替的共同作用会改变土壤三相比,通过减少气相的比例,使土壤结构显著改变。由于冻融7次时土壤结构改变显著,此时土壤完全冻结,液态水体积膨胀减少气相占比并挤压土粒;另外,颗粒间的联结和静电吸附发生断裂,降低团聚体水稳性,颗粒破碎大量发生[3],使Ks显著改变。10次时土壤结构破坏和颗粒粉碎逐渐逼近阈值, 导致Ks第2次发生显著变化。 表2 不同冻融初始含水量和交替次数下结皮土壤的广义结构指数 土壤饱和导水率受土壤结构影响,而土壤结构是土壤固相颗粒的大小及其空间排列的方式。土壤固相主要由土壤颗粒组成,因此通过研究冻融条件下土壤粒级的变化来探究饱和导水率变化的原因。不同含水量水平下藻结皮土壤的粒径分布对冻融次数的响应如图3所示。按照美国土壤质地分级标准进行单因素方差分析发现,不同试验条件下随冻融次数变化,土壤中的极细砂粒(0.05—0.1 mm)(P<0.01),细砂粒(0.1—0.25 mm)和中砂粒(0.25—0.5 mm)(P<0.05)差异显著,而其它粒径的颗粒无显著变化。研究表明,土壤由物理结皮向生物结皮发育过程中,下层土壤粘粉粒含量逐渐增大,在水分入渗过程中堵塞非毛管孔隙从而使结皮土壤入渗性能降低[10,30]。冻融条件下,不同含水量的土壤在冻融后<0.1 mm的颗粒较CK(49.27%)均有增加,尤其在冻融3次和10次时较明显,此时对应的饱和导水率较低。冻融过程中,粒径在不同尺寸之间相互转化,大颗粒崩解为小颗粒后颗粒间的缝隙被填充[17],对饱和导水率有显著降低作用,主体间效应分析表明,冻融条件下有无结皮覆盖(P<0.01)对土壤粒径分布的影响强于冻融次数(P>0.05)。 图3 不同冻融交替次数下藻结皮土壤粒径分布Fig.3 Algae crust soil particle size distribution under different freeze-thaw cycles 王艳艳、Ganiyu等人的研究表明[31—32],土壤具有分形特征,由形状、大小各异的颗粒和孔隙连接而成,并表现出不规则的几何形状,分形理论能定量刻画土壤结构特征,描述土壤的水力特性。本试验条件下,土壤颗粒分形维数(Dp)的结果表明(图4),冻融交替次数和冻融初始含水量改变,Dp无显著差异,未冻融土壤的Dp为2.593;在冻融7次前,冻融交替对不同含水量下土壤几何形状的重塑作用一致,但冻融作用对颗粒几何特征的影响在8%含水量下尤其剧烈,含水量降低使土壤几何形状变化幅度减少。含水量15%时,冻融7次后分形维数随冻融次数增加而增大,饱和导水率则呈现相反趋势。不同含水量和冻融次数下,变异系数随含水量增加呈现先增大后降低的趋势,并在8%时达到最大值为0.139,此时Ks变异最明显。冻融作用对粒径在0.05—0.5 mm间的颗粒有一定影响,<0.1 mm的土壤颗粒在冻融3次和10次时有显著变化(P<0.05)而处于较大值,饱和导水率相应降低。冻融过程中砂粒通过冻胀作用使松散的土壤被压实[33],孔隙度显著降低,由此影响土壤中的三相比值,改变土壤的入渗条件。 图4 不同冻融初始含水量和交替次数下土壤颗粒分形维数 Fig.4 Fractal dimension of soil particles under different initial water content of freeze-thaw and alternating times 不同初始含水量条件下,结皮土壤的Ks与冻融交替次数呈现显著的负相关关系(表3)、因子交互作用分析结果(表4)如下: 当含水量为8%时,指数模型能较好的解释饱和导水率的变化。由拟合结果显示,冻融交替次数能表示饱和导水率变化的34%—88%,模拟效果受初始含水量的影响。通过对结皮覆盖,冻融交替次数,初始含水量,结皮容重,结皮厚度与Ks的单因素ANOVA分析发现,结皮覆盖作用对Ks有极显著影响,不同冻融交替次数对应的Ks呈现极显著差异(P<0.01)。