miR-31-5p通过VPS53介导的自噬对结直肠癌细胞增殖的影响*
2021-11-24黎爽彭洪龚磊唐军伟吴洪黄梅
黎爽,彭洪△,龚磊,唐军伟,吴洪,黄梅
1 南充市中心医院肛肠外科 四川南充 637000
2 南充市中心医院胃肠外科 四川南充 637000
3 南充市中心医院中西医结合科 四川南充 637000
结直肠癌是一种常见的恶性肿瘤,近年来结直肠癌的发病率呈上升趋势,尤其是在亚洲国家[1]。随着医疗技术的进步,结直肠癌的治疗手段越来越多,但是结直肠癌患者的总体预后依然有待改善,尤其是出现淋巴结转移或远处转移的患者。因此,需要进一步探究结直肠癌发生发展的潜在作用机制,以期为结直肠癌患者治疗方案的制定提供新的思路[2]。
MicroRNA(miRNA)是一类非编码RNA,广泛存在于多细胞生物和真核生物中,参与细胞的多种生理过程,包括基因表达的转录后调控。miRNA可调控肿瘤细胞的多种生物学行为,包括增殖、迁移和凋亡[3]。miRNA的异常表达影响着结直肠癌的发生发展,提示miRNA有可能作为结直肠癌新的治疗靶点[4]。
miR-31-5p由miR-31的5’端臂转录加工而成,在多种细胞和组织中表达,在细胞增殖、分化及维持细胞稳态等方面发挥重要作用[5]。既往研究表明miR-31-5p在多种恶性肿瘤中高表达[6-8],我们前期的研究也发现miR-31-5p在结直肠癌组织中的表达水平明显高于正常结直肠黏膜组织[9],然而miR-31-5p在结直肠癌发生发展中的作用机制尚不明确。通过生物信息学数据库,我们发现miR-31-5p可与VPS53靶向结合,进一步通过GEPIA数据库,我们发现VPS53与自噬相关基因Beclin1的表达呈正相关。自噬是指细胞通过降解自身结构物质使细胞存活的自我保护机制,在肿瘤中自噬可发挥抑癌作用[10]。Beclin1基因(编码Beclin1蛋白)是自噬通路的关键调控基因[11],但miR-31-5p是否通过调控自噬通路及Beclin1的表达参与结直肠癌的发生发展尚不清楚。为此,本研究探讨miR-31-5p通过VPS53介导的自噬对结直肠癌细胞增殖的影响,现报告如下。
1 材料与方法
1.1 实验细胞
人正常结直肠黏膜细胞(FHC)与结直肠癌细胞株HT29、HCT116、SW480、LoVo均购自美国种质保藏中心(American Type Culture Collection,ATCC)。
1.2 细胞培养
HT29、HCT116、SW480、LoVo细胞在含10%胎牛血清、1%青霉素(100 μg/mL)和1%链霉素(100 μg/mL)的DMEM培养基中进行培养。细胞置于37℃、含5% CO2的恒温培养箱中培养。
1.3 实时荧光定量PCR(qPCR)实验
