褪黑素诱导采后冬枣抗黑斑病的研究
2022-01-07张乐乐常璐璐王小佳于有伟张少颖
张乐乐,常璐璐,王小佳,于有伟,张少颖
(山西师范大学食品科学学院,山西 临汾 041004)
冬枣作为一种常见的鲜食水果,不但皮薄肉厚,汁多无渣,脆甜可口,同时还含有丰富的果糖、VC 以及较高的环磷酸腺苷(cAMP)等生理活性物质,具有保肝护肝、增强免疫力等保健作用,备受广大消费者青睐。但是冬枣采后易受多种病原真菌感染导致腐烂,其中由链格孢菌侵染引起的黑斑病是冬枣采后最主要的病害之一[1]。因此,研究冬枣的采后防腐保鲜对减少其采后经济损失,增加商品价值有重大意义。
褪黑素(Melatonin,MT)不仅是一种微量多效的吲哚胺类物质,而且还作为一种生物活性分子调节植物的各种生理过程。MT 在促进植物侧根生长、种子萌发以及抗逆等过程中发挥重要作用[2]。MT 也可作为一种天然的氧化剂,在延缓果蔬衰老、抑制褐变、减轻氧化损伤,以及提高果蔬的抗逆胁迫等方面发挥重要作用[3]。在草莓果实应用中,MT 通过清除自由基保护膜完整性,延缓草莓果实的衰老[4]。MT 可以降低细胞膜通透性的变化,延缓荔枝褐变和衰老的进程[5]。MT 处理可延缓果皮褐变,降低相对电导率和MDA积累,提高膜完整性,提高石榴抗寒能力[6]。此外,MT在甜樱桃[7]、香蕉[8]等果蔬采后抗病方面的应用研究也日益增多。近年来研究表明,MT 还可以作为一种信号分子,在诱导果蔬采后抗病和抑制果蔬采后病原菌侵染方面发挥正向调控作用[9-10]。Yin 等[11]研究表明,MT 通过对Diplocarponmali 入侵和扩展的抑制作用提高了苹果对褐斑病的抗性。唐琦[12]也发现,采后MT 处理能够控制枣果实青霉病的发生,显著抑制Penicillium polonicum 的生长。MT 还参与香蕉[13]、西瓜[14]等水果的抗病。但是,目前有关MT 对采后病原菌的具体抑菌机理及其增强果蔬抗病性的机制还缺乏深入研究。
本研究采用不同浓度MT 处理接种链格孢菌的采后冬枣,探讨MT 对冬枣采后黑斑病的抗病作用,为进一步了解MT 在果蔬采后抗病中的作用提供理论依据,并为冬枣的采后黑斑病的防治提供新思路。
1 材料与方法
1.1 材料与设备
1.1.1 材料与试剂
供试的白熟期冬枣,于2020 年9 月28 日采自山西省临汾市尧乡冬枣种植基地。
链格孢菌(Altrenaria alternata),购于北纳生物有限公司,在PDA 培养基上培养备用。
褪黑素(生物试剂:99%),购于上海源叶生物科技有限公司。其他试剂均为分析纯。
1.1.2 仪器与设备
MDF-M53V 型超低温冰箱,日本三洋集团;LH199235 型折光仪,仪斯特电子有限公司;F-940 型便携式乙烯/氧气/二氧化碳分析仪,北京阳光亿事达科技有限公司;XSP-BM-2CA 生物显微镜,上海彼爱姆光学仪器制造有限公司。
1.2 方法
1.2.1 链格孢菌及孢子悬浮液的配制
参照文献[15]中的方法并稍有修改。将链格孢菌接种于PDA 培养基,在28 ℃条件下培养7 d。孢子成熟后,用生理盐水(含有0.05%吐温80)将孢子从平板上刮下,3 层纱布过滤多余菌丝,在显微镜下观察,最终使孢子悬浮液浓度为1×105个/mL。
1.2.2 冬枣的损伤试验