冻融条件下,结皮容重、下层土壤容重和初始含水量对Ks的影响无统计学意义,而不同试验因子间的交互作用对Ks的影响结果表明,冻融次数与土壤初始含水量、结皮层容重、结皮厚度的交互作用对饱和导水率的影响较大。 冻融交替作用下土壤饱和导水率PCA排序分析表明(图5),Ks受环境因子和土壤性质影响明显,环境因子在第一维度上相关性大小排序为:土壤结构指数>结皮厚度>结皮下层土壤容重>冻融交替次数>冻融初始含水量>结皮容重,环境因子与第二排序轴相关性大小为:粘粒>细砂粒>粉砂粒>中砂粒>极细砂粒。一维排序轴反映环境和试验因子的信息,而第二排序轴则反映了土壤物理性质。因子对Ks的影响程度依次为:冻融交替次数>土壤结构指数>结皮厚度>结皮容重>结皮下层土壤容重>冻融初始含水量。7次冻融交替次数下各因子处于多度平均值,随条件改变,样品多数集中于二、三象限,且多为冻融10次以上。由样点间的离散程度得出含水量为8%时样点彼此间较分散,因此受到其它因子的影响较明显。冻融交替作用下,冻融循环次数、冻融初始含水量、结皮容重、结皮下层土壤容重与饱和导水率呈现负相关性,但相关程度有所差异,土壤结皮厚度与Ks呈正相关性。 表3 不同冻融初始含水率下土壤饱和导水率(Ks)与冻融交替次数(n)拟合关系 表4 因子间交互作用对饱和导水率(Ks)影响的显著性分析 图5 土壤饱和导水率主成分分析排序 Fig.5 Principal component analysis (PCA) ranking of soil saturated hydraulic conductivity 图中Z1—Z36为试验样品编号 结果显示裸土的饱和导水率在含水量为4%时是结皮覆盖土壤的的3.86倍,结皮覆盖显著降低了Ks,这与Assouline[8],Lichner[9],Wang[10],以及刘翔等[11]的观点一致。赵春雷[3],樊贵盛等[16]研究表明土壤饱和导水率经冻融循环后呈现总体减小的趋势,与本研究中所得结论一致,随着冻融次数增加,裸土和结皮土壤Ks显著降低,藻结皮土壤的饱和导水率明显低于裸土且随冻融次数增加差异逐渐增大,并在10次时达到最大,这主要是由于藻类特性和土壤中有机质、颗粒组成、土壤结构的差异导致土壤的冻融初始条件不同而引起的。表现为冻融初期,结皮土壤中的微生物获得足够的碳和氮,解冻后变得活跃,通过吸收水分导致拒水性停止[34],而吸水后包裹生物结皮的胶鞘膨胀堵塞气孔延缓了水分在结皮中的入渗[35]导致饱和导水率逐渐减小,部分细胞受损破裂以及胞外多糖含量改变,生物结皮的状态差异发生在冻融4次后[7],因此冻融3—5次时结皮层组成成分正发生不同程度的变化使得前期结皮土壤的Ks出现波动。随着冻融次数增加,严重冻融对结皮内的细胞造成永久性损害,导致结皮对Ks的影响减弱,结皮下层土壤的渗透性逐渐占据主导。主成分分析结果表明,冻融循环、结皮层容重、结皮下层土壤容重与Ks呈负相关关系,其中冻融交替次数,土壤结构(GSSI)对Ks的影响程度较大。当土壤被饱和时,结皮的斥水性降到最低,由于结皮容重普遍小于下层土壤容重,结皮厚度增加使单位体积内土壤重量相对减少,也使水分通过下层土壤的通路减少,通过紧实土层所消耗水的动能降低,单位时间内水通量增加,因此结皮厚度在冻融条件下与Ks呈现正相关性。 Cheng等[36]研究认为土壤水的性质、结构与土壤温度相互影响,结果表明,冻融次数达到7次时土壤结构发生显著改变,冻融作用降低了土体的抗蚀性和稳定性[37]。研究表明,一季冻融交替结束后各层土壤饱和度平均增加13.06%[38],本研究中土壤经过冻融后饱和度平均增加8.91%,冻融过程中气态水部分液化使土壤受到土水势的影响吸收水分,下层水分向表层聚集,间接增加土壤中的水分含量。Viklander等[39]表示松散土经过若干次冻融循环后孔隙度降低且趋向一个稳定的孔隙比,土壤三相逐渐稳定,这与本研究结果一致。