基于系统发育的中国葡萄科植物多样性空间格局研究
2021-12-25张明罡司康杰赵夏云孙文静
张明罡,司康杰,赵夏云,孙文静
(山西大学 黄土高原研究所,山西 太原 030006)
0 引言
生物多样性空间格局研究在生物地理学、宏观生态学及保护生物学等多个学科中发挥着重要作用,是生态学研究的热点之一[1-2]。与生物多样性的形成过程相关的研究在保护策略制定[3]、应对气候变化[4]以及减缓人类活动干扰[5]等领域具有极其重要的生态价值。依据侧重点的不同,生物多样性格局的形成过程又被划分为生态学过程和历史过程两大类[6]。生态学过程关注的是物种与环境之间的关系,着重利用不同的气候组分对生物多样性空间格局进行解释。一方面,能量和水分的总量直接决定生态系统的净初级生产力[7],因此能够较好地解释为什么水热条件充足的地区有着较高的物种丰富度。另一方面,气候因子通过生态位对物种的分布产生影响,如温度的波动和干旱[8]将会增加物种的灭绝速率。在保护生物学实践中,良好的水热条件、稳定的生境也成为保护优先区选择的重要指标。
历史过程关注较长历史时期内物种的进化[9]。为了能更好地阐释高度多样化分类群的进化历史过程[10],新的概念被提出并应用于生物多样性空间格局研究,如Faith(1992)提出的系统发育多样性(phylogenetic diversity,PD)[11]以及 Rosauer(2009)等提出的系统发育特有性(phylogenetic endemism,PE)等指数[12]。PD和PE指数能更全面地反映生物多样性的质量和分布,在识别保护热点地区以及保护空缺分析方面也发挥了重要作用[13],很大程度上提高了相关研究的应用价值。结合物种的进化历史,为特有物种的现代格局增加时间维度以后,使得推断特定区域独特的进化历史和生物多样性的形成过程成为可能,如区分高度集中的短分支(新特有物种)和长分支(古特有物种)谱系的特有物种区域。新特有中心(neo-endemics),即最近分化的物种,由于他们没有从祖先地区扩散或迁移而成为特有性物种;古特有中心(paleo-endemics)是一种古老的分类群,在历史上分布更为广泛,但现在仅限于一个局部地区[14-15]。目前,这一方法已被应用于推断澳大利亚金合欢属植物多样性格局的形成过程[16],验证新世界蝙蝠类群[17]的起源和分化过程以及识别新西兰群岛[18]的生物地理区,为相关类群的生物多样性保护提供了新的视角。
葡萄科植物是重要的经济植物类群,在食用、药用、绿化、保健、工业等方面具有较大的开发利用价值。葡萄科植物共有14属900余种,在世界范围内广泛分布,但野生种多数分布于热带和亚热带地区[19]。中国共有8属146种,其中87种为中国特有种(含两个引入种)[19]。目前国内外对于葡萄科植物的研究主要集中在形态学、分子生态学、栽培、园艺等方面,对葡萄科空间分布格局及其保护的相关研究较少[20-22]。一直以来,具有经济价值的物种是系统发育研究的重要目标,通过以分布于中国行政区域内的葡萄科植物为研究对象,探讨其生物多样性格局以及形成机制,对于理解相应类群的生物地理学、适应和多样化具有重要意义,并且有助于保护和开发有价值的遗传资源[20,23]。
文章具体研究内容如下:(1)结合气候因子和物种分布信息,利用物种分布模型模拟每个物种的潜在分布区;(2)基于物种潜在分布区获取生物多样性空间格局地图,包括物种丰富度、加权特有性、系统发育多样性和系统发育特有性;(3)分析影响葡萄科植物多样性空间格局形成的环境驱动因子;(4)通过对葡萄科植物特有中心进行分类,进一步推断葡萄科植物多样性空间格局的形成过程,为葡萄科植物的利用与保护提供科学依据。