参考文献[16]的方法,采摘并挑选大小、形状一致,无机械损伤,无病害感染的冬枣,将果实平均分成4 组,先用自来水冲洗,再用2%次氯酸进行消毒浸泡2 min,自然晾干后进一步处理。分别用2、20、200 μmol/L MT 溶液浸泡8 min,以蒸馏水浸泡处理为对照(CK),自然晾干。将这4 种处理分成2 份:一份放入保鲜盒中用于测定果实的自然腐烂指数;一份于放置24 h 之后用无菌钉在4 组果实赤道附近表面打1 个直径为4 mm,深3 mm 的伤口,在伤口内接种5 μL 浓度为1×105个/mL 的链格孢菌孢子悬浮液,将接种好的枣果实放入保鲜盒中,用0.05 mm 聚乙烯膜封口后放置于室温条件下贮藏,每2 d 进行1 次取样,测定病斑直径、呼吸速率和失重率等指标。每组处理7 kg 果实,3 次重复。
将处理好的冬枣每2 d 留样,然后用液氮冷冻研磨成果粉,于-80 ℃超低温冰箱中保存,用于后期指标的测定。
1.2.3 测定项目与方法
1.2.3.1 病斑直径和腐烂指数
利用十字交叉法测量冬枣的病斑直径。
参照Cao 等[17]的方法,将腐烂面积分为0~4 级。
0 级,果面无腐烂;1 级,果面腐烂面积<1/4 果面;2 级,1/4≤果面腐烂面积≤1/2;3 级,1/2<果面腐烂面积≤3/4;4 级,果面腐烂面积>3/4。按照下列公式计算:

1.2.3.2 MDA、相对电导率、呼吸强度、失重率
MDA 含量:参照文献[18]测定。取枣粉1 g,加入5 mL 10%(m/V)三氯乙酸,在4 ℃10 000 r/min 离心20 min。然后取1 mL 上清液加入1 mL TCA 溶液,煮沸30 min,取出冷却,再次离心15 min,分别记录在450、532、600 nm 处的吸光值。
相对电导率:参照Zhang 等[19]的方法测定。随机选取5 个冬枣,用打孔器(10 mm)在枣果实接种面上打10 个枣圆片,把枣圆片冲洗之后用滤纸吸干水分。将枣片放入小锥形瓶中,吸取20 mL 去离子水,测电导率L0。然后将锥形瓶放在沸水中煮沸15 min,取出冷却至25 ℃,测其电导率L1。相对电导率计算公式为:
相对电导率(%)=L0/L1×100
失重率:依据文献[19]中的方法进行测定,计算公式如下:

呼吸强度:参照Tang 等[20]的方法,略有修改。将1 kg 枣果实放入大密闭容器罐中,罐口密封,室温静置1 h 后用便携式气体分析仪测密封罐子里的二氧化碳浓度。根据下列公式计算呼吸强度。

式中:K 为换算系数,21.43;V 为容器剩余空间体积(干燥器体积-冬枣体积),L;N 为CO2浓度,%;M 为冬枣质量,kg;H 为放置时间,h。
1.2.3.3 抗病相关酶活性和抗病相关物质含量
苯丙氨酸解氨酶(PAL)活性:参照文献[21]测定。取4 g 枣粉,加入4 mL 提取液,离心,留取上清液。加入0.5 mL 20 mmol/L L-苯丙氨酸和3 mL 50 mmol/L、pH 8.8 硼酸缓冲液,在37 ℃下保温10 min,再加入0.5 mL 酶液,测定混合液在290 nm 处的吸光值。于37 ℃保温60 min,测定第2 次混合液的吸光值。PLA活性单位为U/(h·g)。