张泽等[40]的研究则表明冻融交替会使土壤颗粒间相互转化并逐渐均匀细化,<0.1 mm的颗粒组成变化显著,由于Ks与粘粒呈负相关关系,0.1 mm以下颗粒在水分运动过程中随水流动填埋原有孔隙以及冻融作用后新形成的大孔隙[17],松散的风沙土经过冻融后孔隙比会降低,从而导致土壤饱和导水率降低。土壤颗粒的转化在冻融6次时达到最大,在冻结过程中,孔隙水变成冰晶导致体积增加约9%,使颗粒的微裂缝向宏观发展,颗粒缝隙间的水膜厚度增加导致原生矿物碎裂[41],对于极细砂粒影响最显著,细砂和中砂颗粒变化显著是中砂颗粒向细砂颗粒转化所致,后续的冻融作用使颗粒的棱角度降低,重新排列后粒度逐渐均匀化[4],可能是Ks在冻融7次后逐渐趋向一致的原因。陈卓等[42]通过土柱渗流试验得出土壤饱和渗透率与体积分形维数呈对数关系(R2=0.952),颗粒的体积分形维数越大,土壤渗透率越小,这能较好的解释15%含水量下和冻融3—7次时饱和导水率的变化,而4%、6%、8%的Dp与Ks的变化无明显规律可能是由于含水率较低时土壤孔隙广泛分布,颗粒变化并非影响Ks的主要原因。 结皮的存在一方面改变土壤的理化性质,使土壤对外界环境变化的响应发生改变,另一方面是生物结皮具有生态功能与冻融产生交互作用使土壤饱和导水率进一步变化。本研究结果表明,结皮的容重和厚度与冻融次数存在交互作用,结皮的热传递性能与土壤不同,结皮厚度会影响温度到达土壤下层的时间,导致土壤发生冻融的初始状态、时间有明显差异,由于结皮容重显著小于下层土壤,在温度降至零下时,植物体细胞内的自由水迅速降低,部分功能散失[43],低温对生物结皮结构的破坏程度大于土壤;同时,结皮下层土壤中细粒相较裸土明显增加[30],且冻融作用对<0.1 mm的土壤颗粒的破坏重塑作用最强,因此,生物结皮间接加剧了冻融作用对土壤结构的破坏从而导致最终饱和导水率显著减少。冻融条件下,尽管生物结皮对颗粒组成的影响大于冻融次数,但冻融次数对土壤饱和导水率起主导作用,结皮覆盖对于导水率的影响是有限的。 本研究重点考虑结皮土壤物理性质以及冻融交替对Ks的影响,冻融环境的长期影响导致饱和导水率降低,促进了地表径流的形成,下层不透水冻层的存在使表层土壤受侵蚀的几率增加,结皮层的存在使下层土壤粘化进一步降低了饱和导水率,然而相较结皮的影响,冻融次数对土壤饱和导水率起主导作用。此研究为探究春冬季节干旱半干旱地区结皮地表水文过程,土壤侵蚀评估和生态治理提供了理论基础,而冬季降雨和产流条件下结皮土壤的侵蚀特征和过程变化,还需要深入研究。 (1)试验条件下,风沙结皮土壤的饱和导水率显著小于裸土,冻融初始含水量显著影响饱和导水率的波动幅度、渐进值、上升峰值以及达到峰值的时间。随冻融交替次数增加,两者饱和导水率差距逐渐增大,并在10次时差距达到最大值为10.13倍,冻融作用使藻结皮土壤的饱和导水率呈类指数型下降并在冻融10次后逐渐稳定。 (2)冻融交替通过不断减少气相比例和改变风沙土壤颗粒组成使土壤三相比和结构显著改变,其中<0.1mm的颗粒受冻融影响显著,而结皮覆盖作用对风沙土壤颗粒组成的影响大于冻融次数。冻融7次时土壤结构破坏和颗粒粉碎严重,并在10次时趋近阈值逐渐稳定,含水量为8%时对土壤结构的影响较显著。 (3)冻融次数分别与冻融初始含水量、结皮层容重、厚度存在交互作用对风沙土壤饱和导水率具有显著影响,不同因子间对土壤饱和导水率大小的影响程度为:冻融交替次数>土壤结构>结皮厚度>结皮容重>下层土壤容重>冻融初始含水量。2 结果与分析
2.1 冻融交替作用对结皮土壤饱和导水率的影响
2.2 冻融条件下土壤结构变化对饱和导水率的影响
2.3 结皮覆盖土壤粒径分布对冻融交替的响应
2.4 冻融条件下土壤饱和导水率影响因素分析

3 讨论
3.1 冻融条件下结皮层特征对土壤饱和导水率的影响
3.2 土壤三相对冻融交替的响应
3.3 生物结皮与冻融的交互作用对土壤饱和导水率的影响
4 结论
