Cu2+对文蛤红壳色选育系及自然群体抗氧化能力的影响
2022-01-26张志东陈爱华吴杨平陈素华李秋洁
田 镇,张志东,陈爱华,吴杨平,陈素华,张 雨,曹 奕,李秋洁
( 1.江苏省海洋水产研究所,江苏 南通 226007; 2.上海海洋大学,水产科学国家级实验教学示范中心,上海 201306 )
文蛤(Meretrixmeretrix),属软体动物门、瓣鳃纲、帘蛤目、帘蛤科,是一种广温、广盐性的埋栖型贝类,是我国滩涂养殖的主要经济贝类之一[1]。近年来,由于重金属农药使用及工业废水排放等原因,养殖海水重金属含量超标。一些自然海域文蛤采苗区和养殖区出现文蛤富集中毒现象,影响了文蛤产业的发展[2]。
海水贝类重金属富集程度有所差别,一般通过吸附、滤食等方式,从周围环境积累重金属[3-5],导致贝类出现中毒反应[6]。相关研究表明[7-8],重金属进入贝类体内,贝类细胞内物质会发生氧化还原反应产生活性氧,同时刺激动物体内某些基因的表达,产生相关金属酶类增强体内抗氧化系统,从而减轻重金属对细胞的损伤;一旦体内氧化还原机制平衡遭到破坏,细胞造成损伤,机体会发生中毒反应。超氧化物歧化酶(SOD)和活性氧清除系统中的主要抗氧化酶——过氧化氢酶(CAT)是海水贝类对外界环境应激机制中重要的抗氧化酶,是清除自由基和活性氧的第一道防线。当机体受到环境胁迫时,超氧化物歧化酶和过氧化氢酶能迅速应答,因此其酶活性的变化在一定程度能反映贝类免疫机能的变化,已成为贝类免疫、生态药理学等相关研究的重要指标[9]。
笔者对文蛤自然群体及红壳色选育系进行Cu2+胁迫试验,统计在不同质量浓度Cu2+条件下两个文蛤群体存活率变化趋势,分析其超氧化物歧化酶和过氧化氢酶活性以及Cu/Zn-SOD基因的表达量变化情况,以期为选育抗逆性强优良品质文蛤新品系提供科学参考。
1 材料与方法
1.1 试验材料
本试验所用自然壳色文蛤(称自然群体)来源于江苏省如东县腰沙—冷家沙海域,红壳色选育系文蛤(称红壳色选育系)为原种文蛤经过5代选育所得,平均壳长(3.53±0.34) cm,平均体质量(13.32±2.80) g。文蛤清洗干净后放入试验箱中暂养5 d,每日换水,并定期投喂江苏文蛤良种场培养的球等鞭金藻(Isochrysisgalbana)和绿色巴夫藻(Pavlovaviridis)藻液,水体初始藻密度为1×105个/mL。
1.2 试验环境
将试验海水过滤、纯化。盐度29±0.55,pH 8.0±0.3,水温(18±2) ℃,溶解氧(8.35±0.74) mg/L,少量充气。利用药物CuSO4·5H2O(AR)制备质量浓度为5 g/L的Cu2+母液。
1.3 试验方法
1.3.1 文蛤存活率试验
利用Cu2+母液将质量浓度设置为0、0.1、0.2、0.3、0.4、0.5 mg/L 6个梯度,将暂养后的红壳色选育系和自然群体分别投入到各Cu2+梯度组中,每组各50个,设置3个平行组。试验期间停止投饵,微量充气持续观察试验文蛤活动状态,及时清除死亡个体,48 h后统计各组存活率。死亡判定标准为刺激无反应,双壳张开。
1.3.2 Cu2+胁迫试验
根据1.3.1试验结果来确定Cu2+质量浓度梯度,使得该试验在文蛤存活率高于50%环境中进行。试验箱中Cu2+母液作为胁迫的Cu2+源,使Cu2+的质量浓度分别为0、0.1、0.2、0.3、0.4 mg/L;将选育群体与自然群体分成5组,每组20个,投放于各质量浓度试验箱中,并设置3组平行。分别于3、6、12、24、48 h时随机选择3个个体取样,取样部位为鳃,并将样品立即置于-80 ℃冰箱,用于荧光定量PCR和酶活性测定。
1.3.3 抗氧化酶活性测定
在玻璃匀浆器中加入9倍体积的预冷生理盐水,冰上匀浆组织2 min,匀浆后于离心机上2500 r/min(离心半径8.6 cm)离心10 min,取上清液用于测定酶活。采用南京建成生物工程研究所的试剂盒测定超氧化物歧化酶和过氧化氢酶活性和总蛋白量,酶活性单位为U/mg。