1 方法
1.1 数据获取
本文从国家标本资源共享平台(NSII,<http://www.nsii.org.cn>)中选取中国葡萄科植物的原始标本记录,该数据库是目前中国最大、最完整的植物分布数据库。执行以下步骤以确保数据质量:首先,植物学名通过中国植物志网络版(http://frps.iplant.cn/)和R包plantlist进行核对[19,24]。其次,由于大部分标本记录缺失经纬度信息,因此结合高精度的电子地图和标本采集标签上的地点描述,为标本记录逐一匹配经纬度信息。第三,在10’(arc-minute)的空间单元中确定物种的存在信息,避免在每个栅格内重复物种记录。在物种分布模型中,由于样本量小的物种无法提供足够的统计信息[25],因此将小于5个栅格的物种从分析中剔除,最终通过筛选得到了详细且准确的13 632条记录,包含96个物种,用于模拟研究。
在世界气候数据库(www.worldclim.org)中选择19个生物气候预测因子进行物种分布建模。利用ArcGIS 10.2将所有环境层重采样到精度为10’的空间单元中。然后使用Spearman秩相关分析来选择相关性最低的变量(Spearman’srho<0.75)。目的是避免多共线性导致物种分布模型的过拟合。在Spearmanrho大于0.75的相关变量中,仅保留生态学中最有意义的因子[26]。
1.2 物种分布模型和系统发育树的构建
物种分布建模算法选取最大熵模型MaxEnt 3.3.3k(http://biodiversityinformatics.amnh.org/open_source/maxent/)对中国葡萄科植物现代潜在分布区进行预测。MaxEnt运行遵循的建模规则:(1)物种采集记录为5~9条时,采用线性特征;(2)物种采集记录为10~14条时,采用二次特征;(3)物种采集记录≥15条时,采用复合特征。MaxEnt生态位模型是通过采用受试者工作特征曲线(receiver operator characteristic curves,ROC curves)下的面积(AUC值)作为SDMs(Species Distribution Models)预测准确度的指标,AUC的取值范围为0~1,数值越大表示模型的预测准确度越高。由于AUC值为非阈值依赖型指数,所以成为当前诊断试验中应用最为广泛的评价指标[27-28]。在本研究中,当AUC值>0.75,则认为该模型精度满足要求。
选取四个叶绿体DNA序列(trnC-petN、atpB-rbcL、rps16、psbA)和内转录间隔区(internal transcribed spacer,ITS)进行系统发育树的构建:葡萄科植物的基因数据主要来自GenBank(www.ncbi.nlm.nih.gov/)中下载相关基因序列,将所有的序列在Clustal X 2.1、BioEdit中进行序列比对和手动调整。碱基替代模型为根据PhySuite v1.2.1软件中ModelFinder计算出的GTR+F模型。最后,在PhyloSuite v1.2.1贝叶斯法构建系统发育树。所得的系统发育树在iTOL(https://itol.embl.de/)中进行编辑(图1)。
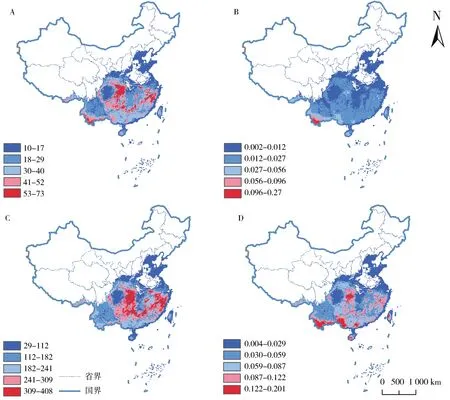
图1 中国葡萄科植物多样性分布格局图。(A)物种丰富度,(B)物种加权特有性,(C)系统发育多样性,(D)系统发育特有性。(阿伯斯投影)Fig.1 Spatial pattern of biodiversity of grape family(Vitaceae)in China,(A)species richness;(B)weighted endemism;(C)phylogenetic diversity;(D)phylogenetic endemism.(Albers Projection)