用 Trizol试剂(Invitrogen,Carlsbad,CA)从FHC细胞及结直肠癌HT29、HCT116、SW480、LoVo细胞株中提取总RNA,然后用逆转录试剂盒(Taka⁃ra,Japan)逆转录为cDNA,用SYBR Green PCR试剂盒(Invitrogen,USA)在ABI stepone plus型PCR仪(Applied Biosystems,Foster City,CA)上进行PCR扩增。PCR反应条件为:95℃2 min,然后94℃ 20 s,58℃ 20 s,72℃ 20 s,共进行40个循环,72℃5 min。再用Stepone plus Real-Time PCR系统(Applied Biosystems,CA)进行检测。采用2-ΔΔCt法计算目的基因的相对表达量。
(1)miR-31-5p的表达水平。miR-31-5p引物如下:上游5’-ACACTCCAGCTGGGAGGCAAGAT⁃GCTGGCATA-3’,下游 5’-CTCAACTGGTGTCGTG⁃GAGTCGGCAATTCAGTTGAGAGCTATG-3’。U6引物如下:上游 5’-CTCGCTTCGGCAGCACA-3’,下游5’-AACGCTTCACGAATTTGCGT-3’。根据本实验结果及查阅相关文献资料[12],本研究选择LoVo细胞进行后续实验。
(2)VPS53的表达水平。分别转染miR-31-5p模拟物(mimics)、miR-31-5p阴性对照(NC)和不作任何处理,分为miR-31-5p过表达组、miR-31-5p阴性对照组和空白对照组。VPS53引物序列如下:上游5’-TGTTCCCAACCGAGCAATCTC-3’,下游5’-ACGTTCGTCTGACCTCTTACA-3’。GAPDH 引物如下:上游5’-TGTTCGTCATGGGTGTGAAC-3’,下游5’-ATGGCATGGACTGTGGTCAT-3’。
(3)Beclin1的表达水平。分别转染miR-31-5p模拟物(mimics)、miR-31-5p阴性对照(NC)、miR-31-5p mimics+pcDNA-VPS53和不作任何处理,分为miR-31-5p过表达组、miR-31-5p阴性对照组、miR-31-5p/VPS53过表达组和空白对照组。Beclin1引物序列如下:上游5’-AGGTTGAGA⁃AAGGCGAGACA-3’, 下 游 5’-GTCCACTGCTCCT⁃CAGAGTT-3’。GAPDH引物如下:上游5’-GGGCT⁃GCCTTCTCTTGTGAC-3’,下游5’-CCCGTTGATGAC⁃CAGCTTCC-3’。
1.4 CCK- 8法检测细胞活力
将LoVo细胞(1×104/孔)接种在96孔板中培养24 h(37 ℃,5% CO2)。
(1)当细胞密度达80%时,分别转染miR-31-5p模拟物(mimics)、miR-31-5p阴性对照(NC)和不作任何处理,分为miR-31-5p过表达组、miR-31-5p阴性对照组和空白对照组。
(2)构建VPS53过表达载体,用PrimerSTAR Max DNA聚合酶复合物(Takara,Japan)扩增全长VPS53基因,并将其克隆到pcDNA3.1+载体(Invitro⁃gen)。当细胞密度达80%时,分别转染pcDNAVPS53、pcDNA-VPS53阴性对照(NC)和不作任何处理,分为VPS53过表达组、VPS53阴性对照组和空白对照组。
