牦牛TLR-4基因3′-UTR区扩增克隆及相关miRNAs生物信息学分析
2022-01-18栗亮亮温东旭徐业芬王运路常新明赵启涵秦忠璞索朗斯珠牛家强
栗亮亮 , 温东旭,2 , 徐业芬,2 , 王运路,2 , 常新明 , 赵启涵 , 秦忠璞 , 索朗斯珠,2 , 牛家强,2
(1. 西藏农牧学院动物科学学院 , 西藏 林芝 860000 ; 2. 西藏农牧学院西藏高原动物疫病研究自治区高校重点实验室 , 西藏 林芝 860000)
牦牛起源于中国青藏高原,早期家养牦牛由野牦牛驯化而来,在当地农牧民生产生活和畜牧业发展中占有重要地位,是最具高原特色的经济性动物。虽然经过长期的进化,牦牛已经能够适应高海拔、寒冷、低氧等环境,但受到遗传因素和环境的影响,牦牛生长缓慢且疾病多发。近年来,细菌耐药性增强的研究报道屡见不鲜,免疫接种也未能达到预期效果。因此,如何提高牦牛群体免疫力,降低农牧民损失是亟需解决的问题。抗病育种是近年来研究的热点之一,可通过选育抗病力强的个体进行育种从而提高群体免疫能力,避免过度依赖药物和疫苗。Toll样受体(Toll-like receptors,TLRs)基因是重要的抗病候选基因之一,表达产物为 Ⅰ 型跨膜蛋白受体,可识别多种细菌、病毒等微生物的固有成分,激活相应信号通路,从而引发机体免疫反应,其中TLR-4受体在单核细胞、巨噬细胞、树突状细胞、B细胞系等免疫细胞上均有表达,主要识别革兰阴性菌细胞壁组成成分脂多糖(Lipopolysaccharide,LPS)[1],并与烟酰胺磷酸核糖转移酶结合后可诱导增强核因子(Nuclear factor-κB,NF-κB)的转录活性,调节肺组织炎症损伤及天然免疫过程[2]。
微小RNA(microRNA,miRNA),是一种长度大小为18~25个核苷酸的内源性非编码RNA,主要通过靶向mRNA的3′非编码区(3′-Untranslated region,3′-UTR)抑制基因的翻译,进而在生长发育和免疫调节等过程中发挥调控作用[3]。研究显示,牛血清中miRNAs在口蹄疫感染的不同时期表达量存在差异,揭示miRNAs可能与牛口蹄疫免疫调节相关[4],此外发现miR-146a和miR-181c可能通过靶向TLR-4基因参与调控奶牛乳腺炎的发生发展[5],但鲜见牦牛miRNAs的研究报道。本试验旨在扩增克隆牦牛TLR-4基因3′-UTR区,并对可能靶定TLR-4基因miRNAs进行生物信息学分析,探索牦牛体内miRNAs与TLR-4基因的3′-UTR区可能靶向关系,为牦牛疫病防治及抗病性育种提供一定理论基础。
1 材料与方法
1.1 材料与试剂仪器
1.1.1 试验动物 牦牛血样共341份,分别来源于青海称多县30份、湟源县58份、大通县208份、西藏林芝市周边15份、波密县康玉乡30份,牦牛免疫程序及饲养条件基本一致。用ACD抗凝管进行颈静脉采集血样,采集完毕后立刻置于-20 ℃冰箱内保存。
1.1.2 主要试剂 T5全血直扩试剂盒、琼脂糖、核酸染料,均购自成都擎科梓熙生物技术有限公司。
1.1.3 主要仪器 PCR仪,美国ABI公司产品;电泳仪和电泳槽,北京六一生物科技有限公司产品;凝胶成像仪,美国GEN公司产品。
1.2 试验方法
1.2.1 引物设计 根据NCBI中牦牛XM_005891938.1序列号,由成都擎科梓熙生物技术有限公司合成引物。上游引物:5′-GGAGAATCCCCTGATGTGCT-3′,下游引物:5′-ATGGGTTGATAGAGAAGACGTGG-3′,目的片段大小为623 bp。