几丁质酶(CHI)活性:参照文献[17]测定。取3 g枣粉,加入4 mL 预冷的提取缓冲液,离心,收集上清液。分别加入0.5 mL 乙酸-乙酸钠缓冲溶液(50 mmol/L、pH 5.2),0.5 mL 10 g/mL 胶装几丁质。然后,一组加入0.5 mL 酶液,一组加入0.5 mL 煮沸的酶液,在37 ℃保温1 h,之后继续加0.1 mL 30 g/L 的脱盐蜗牛酶,在37 ℃保温1 h 后,加入0.2 mL 0.6 mol/L 四硼酸钾溶液,最后煮沸3 min,取出冷却,于585 nm 处测吸光值。CHI 活性单位为U/mg。
β-1,3 葡聚糖酶(GLU)活性:参照文献[17]的方法,按照CHI 酶液提取方法,分别加入100 μL 4 g/L昆布多糖溶液,一只加入100 μL 煮沸的酶液,另一只加入100 μL 酶液。在37 ℃保温40 min,保温后将1.8 mL 蒸馏水和1.5 mL DNS 试剂加入2 只试管中,于540 nm 处测定吸光值。GLU 活性单位为U(/s·g)。
CAT 活性:参照文献[21]测定。取2 g 枣粉,加2 mL 提取缓冲液(5%聚乙烯吡咯烷酮和5 mmol/L二硫苏糖醇),4 ℃、12 000 r/min 离心30 min,加入2.9 mL 20 mmol/L H2O2溶液和100 μL 酶提取液,于240 nm 处测定吸光值的变化量。CAT 活性单位用U/g表示。
POD 活性:参照文献[17]中的方法测定。参照PAL 酶液提取的方法提取酶液。加入3 mL 愈创木酚和0.5 mL 酶液,再加入20 μL H2O2溶液,于470 nm测定吸光值变化。POD 活性单位用U/g 表示。
黄酮含量:参照文献[22]测定。称0.8 g 枣粉,加入4 mL 80%乙醇,离心。向试管中分别加入0.1 mL 上清液,再加入0.3 mL 5%NaNO2静置5 min,再加入0.3 mL 10%AlCl3溶液,反应6 min,最后加入2 mL 0.1 mol/L NaOH 溶液,于510 nm 处测定吸光值。黄酮含量单位为mg/100 g。
总酚含量:参考文献[23],略有改动。称0.8 g 枣粉,加入4 mL 80%乙醇,离心。向试管中分别加入0.1 mL 上清液,再加入1 mL Folin-Ciocalteau 试剂(稀释3 倍)混匀后静置5 min,再加入1 mL 10%Na2CO3溶液,室温避光放置90 min,于765 nm 处测定吸光值。总酚含量单位为mg/100 g。
木质素含量:参照文献[21],略有改动。取1 g 枣粉,加入4 mL 提取液,离心。用95%乙醇冲洗沉淀物3 次,再用乙醇和正己烷混合液洗3 次。加入1 mL 2 mol/L NaOH 溶液、2 mL 冰醋酸和0.1 mL 7.5 mol/L盐酸羟胺,再次离心,于280 nm 处测定吸光值。木质素含量以OD280/g 表示。
1.2.4 数据处理
使用Excel 2010 进行数据整理,SPSS 25.0 进行统计分析,Origin 2021 软件作图。
2 结果与分析
2.1 不同浓度MT 处理对接种A.alternata 冬枣果实病斑直径和自然腐烂指数的影响
由图1 知,适宜浓度MT 处理可以抑制冬枣病斑直径的扩展。不同浓度MT 处理的枣果实病斑直径均在4 d 后快速增大。处理6 d 后,200 μmol/L MT处理组的病斑直径均高于CK 组,2 μmol/L MT 和20 μmol/L MT 处理组的果实病斑直径显著低于CK组(P<0.05),其中20 μmol/L MT 处理的果实病斑直径最小。贮藏至16 d 时,20 μmol/L MT 处理的果实病斑直径分别比CK、2 μmol/L MT 和200 μmol/L MT 处理组低19.17%、15.36%和31.07%。