(3)当细胞密度达80%时,分别转染miR-31-5p模拟物(mimics)、miR-31-5p阴性对照(NC)、miR-31-5p mimics+pcDNA-VPS53和不作任何处理,分为miR-31-5p过表达组、miR-31-5p阴性对照组、miR-31-5p/VPS53过表达组和空白对照组。
(4)当细胞密度达80%时,分别转染pcDNAVPS53、pcDNA-VPS53阴性对照(NC)、pcDNAVPS53+3-MA和不作任何处理,其中3-MA为自噬抑制剂,分为VPS53过表达组、VPS53阴性对照组、VPS53过表达联合自噬抑制剂组和空白对照组。
每组设4个复孔,分别转染24 h、48 h、72 h后,每孔加入15 μL CCK-8试剂,孵育3 h后在450 nm处用酶标仪(Thermo Fisher)测量OD值。
1.5 生物信息学分析
(1)利用数据库 Targetscan(http://www.tar⁃getscan.org/vert_71/)进行生物信息学分析,筛选与miR-31-5p靶向结合的靶基因,并通过双荧光素酶报告实验进行验证。
(2)利用数据库GEPIA(http://gepia.cancerpku.cn/detail.php)分析VPS53和Beclin1(自噬通路关键基因)表达的相关性。
1.6 双荧光素酶报告实验
荧光素酶报告基因质粒pcDNA-VPS53-野生型(WT)或pcDNA-VPS53-突变型(MUT,突变位点为UCUUGCC)由Generay(上海)设计和构建。使用Lipofectamine™3000转染试剂在LoVo细胞内转染荧光素酶报告基因质粒,并共转染miR-31-5p模拟物(mimics)或miR-31-5p阴性对照(NC)。用双荧光素酶报告系统检测荧光素酶活性。
1.7 透射电子显微镜实验
将LoVo细胞(1×107/孔)接种在6孔板中培养24 h(37℃,5% CO2),细胞铺满70%左右时,分别转染pcDNA-VPS53、pcDNA-VPS53阴性对照(NC)和不做任何处理,分为VPS53过表达组、VPS53阴性对照组和空白对照组,继续培养24 h,收集LoVo细胞并进行离心(500 g,5 min)。在细胞沉淀中加入4℃条件下预冷的5%戊二醛溶液混合5 min。悬浮并离心细胞,将沉淀用5%戊二醛和10%牛血清白蛋白(BSA)在冰上处理4 h。洗涤细胞后用8%焦磷酸钾和1%氧化四氧化锇固定细胞。细胞用丙酮脱水,再用环氧树脂浸泡2 d。使用徕卡超薄切片机(EM UC7)进行切片,用2%醋酸铀酰染色后使用透射电子显微镜进行观察。
1.8 蛋白免疫印迹实验(Western blot)
将LoVo细胞(1×107/孔)接种在6孔板中培养24 h(37℃,5% CO2),细胞铺满70%左右时,分别转染pcDNA-VPS53、pcDNA-VPS53阴性对照(NC)和不做任何处理,分为VPS53过表达组、VPS53阴性对照组和空白对照组,继续培养24 h后弃去6孔板中的培养基,将6孔板置于冰上,用1 mL PBS重悬细胞,4℃13 000 rpm离心5 min后弃去上清,以除去样本中血清。