1.2.2 DNA混池构建 将341份血样分别吸取20 μL构建5个DNA混池,分别命名为T1、T2、T3、T4和T5。T1含波密县康玉乡30份牦牛血样,林芝周边15份牦牛血样;T2含青海湟源县58份牦牛血样;T3含青海大通县40份牦牛血样;T4含青海大通县102份牦牛血样;T5含青海称多县30份牦牛血样,青海大通县66份牦牛血样。
1.2.3 PCR反应 PCR反应总体系(50 μL):T5 Mix 25 μL,上游引物2 μL,下游引物2 μL,牦牛血样5 μL,加ddH2O至50 μL。PCR反应条件:94 ℃预变性5 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸50 s,扩增52个循环;72 ℃延伸10 min,4 ℃保存。扩增产物采用2%琼脂糖凝胶电泳检测,并送至成都擎科梓熙生物技术有限公司进行测序。
1.2.4 miRNAs预测 通过TargetScan 7(http://www.targetscan.org/vert_72/)数据库对可能靶定TLR-4基因的miRNAs进行预测。物种选择牛,靶定基因输入TLR-4,其余参数默认。
1.3 数据分析 采用DNAMAN 8.0软件进行序列对比分析。
2 结果
2.1TLR-4基因的PCR检测 PCR反应结果见图1。通过测序对比发现,TLR-4基因3′-UTR区的扩增产物为623 bp,与预期片段大小基本一致,可以进行下一步试验。
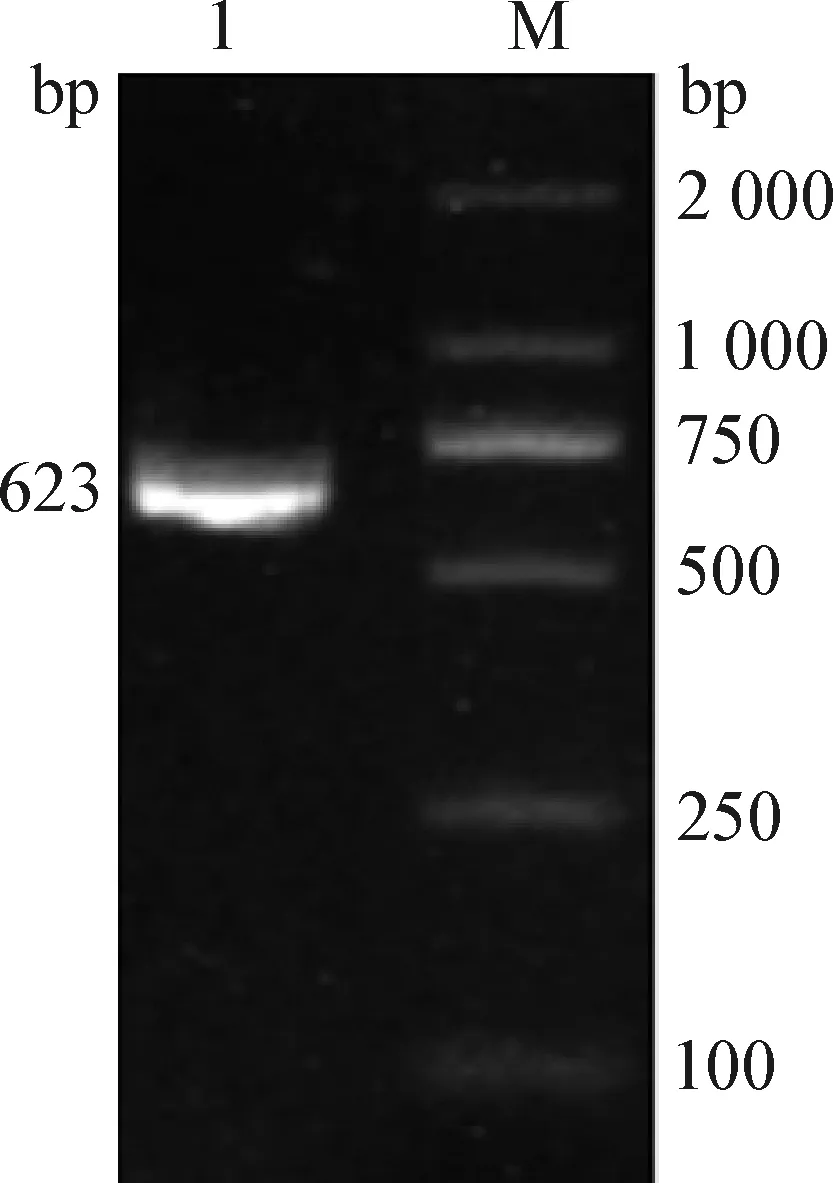
图1 TLR-4基因的PCR扩增Fig.1 PCR amplification of TLR-4 geneM:DNA分子量标记(DL2 000 DNA marker);1:牦牛TLR-4基因3′-UTR区扩增片断M:DNA molecular weight marker(DL2 000 DNA marker);1:Amplified fragment of 3′-UTR region of TLR-4 gene in yak
2.2 DNA混池测序 DNA混池测序结果与GenBank中牛TLR-4基因序列相比较,检测出扩增片段可能含有2个单核苷酸位点(Single nucleotide polymorphism,SNP)发生突变(图2),其中T1组TLR-4基因3′-UTR区在存在A382G突变,T3组TLR-4基因3′-UTR 区未检测到突变,T2组、T4组、T5组TLR-4基因3′-UTR区均检测到A380G、A382G两处突变。
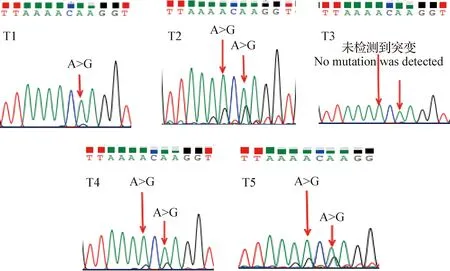
图2 DNA混合池测序结果Fig.2 Results of DNA mixing pool sequencing
2.3 miRNAs预测 通过TargetScan 7数据库进行预测,物种选择牛,靶定基因为TLR-4,其余参数默认,共预测到18个可能靶向牛TLR-4基因3′-UTR区的miRNAs,具体见表1。

表1 TargetScan 7数据库靶定TLR-4基因3′-UTR区的miRNAs预测Table 1 Prediction of miRNAs in 3′-UTR region of TLR-4 gene based on TargetScan 7 database
3 讨论