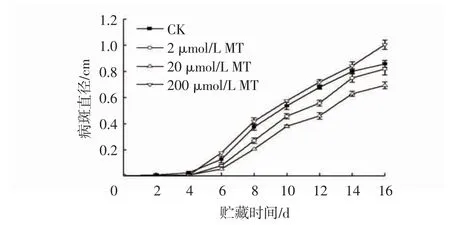
图1 MT 处理对冬枣病斑直径的影响Fig.1 Effects of MT treatments on colony diameters of winter jujube
由图2 可以看出,冬枣腐烂指数随贮藏时间的延长呈上升趋势,且对照组的自然腐烂指数在处理10 d 后大幅升高。与对照相比,MT 处理显著降低了冬枣的自然腐烂指数(P<0.05)。在贮藏结束时,对照组果实腐烂指数为4.2,20 μmol/L MT 处理组果实腐烂指数仅为1.5。
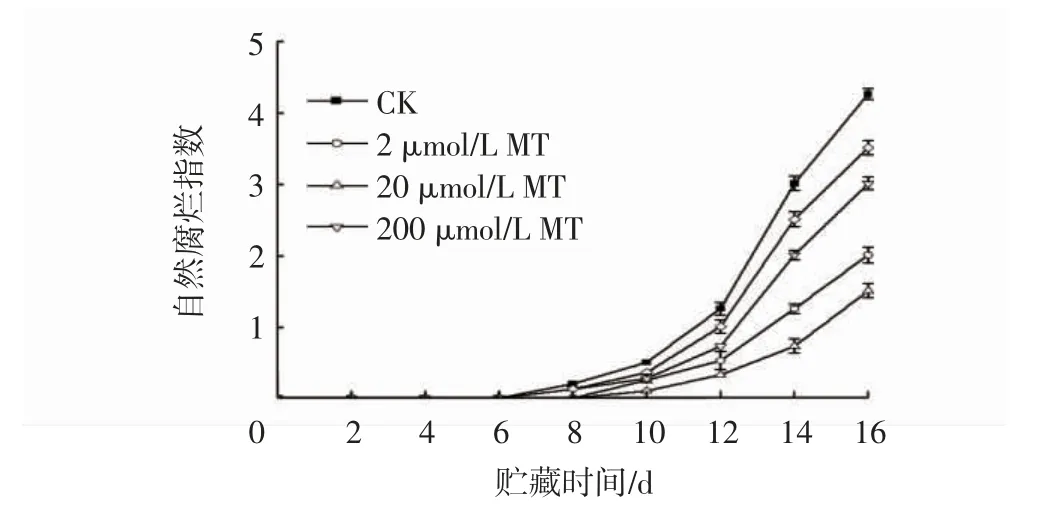
图2 MT 处理对冬枣自然腐烂指数的影响Fig.2 Effects of MT treatments on natural rot indices of winter jujube
2.2 MT 处理对冬枣果实MDA 含量、相对电导率、失重率和呼吸速率的影响
由图3 可知,随着贮藏时间的延长,MDA 含量呈先升高后降低的趋势。贮藏4~6 d,2 μmol/L MT 和200 μmol/L MT 处理组的MDA 含量均高于CK,而20 μmol/L MT 处理组MDA 含量始终低于CK 组,处于较低水平。贮藏第10 天和第16 天时,20 μmol/L MT处理组MDA 含量为各处理中最低,MDA 含量分别为1.09 μmol/mg 和0.28 μmol/mg,比CK 组低27.52%和36.13%。
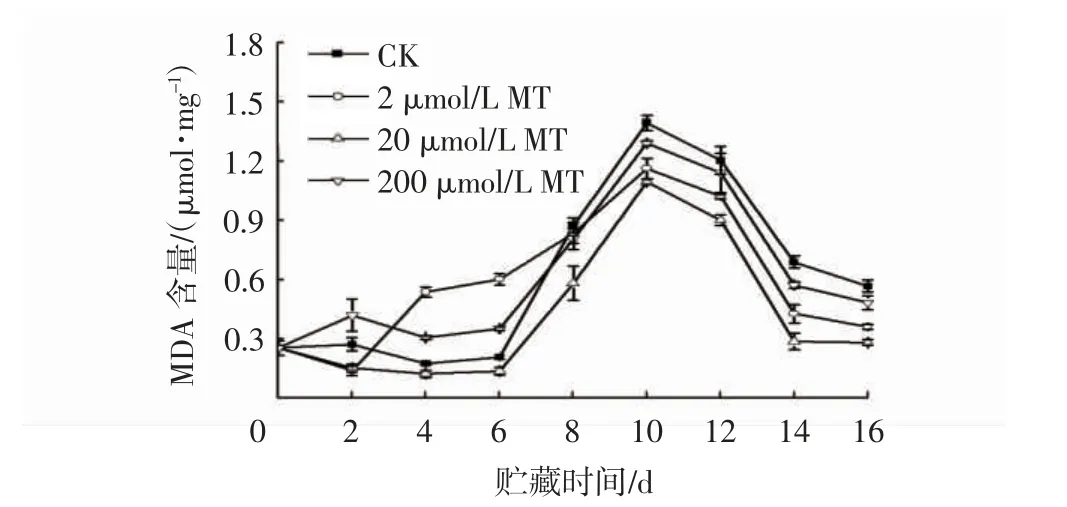
图3 MT 处理对接种链格孢菌冬枣果实MDA 含量的影响Fig.3 Effects of MT treatments on MDA contents in winter jujube inoculated with Alternaria alternata
由图4 可以看出,总体来讲,随着贮藏时间的延长,各组相对电导率均逐渐升高。与CK 组相比,MT 处理组的相对电导率均保持较低水平。其中,20 μmol/L MT 处理的相对电导率水平最低,上升最慢。贮藏16 d 时,2、20、200 μmol/L MT 处理组的相对电导率分别比CK 低11.55、21.42、3.55 个百分点(P<0.05)。

图4 MT 处理对接种链格孢菌冬枣果实相对电导率的影响Fig.4 Effects of MT treatments on relative conductivity in winter jujube inoculated with Alternaria alternata
由图5 可以看出,各处理组冬枣的失重率呈现不断上升的趋势。贮藏0~10 d 果实失重率缓慢增加,10 d 之后迅速增加。本试验中,MT 处理可以减少冬枣质量损失,CK 组的失重率显著高于MT 处理组(P<0.05)。贮藏第16 天时,CK 组果实失重率高达到11.5%,而20 μmol/L MT 的失重率最低,为6.1%。