1.3.4 总RNA提取、逆转录和荧光定量PCR
RNA提取采用常规TRizol RNA 提取方法进行,RNA提取完成后立即进行逆转录,逆转录步骤参考Revert Aid TM First Strand cDNA Synthesis Kit说明书。逆转录后的cDNA为模板稀释后作为荧光定量PCR的模板,以文蛤β-actin为内参基因,Cu/Zn-SOD基因引物(表1)进行荧光定量PCR试验,分析选育群体与自然群体在Cu2+胁迫下鳃组织的基因表达量及表达变化。

表1 PCR引物信息Tab.1 Primers used for PCR
1.4 数据处理
运用最小二乘法2-ΔΔCt法处理荧光定量PCR数据,用SPSS 22.0软件对存活率、酶活性和Cu/Zn-SOD基因相对表达量数据进行分析,数据用平均值±标准差表示,单因素方差分析试验数据的差异显著性(α=0.05)。通过Origin 8.0软件对数据进行处理并导出柱状图。
2 结 果
2.1 不同Cu2+质量浓度对文蛤群体存活率的影响
统计不同Cu2+质量浓度下文蛤的存活率(图1)。试验结果表明,红壳色选育系和自然群体的存活率随着Cu2+质量浓度的升高而逐渐降低,有着共同的趋势。在相同Cu2+质量浓度下,红壳色选育系和自然群体的存活率差异不显著(P>0.05)。当Cu2+质量浓度为0.5 mg/L时,红壳色选育系和自然群体的存活率均低于50%。
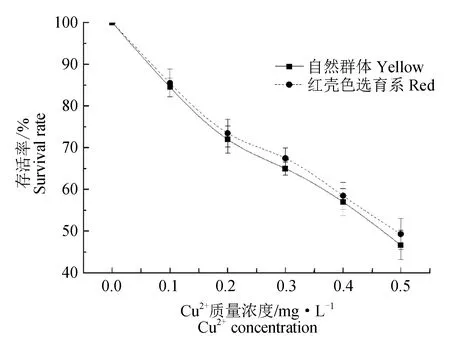
图1 不同Cu2+质量浓度下文蛤的存活率Fig.1 The survival rate of hard clam M. meretrix at different Cu2+ concentrations
2.2 不同Cu2+质量浓度对文蛤群体超氧化物歧化酶和过氧化氢酶活性的影响
Cu2+质量浓度为0 mg/L时,红壳色选育系与自然群体的超氧化物歧化酶活性差异不显著(P>0.05)(图2a);Cu2+质量浓度为0.1 mg/L时,2组文蛤随着胁迫时间延长呈升高趋势,超氧化物歧化酶活性差异显著(P<0.05)(图2b);Cu2+质量浓度为0.2~0.3 mg/L时,2组文蛤超氧化物歧化酶活性前期(3~6 h)变化缓慢,在后期(12~48 h)升高幅度较大(图2c~d);Cu2+质量浓度为0.4 mg/L时,2组文蛤超氧化物歧化酶活性随着胁迫时间延长逐渐增加(P<0.05)(图2e)。48 h时,2组文蛤超氧化物歧化酶活性随着Cu2+质量浓度增加呈上升趋势,除对照组(0 mg/L)外,红壳色选育系与自然群体差异显著(P<0.05)(图2f)。
Cu2+质量浓度为0 mg/L时,红壳色选育系与自然群体的过氧化氢酶活性无显著变化(P>0.05)(图3a);Cu2+质量浓度为0.1 mg/L时,2组文蛤过氧化氢酶活性随时间呈先降后升趋势,24~48 h时间段两者过氧化氢酶活性差异显著(P<0.05)(图3b);Cu2+质量浓度为0.2~0.4 mg/L时,2组文蛤过氧化氢酶活性差异显著(P<0.05),随时间变化呈先降后升趋势(图3c~e)。在48 h时,Cu2+质量浓度为0.2 mg/L组过氧化氢酶活性开始有上升趋势,并且与对照组(0 mg/L)差异显著(P<0.05),除对照组(0 mg/L)外,其他各组红壳色选育系与自然群体过氧化氢酶活性差异显著(P<0.05)(图3f)。另外,在试验过程中,添加不同质量浓度的Cu2+组3 h内红壳色选育系和自然群体均吐出大量黏液,足全部伸展,做掘土状,3 h后足开始收缩,双壳逐渐闭紧。
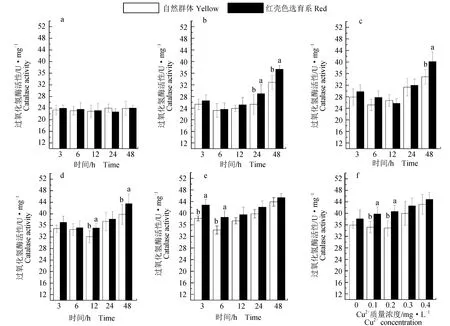