TLRs是天然免疫中重要的模式识别受体之一,最初在果蝇中发现,至今在哺乳动物中共发现了13种不同的TLRs,可在多种细胞上表达,并通过识别病原微生物细胞内、外的固有成分诱导炎性因子产生,参与天然免疫反应[6]。其中TLR-4是目前研究较为广泛的受体之一,可识别革兰阴性菌LPS[1]、呼吸道合胞病毒的融合蛋白[7]、枯氏锥虫的糖基磷脂酰肌醇锚定蛋白[8]等成分。研究显示,TLR-4受体被激活后可通过NF-κB/MAPK信号通路上调乳腺炎相关的促炎细胞因子TNF-α、IL-1β和IL-6的表达[9],其与乳腺炎、肺损伤及纤维化[2]、免疫排斥反应[10]等密切相关,而且转录后水平的调控对TLR-4受体蛋白的表达具有重要影响。研究发现,基因的3′-UTR区是转录后水平调控的重要区域,其参与miRNAs的定位、稳定性和翻译过程,并可调节蛋白质之间相互作用,可将3′-UTR区遗传信息传递给蛋白质,在遗传中发挥重要功能[11],如在地中海水牛干扰素γ(Interferon gamma,IFNG)基因[12]和奶牛中性粒细胞胞浆因子4(Neutrophil cytosolic factor 4,NCF-4)基因[13]3′-UTR区均存在1个SNP位点,分别与结核病和乳腺炎相关,但目前鲜见牛TLR-4基因3′-UTR区相关研究报道。医学研究表明,人类TLR-4基因3′-UTR区存在3个SNP位点,与呼吸道合胞病毒(Respiratory syncytial virus,RSV)疾病调控有关[14]。本试验成功扩增了牦牛TLR-4基因3′-UTR 区片段,产物大小为623 bp,DNA混池测序结果显示,TLR-4基因3′-UTR区可能存在A380G和A382G 两个突变位点,但在牦牛上是否也与乳腺炎、结核病及RSV等疾病相关,仍需进一步研究确认。
MicroRNA是一种内源性非编码微小RNA分子,参与细胞分化、机体生长发育、免疫应答、肿瘤发生等过程,可在多种动物体内均有表达[15],并表现为高度保守性。MicroRNA主要通过与3′-UTR区靶向结合,在转录后水平上对基因的表达具有重要调控作用,参与机体多种疾病过程[3]。本试验通过TargetScan 7数据库共预测到18个可能靶向牦牛TLR-4基因3′-UTR区的miRNAs,但这些miRNAs在动物上鲜有报道,大多与人类多种癌症及炎症相关。与TLR-4基因相关的有miR-448和bta-miR-216a-5p,miR-448[16]可通过调节巨噬细胞极化作用抑制TLR-4基因表达,与人类糖尿病发展有关,而bta-miR-216a-5p[17]可与TLR-4受体3′-UTR区靶向结合,抑制TLR4-MyD 88信号通路和炎性细胞因子的产生,与梅毒病相关。bta-miR-338、bta-miR-146a和bta-miR-146b在人类和动物上研究均较为深入,bta-miR-338在牛骨骼肌中差异表达,可影响牛肌肉蛋白质水解和嫩度[18],miR-338-3p可靶向大鼠肉瘤相关蛋白23(Ras-related protein-23,RAB-23)基因的3′-UTR区,调控前列腺癌[19]的发展。miR-146a[20]可抑制人自分泌的IL-6和IL-21产生,阻断辅助性T细胞17分化进程,与自身免疫性疾病相关。此外,bta-miR-146a以牛肿瘤坏死因子受体相关因子6(Tumor-necrosis factor receptor-associated factor 6,TRAF-6)基因的3′-UTR区为靶点,下调TLR-4/TRAF-6/NF-κB通路,对牛乳腺炎及先天性免疫有负反馈调节作用[21],bta-miR-146a还与肺泡巨噬细胞具有特异性,上调牛结核病相关miRNAs,激发癌基因表达[22],在牦牛上可通过调节Smad4基因表达影响牦牛卵泡发育[23],bta-miR-146b过度表达对牛雄性生殖细胞增殖有抑制作用[24],另有研究表明,其与乳腺发育、免疫应答相关[25]。因此,本试验预测筛选到的18个miRNAs与牦牛TLR-4基因3′-UTR 区2个SNP位点是否存在真实靶向关系及在疾病防控和免疫方面能否起到作用,有待深入研究。