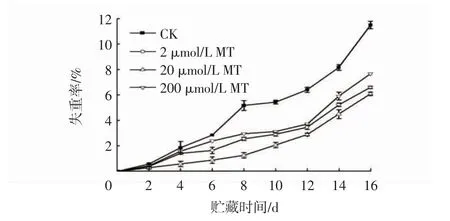
图5 MT 处理对接种链格孢菌冬枣果实失重率的影响Fig.5 Effects of MT treatments on weight loss rates of winter jujube inoculated with Alternaria alternata
由图6 可见,冬枣果实的呼吸强度均处于上升趋势,MT 处理显著抑制了冬枣的呼吸作用。其中,20 μmol/L MT 处理组果实呼吸强度随时间的延长上升趋势最为缓慢,显著低于2、20、200 μmol/L MT 处理组(P<0.05)。贮藏16 d 时,2、20、200 μmol/L MT 处理组果实呼吸强度分别比CK 低71.95%、70.78%和44.27%。显著性分析结果显示,在整个贮藏期间,CK组与各浓度MT 处理组间均存在显著性差异(P<0.05)。

图6 MT 处理对接种链格孢菌冬枣果实呼吸强度的影响Fig.6 Effects of MT treatments on respiration rates of winter jujube inoculated with Alternaria alternata
2.3 MT 处理对冬枣果实抗病相关酶活性和抗病相关物质含量的影响
由图7 可见,CK 组的PAL 活性整体呈现上升的趋势。与CK 组相比,MT 处理提高了冬枣的PAL 活性。贮藏第10 天,2 μmol/L MT 和20 μmol/L MT 组PAL 活性均达到最高峰,峰值分别为103.20 U/(h·g)和157.20 U/(h·g),分别是CK 组的1.97 和3.00 倍。各处理中,以20 μmol/L MT 处理的PAL 活性最高。贮藏至第16 天时,20 μmol/L MT 处理组的PAL 活性比CK 高41.05%。

图7 MT 处理对接种链格孢菌冬枣果实PAL 活性的影响Fig.7 Effects of MT treatments on PAL activities in winter jujube inoculated with Alternaria alternata
由图8 可知,在贮藏期间,冬枣果实内CHI 活性在贮藏前期先快速增高而后降低,但下降的幅度不大,第12 天时又略有上升,之后缓慢下降。整个贮藏期间,2 μmol/L MT 和20 μmol/L MT 处理组CHI活性均高于CK 组。贮藏第14 天时,2 μmol/L MT 和20 μmol/L MT 处理组分别是CK 组的1.17 倍和1.56倍。贮藏第16 天,2、20、200 μmol/L MT 处理的CHI活性分别是CK 组的1.24、1.46、1.12 倍。由此可见,20 μmol/L MT 处理在提高果实CHI 活性方面的作用效果最好。
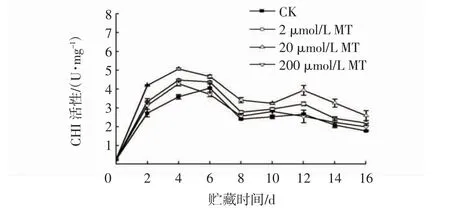
图8 MT 处理对接种链格孢菌冬枣果实CHI 活性的影响Fig.8 Effects of MT treatments on CHI activities in winter jujube inoculated with Alternaria alternata