图3 不同Cu2+质量浓度下文蛤鳃组织过氧化氢酶活性随时间变化趋势Fig.3 Variation trend of catalase activity (CAT) in gill tissue of hard clam M. meretrix at different Cu2+ concentrationsa.0 mg/L Cu2+; b.0.1 mg/L Cu2+; c.0.2 mg/L Cu2+; d.0.3 mg/L Cu2+; e.0.4 mg/L Cu2+; f.48 h时红壳色选育系和自然群体鳃组织的过氧化氢酶活性随Cu2+质量浓度变化趋势.a.0 mg/L Cu2+; b.0.1 mg/L Cu2+; c.0.2 mg/L Cu2+; d.0.3 mg/L Cu2+; e.0.4 mg/L Cu2+; f.the change trend of CAT in gill tissue of red shell color breeding lines and natural populations at Cu2+ concentration at 48 h.
2.3 不同Cu2+质量浓度对文蛤群体Cu/Zn-SOD基因表达量变化
不同Cu2+质量浓度胁迫48 h后Cu/Zn-SOD基因表达量变化见图4。低Cu2+质量浓度时,红壳色选育系和自然群体的Cu/Zn-SOD基因表达量水平差异不显著(P<0.05);Cu2+质量浓度为0.2~0.4 mg/L时,Cu/Zn-SOD基因表达量水平呈现上调且红壳色选育系和自然群体的Cu/Zn-SOD基因表达水平差异显著(P<0.05)。

图4 48 h时文蛤鳃组织Cu/Zn-SOD相对表达量随Cu2+质量浓度变化趋势Fig.4 Variation trend of Cu/Zn-SOD relative expression level in gill tissue of hard clam M. meretrix with Cu2+ concentration at 48 h
3 讨 论
3.1 重金属Cu2+对文蛤存活率的影响
随着沿海工业的发展,水体中重金属污染已成为水产养殖不可忽视的风险因素之一。加之贝类多集中分布在沿岸滩涂上,移动性差,可通过吸附和滤食的方式,直接从周围环境积累重金属。大量研究表明,重金属Cu2+对海水贝类的生长繁殖会产生不利影响,甚至导致海水贝类的大量死亡[10-11]。
本试验结果表明,当Cu2+质量浓度为0.5 mg/L时,红壳色选育系的存活率比自然群体略高,但差异并不显著(P>0.05),且48 h后存活率均低于50%。虽然Cu是生物体必需的一种微量元素,在机体代谢过程中起到重要的作用,但当其含量超过一定的限度时,会对机体造成不可逆转的毒害作用。造成上述结果的原因可能是,在选择育种的过程中,虽然仅以“壳色和生长”为目标性状进行定向选育,但选育出来的品系可具有其他的特性。杜晓东等[12]以“壳长+壳宽”为选育指标,采用群体选育辅以家系选育技术,经连续5代选育出的马氏珠母贝(Pinctadamartensi)“海选1号”,其不但具有较优的生长速度,还具有育珠期间母贝的留核率更高、珠层厚度更厚和珍珠产量高的表型差异。本试验中,文蛤红壳色选育系对Cu2+的耐受性的适当提高,与上述研究结果类似。因此,需要后续深入研究才能充分阐述新品系存活率提高的机理。
3.2 重金属Cu2+对文蛤超氧化物歧化酶和过氧化氢酶活性的影响