1.3 生物多样性指数的计算与相关性分析
将构建的中国葡萄科系统发育树与物种的潜在分布数据相结合,计算:(1)物种丰富度(species richness,SR):区域内所有物种的总和;(2)加权特有性(weighted endemism,WE):根据空间范围分布设置权重,即其分布区面积(所覆盖栅格数量)的倒数;(3)系统发育多样性(phylogenetic diversity,PD):区域内所有分类群的短路径支长总和,反映了物种组成的系统进化特征多样性;(4)系统发育特有性(phylogenetic diversity,PE):区域内每个物种范围比例加权的长度总和,反映了分类群在系统进化树上的离散程度。采用自然间断点分级法(Jenks’natural breaks)将每个指数划分为5个等级,绘制出葡萄科植物在中国的物种丰富度(SR)、物种加权特有性(WE)、系统发育多样性(PD)和系统发育特有性(PE)的4个空间分布格局图。由于小样本物种的缺失,而且由模型获取的潜在分布区存在一定的偏差,丰富度数值较低的区域通常存在极大的偏差率,因此物种数量在10以下的区域未予考虑,上述指数在Biodiverse 3.0软件中进行计算,在ArcGIS 10.2中生成地图。
采用最小二乘法(ordinary least squares,OLS)回归与空间自回归(spatial error simultaneous autoregressive,SAR)模型两种方法分析分别建立生物多样性格局指数与各气候因子之间的关系模型[29,30]。最小二乘法只关注变量之间的线性关系,而在生物多样性空间分析中,物种间的关系通常并不相互独立,因为种间竞争和环境的结合作用,导致种间存在空间自相关,因此另外建立了空间自回归模型。SAR模型是基于空间自相关过程发生在模型因变量、模型解释变量及模型误差的可能性而构建的,因此是最为常用的空间自相关模型,以上模型利用宏观生态学软件SAM 4.0进行构建。
1.4 新、古特有中心分类分析
新、古特有中心分类分析法(categorical analysis of neo-and paleo-endemism,CANAPE)是根据区域内受空间限制的分支长度,在系统发育和空间层次上对其进行分类,寻找物种的特有中心[5]。该方法可以识别出4种类型的特有中心:新特有中心、古特有中心、混合特有中心、超级混合特有中心。为了发现和区分不同类型的特有性中心,使用CANAPE进行分类:首先,确定一个地方是否为特有中心,栅格的相对系统发育特有性(RPE)的分子、分母或两者都需要显著高(单尾检验,P=0.05)。RPE是比较实际树的PE与比较树(保留了实际树的拓扑结构,但使所有分支的长度相等)的PE的比值。当栅格确定为特有中心后,它们会被分为三个有意义的、不重叠的特有中心。如果栅格RPE比率显著高或显著低(双尾检验,P=0.05),则为古特有中心或新特有中心;RPE不显著的栅格,则是混合特有性中心。混合特有中心说明它混合了稀有的长枝和稀有的短枝,因此没有显著地被古特有中心或新特有中心所主导。最后,进一步细分混合特有中心,分子和分母均显著高于P=0.01水平的栅格称为超级混合特有中心。新、古特有中心分类使用Biodiverse 3.0软件进行分析。
2 结果