由图9 可知,整个贮藏期间,MT 处理组冬枣果实的GLU 活性整体高于CK 组。MT 处理组和CK 组的GLU 活性均在第8 天到达峰值,此时20 μmol/L MT组的GLU 活性分别是2 μmol/L MT 组、200 μmol/L MT 组和CK 组的1.28、1.76、1.49 倍。贮藏第16 天时,20 μmol/L MT 处理的GLU 活性为CK 组的1.62倍。这说明MT 处理可以诱导GLU 活性的升高,提高冬枣果实对病原真菌的防御能力。
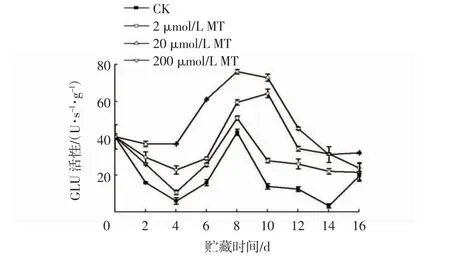
图9 MT 处理对接种链格孢菌冬枣果实GLU 活性的影响Fig.9 Effects of MT treatments on GLU activities in winter jujube inoculated with Alternaria alternata
由图10 可知,各处理冬枣果实中CAT 活性呈现先升高后降低的趋势,并且在第12 天达到峰值。在整个贮藏过程中,CK 处理的冬枣果实CAT 活性均低于各MT 处理组。20 μmol/L MT 处理对于冬枣果实中CAT 活性的提高有明显的促进作用。贮藏第12 天,20 μmol/L MT 处理冬枣果实中CAT 活性为11.28 U/g,是CK 组的1.86 倍;贮藏第16 天,20 μmol/L MT 处理冬枣果实中CAT 活性为5.90 U/g,是CK 组酶活性的2.03 倍。

图10 MT 处理对接种链格孢菌冬枣果实CAT 活性的影响Fig.10 Effects of MT treatments on the CAT activities in winter jujube inoculated with Alternaria alternata
由图11 可见,CK 组和MT 处理组的冬枣采后POD 活性在贮藏期间呈波浪式变化。POD 活性出现两次高峰,分别在第4 天和第8 天。在整个贮藏期间,20 μmol/L MT 处理组POD 活性均高于其他MT 处理组和CK 组,且与CK 组差异显著(P<0.05)。贮藏第16 天,CK 组的POD 活性高于200 μmol/L MT 组,20 μmol/L MT 处理组的POD 活性是CK 组的1.89倍。说明适宜浓度的MT 处理可以诱导冬枣中POD 活性的增强,其中20 μmol/L MT 处理效果最好。

图11 MT 处理对接种链格孢菌冬枣果实POD 活性的影响Fig.11 Effects of MT treatments on POD activities in winter jujube inoculated with Alternaria alternata
由图12 可知,整个贮藏期间,MT 处理的冬枣果实中黄酮含量均高于对照组,且随着贮藏时间的延长,MT 处理的冬枣果实中黄酮含量出现两个峰值,分别在第4天和第10 天。贮藏至第10天时,2、20、200 μmol/L MT 处理冬枣果实黄酮含量分别为CK组的1.15、1.56、1.12 倍。贮藏至第16 天,20 μmol/L MT处理组的黄酮含量比CK 组高27%。