文蛤作为无脊椎动物,体内仅有由血细胞主导的非特异性免疫[13],温度、饵料、重金属等因素均会对贝类体内的超氧化物歧化酶和过氧化氢酶活性产生较大的影响。超氧化物歧化酶和过氧化氢酶是生物体内抗氧化物酶体系中重要的两种酶,在重金属压力刺激下的机体抗氧化免疫反应和损伤修复中起重要作用[14]。本试验结果表明,随着Cu2+质量浓度上升,红壳色选育系和自然群体体内超氧化物歧化酶和过氧化氢酶活性明显上升,这属于生物机体对外界环境胁迫做出的应激反应。在较低Cu2+质量浓度时,文蛤过氧化氢酶活性出现下降趋势,后随Cu2+质量浓度上升,又呈现升高趋势。这可能是由于“毒物兴奋作用”导致过氧化氢酶活性在生物体受到较为轻微污染的时候经常会出现降低趋势,当增加毒物污染时间又呈现上升趋势[15-16]。这与在中华绒螯蟹(Eriocheirsinensis)[17]、栉孔扇贝(Chlamysfarreri)[18]、紫贻贝(Mytilusgalloprovincialis)[19]中所表现的先抑制后诱导趋势相一致。而此次试验中超氧化物歧化酶活性是随质量浓度的增加和时间的延长而升高,这一趋势与在青蛤(Cyclinasinensis)[20]、近江牡蛎(Crassostreaariakensis)[21-22]、虾夷扇贝(Patinopectenyessoensis)[23]、菲律宾蛤仔(Ruditapesphilippinarum)[24]等中所表现出的先诱导后抑制趋势不同,可能是由于此次试验的胁迫时间过短,未观察后续的抑制过程。诱导上升可能是细胞和组织受到轻度破坏时,因受到污染物胁迫而不断产生新的超氧化物歧化酶使得氧化还原体系保持平衡,这是机体自身正常的应激反应[25];而下降趋势可能是由于生物受胁迫时间延长,体内自由基过多,抗氧化体系平衡被破坏所致[26]。另外,在试验过程中,添加不同质量浓度的Cu2+试验组3 h内红壳色选育系和自然群体均吐出大量黏液,足全部伸展,做掘土状;3 h后足开始收缩,双壳逐渐闭紧。这是文蛤受到污染物胁迫,机体受应激后逐步适应的生理表现。而文蛤黏液的产生与抗氧化活性之间关系需进一步研究。
3.3 重金属Cu2+对文蛤Cu/Zn-SOD基因相对表达量的影响
Cu/Zn-SOD基因在阻止金属毒性胁迫、保护细胞免受氧化损伤方面发挥重要作用,当机体遭受重金属氧化损伤时,Cu/Zn-SOD基因高表达可有效消除过多的氧自由基,降低和阻止细胞损伤[27-28]。本试验结果表明,自然群体和红壳色选育系在不同Cu2+胁迫下,48 h时间段的Cu/Zn-SOD基因表达量变化均呈现质量浓度—表达量线性关系,与虾夷扇贝、海湾扇贝(Argopectenirradians)[29]等贝类表达趋势相类似,但与皱纹冠蚌(Cristariaplicata)[30]、湖北钉螺(Oncomelaniahupensis)及中华圆田螺(Cipangopaludinacahayensi)[31]等淡水贝类24 h后表达量出现下降不同;并且红壳色选育系Cu/Zn-SOD基因相对表达量在同一Cu2+质量浓度下与自然群体相比差异显著,这与超氧化物歧化酶活性的结果趋势相一致,也说明了红壳色选育系文蛤在一定Cu2+质量浓度下的抗氧化性优于自然群体。这可能源于红壳色选育系是课题组以“壳色+生长”采用闭锁群繁选育的新品系,在壳色和生长性状对某些多功能优良基因变异有一定程度累积[12]。
4 结 论
通过设计不同Cu2+质量浓度对文蛤自然群体及红壳色选育系进行胁迫试验,获得Cu2+质量浓度胁迫下红壳色选育系和自然群体的超氧化物歧化酶和过氧化氢酶的活性变化,共同表现出相应的时间—效应关系和质量浓度—效应关系;同时采用荧光定量PCR技术检测了Cu2+胁迫下Cu/Zn-SOD基因相对表达量,其在两组文蛤体内与Cu2+质量浓度呈正相关关系。综上所述,红壳色选育系的Cu2+抗氧化性在一定程度优于自然群体。这可能源于红壳色选育系是以“壳色+生长”采用闭锁群繁选育的新品系,在壳色和生长性状对某些多功能优良基因变异有一定程度累积,如SCD、CPOX基因是文蛤壳色性状重要控制基因,同时也是贝类参与免疫反应的关键基因[31]。本试验结果将对文蛤抗逆性选育和新品种的开发提供科学参考基础。