对环境因子进行相关分析后,共有6个环境因子用于作物种分布建模(表S1)。在葡萄科植物中共有96个物种能够提供充分的统计信息满足物种分布建模的要求,并且能利用其在Genbank中的基因片段进行系统发育树的构建(表S2)。在对96个物种潜在分布区图层进行叠加以后,获取了中国葡萄科植物多样性分布格局地图(图1),分别是:物种丰富度(图1 A);物种加权特有性(图1 B);系统发育多样性(图1 C)和系统发育特有性(图1 D)。物种丰富度最高的地区为云南南部、秦岭及其南部山区、福建和浙江沿海区域,物种数量最高可达73种。在云南南部物种加权特有性显示为最高。系统发育多样性最高的区域为秦岭南部,湖南、湖北和重庆交界处,浙江和福建交界的东南沿海地区。系统发育特有性与物种丰富度有着相似的格局,但系统发育多样性在云南南部数值较低。系统发育特有性在云南南部,广西西部和湖北西部数值最高。
分别采用单变量最小二乘线性回归模型和SAR对生物多样性格局指数进行预测(表1),结果表明:在最小二乘线性回归模型中年平均温度和年降雨量对物种丰富度解释率最高,解释率分别达到62.8%和59.4%,且均与物种丰富度呈正相关,在增加空间权重后,等温性(负相关)和年降水量(正相关)对物种丰富度解释率最高,分别为82%和80%。对于物种加权特有性,使用最小二乘法回归,温度的季节性变化解释率最高(64.1%),并且呈负相关,在增加空间权重以后,年降水量(正相关)和季节性降水(负相关)解释率为最高,分别为77.4%和77.1%。在最小二乘线性回归模型中年降水量对系统发育多样性的解释率最高(55.1%),呈正相关,在增加空间权重以后,气温平均日较差和等温性解释率最高(88.4%),且均与系统发育多样性呈负相关。对于系统发育特有性,最小二乘法回归表明年降水量解释率最高(58.7%),且呈正相关,在增加空间权重以后,气温平均日较差解释率为最高(86%),呈负相关。
通过对相对系统发育特有性、实际树的系统发育特有性以及比较树的系统发育特有性进行显著性检验,将葡萄科植物特有中心划分为4类(图2)。秦岭南部山区、云南南部,广西西部、广东西部以及台湾部分地区的相对系统发育特有性比率显著性较低(P<0.05),显示为系统发育树上短枝聚集的区域,即区域内包含大量新分化的物种,因此被判定为新特有中心。古特有中心出现在云南南部和海南部分区域的相对系统发育特有性比率显著性较高(P>0.95),显示为系统发育树上长枝聚集的区域,即区域内含有大量进化时间较长的物种,因此被判定为古特有中心。混合特有中心和超级混合特有中心表现为混合交替出现,系统发育树上的稀有的长枝和稀有的短枝交替出现,此类区域包括:秦岭、横断山区中部的三江并流地区以及湖南、重庆和贵州省交界处。
2 讨论
葡萄科作为重要的经济植物类群,长期以来受到广泛关注[20,31]。虽然其人工栽培范围较广,但野生种仍多分布在热带和亚热带地区,在我国东北、华北和西北等各省区野生种类较少[19]。在本研究模拟的物种丰富度地图(图1 A)中,葡萄科植物在我国东北和西北区域内物种数均低于10种,物种多样性中心集中于热带和亚热带地区,但在此区域内纬度梯度并不明显,推测与该区域内充足的水热条件有关。物种多样性最高的区域出现在东南沿海、云南南部、秦岭包括南部的金佛山、大巴山、武陵山和神农架等地区。这些区域也是我国生物多样性研究的热点区域,如:云南南部是我国生物多样性最丰富的地区之一,热带生态系统保存最为完整;秦岭及其南部山区植被类型以亚热带常绿落叶阔叶混交林为主[32],保存有大面积的原始天然林和次生林,是中国种子植物“川东—鄂西特有现象中心”的核心区域[33-34]。特有中心的分布则与丰富度格局有所区别,多集中于云南南部的热带或其周边区域,表明该区域内的葡萄科植物多呈狭域分布。系统发育多样性与物种丰富度格局较为类似,但在云南南部地区数值较低,表明该区域内由新分化出的物种组成。