图12 MT 处理对接种链格孢菌冬枣果实黄酮含量的影响Fig.12 Effects of MT treatments on flavonoids content in winter jujube inoculated with Alternaria alternata
由图13 可以看出,在整个贮藏过程中,CK 组冬枣的总酚含量在第6 天时出现高峰,之后下降,贮藏第8 天后又开始上升,MT 处理组的总酚含量分别在第6 天和第12 天出现两个峰值。与CK 相比,MT 处理提高了果实中的总酚含量。贮藏0~10 d,20 μmol/L MT 处理组的总酚含量由初值49.87 mg/100 g 升高至112.425 mg/100 g。贮藏第10 天时,2、20、200 μmol/L MT 组的总酚含量分别是CK 组的1.36、1.61、1.22倍。贮藏第16 天时,20 μmol/L MT 处理组的总酚含量比CK 组高31.25%。

图13 MT 处理对接种链格孢菌冬枣果实总酚含量的影响Fig.13 Effects of MT treatments on total polyphenol content in winter jujube inoculated with Alternaria alternata
由图14 可知,处理2d 后,2μmol/LMT 和20μmol/L MT 处理枣果实内的木质素含量呈先升高后降低的趋势,且在第8 天时达到峰值,而200 μmol/L MT 处理组和CK 组的木质素含量在整个贮藏期间一直保持较低水平。贮藏第16 天时,20 μmol/L MT 处理组的木质素含量分别是CK 组、2 μmol/L MT 处理组和200 μmol/L MT 处理组的5.13 倍、1.84 倍和3.28倍。

图14 MT 处理对接种链格孢菌冬枣果实木质素含量的影响Fig.14 Effects of MT treatments on lignin content in winter jujube inoculated with Alternaria alternata
3 讨论
冬枣在贮藏后期,随着果实的成熟衰老,其抗病性也逐渐降低,病斑直径扩大,自然腐烂指数也大大增加。在本试验中,适宜浓度的MT 处理可以有效抑制链格孢菌病斑的扩展,降低冬枣的腐烂指数,这与唐琦[12]的研究结果相似,MT 可以抑制青霉菌病斑扩展,降低冬枣的自然腐烂指数,增强枣果实抗病性。
膜完整性是生物体正常生理活动的基本保证。当膜脂肪酸被氧化时,细胞膜变得黏稠(更硬),会导致细胞的衰老。MDA 含量和细胞膜相对透性的升高,表明细胞膜的通透性变大,膜的完整性被破坏。CAT 作为清除自由基的关键酶,在保护膜的完整性中起重要作用。有研究表明,MT 处理可以保持较高的CAT、SOD 活性,降低梨果实的MDA 含量和相对电导率,抑制梨果实膜脂过氧化程度[24-25];MT 处理也可以通过增强CAT、SOD 活性来清除细胞中自由基保护膜的完整性,延缓果实衰老[17]。本试验结果表明,MT 处理可明显抑制冬枣果实的MDA 含量和相对电导率,提高枣果实CAT 活性,从侧面证实了MT 可以作为一种抗氧化剂,维持膜结构的完整性,延缓冬枣衰老,进而提高果实的抗病能力。
由于冬枣采后的水分蒸腾作用以及呼吸代谢对有机物质的消耗增加,果实逐渐萎缩,导致冬枣贮藏期间失重率也逐渐升高[19],果实的抗病性变差。在本研究中我们发现,MT 处理可以降低冬枣的失重率。前人的研究也表明,适宜浓度的MT 处理能够降低甜樱桃、苹果、柑橘、青花菜等果蔬的质量损失[4-5]。