加入500 μL RIPA裂解液(Beyo⁃time)置冰上裂解10 min并振荡3次,4℃13 000 rpm离心10 min后将上清转移至离心管中。用SDS-PAGE分离蛋白,蛋白上样量为30 μg,将蛋白转移至PVDF膜(Bio-Rad)。用5%脱脂牛奶封闭1 h,洗膜,加入一抗在4℃条件下孵育过夜。洗膜,加入二抗(Santa Cruz)孵育40 min,洗膜,取适量发光液置于暗盒中孵育5 min,用凝胶成像系统曝光并拍照。实验中用到的一抗有:anti-GAPDH抗体(Abcam,ab9485,1:3 000)、anti-Beclin1抗体(Abcam,ab62557,1:500)。
1.9 统计学分析
使用GraphPad 8.0进行数据分析并绘图,两组间比较采用t检验。以P<0.05为差异有统计学意义。
2 结果
2.1 miR-31-5p在结直肠癌细胞中高表达
与FHC细胞相比,HCT116细胞、LoVo细胞、HT29细胞、SW480细胞的miR-31-5p表达水平均升高(均P<0.001),见图1。我们选择LoVo细胞进行后续实验。

图1 miR-31-5p在结直肠癌细胞中高表达
2.2 过表达miR-31-5p促进细胞增殖
转染24 h,miR-31-5p过表达组细胞活力分别与阴性对照组、空白对照组比较,差异均无统计学意义(均P>0.05)。转染48 h、72 h,miR-31-5p过表达组细胞活力高于其余两组(均P<0.05)。见图2。
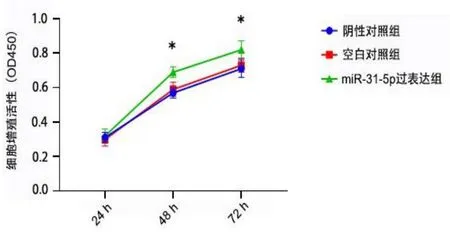
图2 过表达miR-31-5p促进细胞增殖
2.3 miR-31-5p与VPS53靶向结合
通过Targetscan数据库,我们发现miR-31-5p含有VPS53的结合位点,进一步通过双荧光素酶报告实验证实miR-31-5p与VPS53靶向结合(图3、图4)。

图3 miR-31-5p与VPS53靶向结合
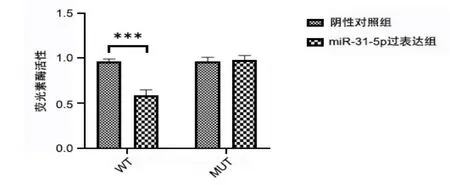
图4 双荧光素酶报告实验
2.4 过表达miR-31-5p下调VPS53的RNA表达水平
与阴性对照组、空白对照组相比,miR-31-5p过表达组VPS53的RNA表达水平均降低(均P<0.001)。见图5。
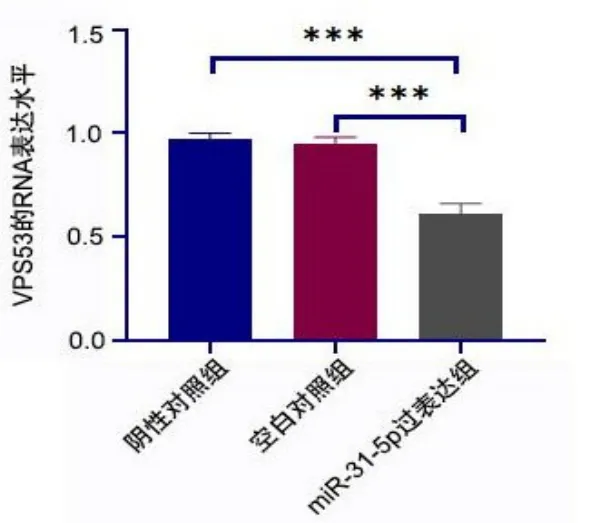
图5 过表达miR-31-5p下调VPS53的RNA表达水平
2.5 过表达VPS53抑制细胞增殖
转染24 h,VPS53过表达组细胞活力分别与阴性对照组、空白对照组比较,差异均无统计学意义(均P>0.05)。转染48 h、72 h,VPS53过表达组细胞活力低于其余两组(均P<0.05)。见图6。

图6 过表达VPS53抑制细胞增殖
2.6 VPS53逆转miR-31-5p对细胞增殖的促进作用
转染24 h,miR-31-5p过表达组细胞活力分别与空白对照组、阴性对照组、miR-31-5p/VPS53过表达组比较,差异均无统计学意义(均P>0.05)。转染48 h、72 h,miR-31-5p过表达组细胞活力高于空白对照组、阴性对照组,miR-31-5p/VPS53过表达组细胞活力低于miR-31-5p过表达组(均P<0.05)。见图7。