与最小二乘法回归模型相比较,在引入增加距离权重的空间自回归模型后,单变量对生物多样性格局指数的解释率均有所提高,表明各空间格局指数存在较强的空间自相关关系。在生物多样性格局指数的驱动因子中,年降水量和与气温稳定性相关的因子(等温性、季节性温度以及气温日较差)显示出较强的解释能力。在解释生物多样性空间格局的水-热动态假说中,低纬度地区的限制因子为水分,与本研究的结果具有一致性,但能量的限制在高纬度地区目前无法用本植物类群得到证明。气候的稳定性对生物多样性格局有两个方面的影响,葡萄科植物中仅有葡萄属(Vitis)、地锦属(Parthenocissus)和蛇葡萄属(Ampelopsis)的少数种分布在温带,在向北扩散迁移的过程中,极端低温限制了植物的生长,尤其是葡萄科植物带有明显的热带属性。另外,不稳定的气候会增加物种的灭绝速率,因此生物多样性格局指数较高的区域与气候稳定的区域相一致。由此可以做出如下推断:当未来气候发生波动时,降水量的减少以及气候的不稳定性会增加物种的本地灭绝速率。
川东-鄂西、川西-滇西北、滇东南-桂西是中国种子植物特属的三大分布中心[35-36]。葡萄科植物的特有中心与中国种子植物特有属的分布中心均有所重合。新特有中心的集中区域可分为两种类型,一类是以云南南部、广西西部、广东南部沿海和台湾区域为代表的热带,及秦岭及其南部山区。植物种在热带有着较快的分化速率已在多项研究中得到证明,相关结论适用于葡萄科植物类群。秦岭及其南部山区有着较高的地形复杂度,地理因素造成的种间隔离促进了物种的分化,因此本区域内以新物种的分化过程为主,因此系统发育树上的短枝较为集中。云南南部和海南的特有种系统发育较为古老,推测属于古地质时期古老种类自然分布区缩小残留形成,同时这两个区域热带特征也较为明显。混合特有中心和超级混合特有中心出现在川西-滇西北和川东-鄂西两个大的区域,这两个区域内同时存在物种分化和古老分布区的残留。川西-滇西北位于横断山区的高山地带,由于特殊的地理特征,本区域成为许多原生植物在更新世冰期的“避难所”,有着大量的古老植物的遗存。鄂西区域是东亚亚热带植物区系华中区系成分分布的核心地带,受第四季冰川的影响较小,因此也有较多的古老孓遗物种。除古老物种的遗存以外,川西-滇西北和川东-鄂西也有着较高的地形复杂度,以川西-滇西北为例,该区域内的植物类群由于地理隔离以及适应高寒和干旱的自然环境,从而成为许多新的植物种属的分化中心。因此,混合特有中心和超级混合特有中心同时具备葡萄科植物“摇篮”和“博物馆”的作用[36]。
3 结论
在全国尺度上,已有大量研究对木本植物、特有植物以及珍稀濒危植物的多样性格局进行分析[36-37],以特定的植物类群为对象的研究能够更有针对性的对宏观保护策略进行补充[38]。当前,葡萄科植物分布数据以及分子数据的积累已相对完善,具备开展大尺度研究的基础。国家标本资源共享平台中的标本数据在其收录的标本馆中都能根据其采集号与实物相对应,确保其分布地点真实可靠。物种分布模型能够结合气候因子对未采样区域进行补充,为生物多样性空间格局指数的获取提供了便利。生物多样性空间格局指数高低直观地反映了相应区域的重要程度,在保护生物学实践中被广泛应用。在各指数的基础上,本研究特别指出了年降水量和气候稳定性两类环境因子对葡萄科植物的分布起到决定性作用,也是气候变化背景下应重点关注的方向。最后,对葡萄科植物特有中心格局进行了分析,明确了该类群的分化中心以及古老分布区的残留等格局。上述结果将有助于该植物类群的保护以及开发有价值的遗传资源。