据报道,水果的成熟和衰老与采后病原菌的传染特性有关[26]。在果实采后贮藏过程中,呼吸强度增大会导致果蔬的生理紊乱,促进营养成分的消耗,加速品质劣变,引起果实衰老,从而增加果蔬对病原微生物的敏感性。因此,控制果蔬的采后呼吸强度对减缓果蔬成熟,提高其抗病性是十分必要的[26]。已有研究表明,MT 处理可以降低甜樱桃[19]和红枣[28]的呼吸速率。在本试验中,枣果实在贮藏期间呼吸强度缓慢升高,但并未出现呼吸高峰,这说明冬枣是一种非呼吸跃变型果实。
苯丙烷途径与植物的抗病性密切相关[11]。大量研究发现,在植物受到逆境胁迫时,特别是生物胁迫时,PAL 活性呈现动态上升的趋势,达到高峰之后经过迟滞期然后又急剧下降。这与本研究结果一致,处理后的冬枣果实PAL 活性在第10 天时达到峰值之后开始下降,而枣果实内的次级代谢产物黄酮、总酚和木质素含量的变化趋势也与之相似,黄酮和总酚含量均在第10 天达到峰值,而木质素在第8 天达到峰值。大量的研究表明,POD 活性的变化在植物抵抗病原菌过程中起着多样性和广泛性的作用,其地位非常重要[28]。POD 参与植物细胞壁的建立过程,例如在对病原菌的抗病反应中,参与酚类化合物的氧化、亚甲基化和细胞木质化的过程,从而增强植物对真菌的抗攻击能力[17]。有研究提出,MT 可以参与苯丙烷代谢过程,上调相关基因的表达并通过增加其次级代谢产物酚类和木质素等植保素的积累来提高植物的抗病性。MT 处理也可以通过激活PAL、C4H、POD 的活性来调节苯丙烷代谢,提高番茄果实中总酚、黄酮和木质素的含量,增强果实对灰霉病的抗性[29]。MT 在酸枣、葡萄上的研究也有相同的结果,它提高了苯丙烷代谢相关基因表达水平,促进果实中黄酮、多酚含量的积累[7,29]。本研究结果也表明,MT 处理诱了导枣果实中苯丙烷代谢相关酶活性上升,同时提高了果实中提高抗病相关物质的含量,从而增强了果实对病原菌入侵的抗性。
CHI 和GLU 作为重要的病程相关蛋白,CHI 和GLU 活性与抗病性呈正相关[30-31]。CHI 在植物抗真菌病害中发挥重要作用,CHI 不仅可以通过水解真菌细胞壁的主要成分——几丁质,破坏病原菌细胞结构,还可以直接抑制真菌孢子萌发、菌丝生长和芽管伸长等发育活动[32]。GLU 不仅可以参与胼胝体的形成,堵塞筛板,对病原扩散有阻止作用,还可以降解真菌细胞壁中的β-1,3 葡聚糖,影响真菌的形态构建和生长发育,抑制其致病性[33-34]。之前的研究已经表明,这两种抗病蛋白均参与MT 对果实防御系统的诱导[28,30]。在本研究中,MT 诱导冬枣果实中的CHT 和GLU 活性的增强。因此,MT 可能通过诱导这些病程相关蛋白的活性从而增强枣果实的抗病性。
4 结论
本试验结果显示,适宜浓度的MT 处理可以抑制病斑直径的扩展,降低冬枣的自然腐烂指数,其中,以20 μmol/L MT 处理的效果最好;进一步的研究结果表明,MT 处理降低了MDA 含量和相对电导率的积累,维持了较为完整的果实细胞膜结构,延缓了冬枣的失重率和呼吸速率的升高,提高了冬枣果实中抗病相关酶活性和抗病相关物质的含量。因此,MT处理可诱导冬枣果实对由A.alternata 侵染引发的黑斑病产生抗性。