图7 VPS53逆转miR-31-5p对细胞增殖的促进作用
2.7 过表达VPS53促进自噬小体的形成
空白对照组、阴性对照组、VPS53过表达组各取3张片,每张片随机取一个图进行观察(如图8)。与空白对照组、阴性对照组相比,VPS53过表达组自噬小体数量增多,差异均有统计学意义(均P<0.05),见图9。
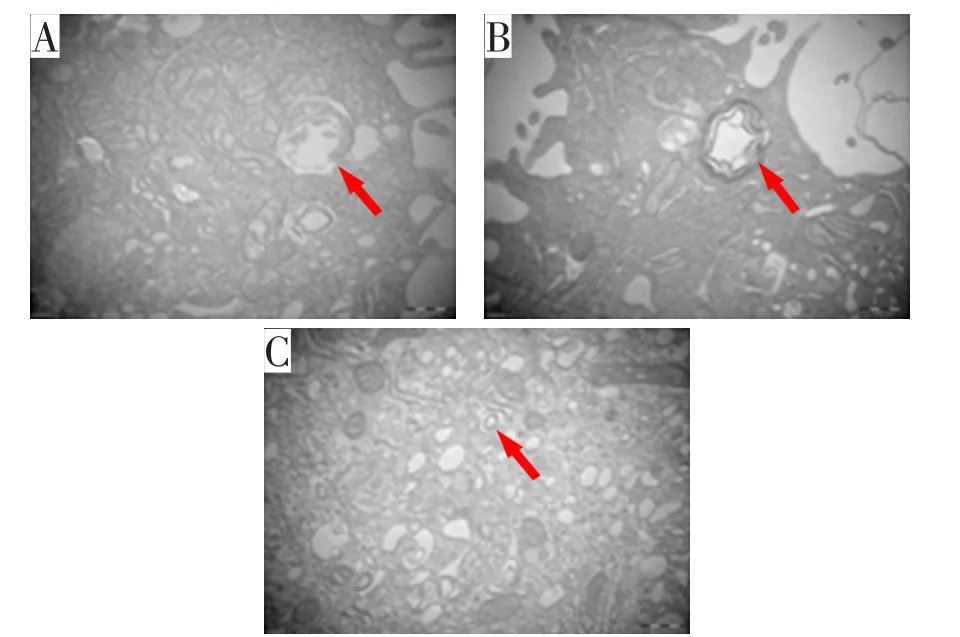
图8 透射电子显微镜下自噬小体图(×40 000)

图9 过表达VPS53促进自噬小体的形成
2.8 VPS53和Beclin 1的表达呈正相关
通过GEPIA数据库,我们发现结直肠癌组织中的VPS53和自噬相关基因Beclin1的表达呈正相关(图10)。

图10 VPS53和Beclin 1的表达呈正相关
2.9 VPS53上调Beclin 1蛋白的表达
Western blot结果见图11。灰度分析采用Image J,与空白对照组、阴性对照组相比,VPS53过表达组Be⁃clin1蛋白表达水平均升高(均P<0.001),见图12。

图11 Western blot结果
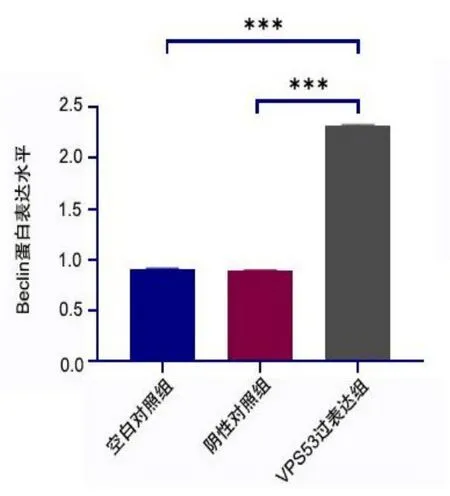
图12 VPS53上调Beclin 1蛋白的表达
2.10 自噬抑制剂逆转VPS53对细胞增殖的抑制作用
转染24 h,VPS53过表达组细胞活力分别与空白对照组、阴性对照组、VPS53过表达联合自噬抑制剂组比较,差异均无统计学意义(均P>0.05)。转染48 h、72 h,VPS53过表达组细胞活力低于空白对照组、阴性对照组,VPS53过表达联合自噬抑制剂组细胞活力高于VPS53过表达组(均P<0.05)。见图13。
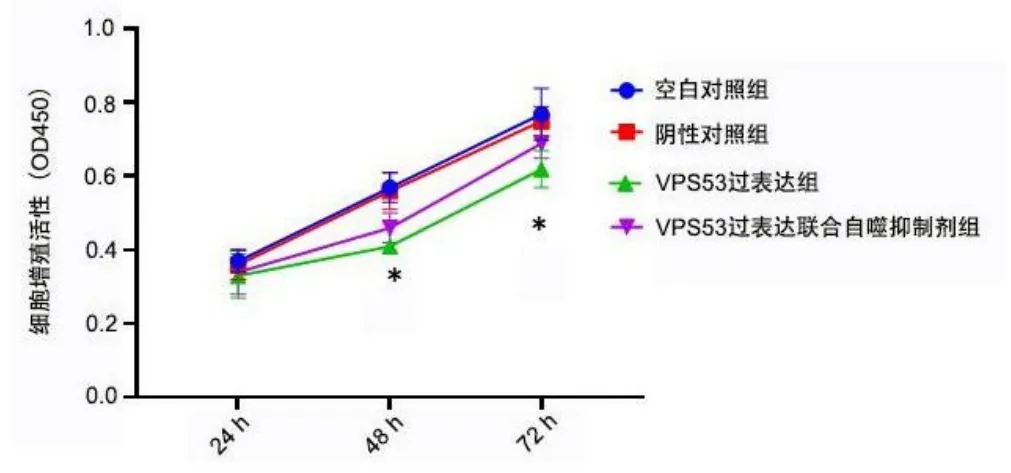
图13 自噬抑制剂逆转VPS53对细胞增殖的抑制作用
2.11 VPS53逆转miR-31-5p对Beclin 1 RNA表达的抑制作用
miR-31-5p过表达组Beclin1的RNA表达水平低于空白对照组、阴性对照组,miR-31-5p/VPS53过表达组Beclin1的RNA表达水平高于miR-31-5p过表达组(均P<0.05)。见图14。

图14 VPS53逆转miR-31-5p对Beclin 1 RNA表达的抑制作用
3 讨论
miR-31-5p位于染色体9p21.3上,由单基因座编码。目前miR-31-5p的研究主要集中在其调控肿瘤增殖、转移和侵袭等方面,有研究发现miR-31-5p在食管癌、肾癌和子宫内膜癌等多种恶性肿瘤中的表达异常并与肿瘤的发生发展有关[13-15]。既往研究显示miR-31-5p在结直肠癌组织中表达上调,并与结直肠癌的分化程度、浸润深度及预后有关[16]。我们本次研究也发现miR-31-5p在结直肠癌细胞中的表达水平较人正常结直肠黏膜细胞升高,并且过表达miR-31-5p可促进结直肠癌细胞的增殖,提示miR-31-5p可能在结直肠癌的发生发展中扮演着致癌基因的角色,未来有可能成为一个新的结直肠癌标志物,但目前miR-31-5p在结直肠癌发生发展中的作用机制尚未明确。
miRNA主要通过与其靶基因序列特异性结合,在转录后水平负调控靶基因的表达[17]。为探讨miR-31-5p促进结直肠癌细胞增殖的潜在机制,我们利用Tar⁃getscan数据库对miR-31-5p的靶基因进行预测,发现miR-31-5p含有VPS53的结合位点,并通过双荧光素酶报告实验证实VPS53与miR-31-5p靶向结合。进一步通过qPCR实验发现miR-31-5p可负调控VPS53的表达,符合miRNA与其靶基因的常规调控关系。
VPS53是高尔基相关逆向运输蛋白(Golgi-asso⁃ciated retrograde protein,GARP)复合体的一个亚单位,参与蛋白质从内质体向高尔基体的逆向转运,并加速多能干细胞的分化[18]。研究表明VPS53在多种肿瘤中表达下调,如甲状腺髓样癌和前列腺癌[19-20],并且VPS53还可诱导异位内膜间质细胞凋亡[21]。有研究报道VPS53在结直肠癌组织中低表达[22],我们的研究结果显示过表达VPS53可抑制结直肠癌细胞的增殖,进一步通过CCK-8法研究发现VPS53可逆转miR-31-5p对细胞增殖的促进作用,提示VPS53在结直肠癌中可能作为抑癌基因发挥作用。
自噬是一种高度保守的溶酶体依赖性降解途径,在维持细胞稳态方面发挥着重要作用[23]。自噬可以通过清除受损的细胞器和错误折叠的蛋白质,影响肿瘤的发生发展[24],在结直肠癌的发生发展过程中起着重要作用,并与结直肠癌的预后密切相关[25]。诱导自噬可促进结直肠癌细胞凋亡,抑制自噬可促进结直肠癌细胞增殖[26],并且激活肿瘤相关巨噬细胞的自噬可增强结直肠癌细胞的放疗敏感性[27]。因此,自噬相关通路可作为结直肠癌预防和治疗的思考方向。Beclin1蛋白是自噬通路的关键蛋白,在自噬起始阶段起到关键作用,并在细胞的生命活动中发挥着重要作用[28]。我们的研究发现过表达VPS53促进自噬小体的形成,且通过对GEPIA数据库进行分析,发现VPS53和Beclin1的表达呈正相关,然而VPS53是否通过调控自噬相关通路参与结直肠癌的发生发展尚未清楚。我们进一步的研究发现过表达VPS53可上调Beclin1蛋白的表达,而自噬抑制剂可以逆转VPS53对结直肠癌细胞增殖的抑制作用,这些结果提示VPS53可能通过调节自噬相关通路抑制结直肠癌的发生和发展。此外,我们的研究发现过表达miR-31-5p可下调Beclin1的RNA表达水平,同时过表达VPS53可逆转过表达miR-31-5p对Beclin1 RNA表达的抑制作用,提示miR-31-5p可能通过下调VPS53来抑制Beclin1的表达。
综上所述,本研究发现miR-31-5p是结直肠癌发生发展中的重要调控因子。miR-31-5p在结直肠癌细胞中高表达,miR-31-5p可能通过下调VPS53来抑制Beclin1的表达,进而通过抑制自噬来促进结直肠癌细胞的增殖。因此,miR-31-5p可能是一个新的结直肠癌治疗的潜在靶点。
