植物激素在大叶落地生根胎生苗无性繁殖过程中的作用
2021-11-20钟天秀
刘 华,钟天秀
(1.青海省草原总站,青海 西宁 810008;2.华南农业大学林学与风景园林学院草业科学系 广东省草业工程技术研究中心,广东 广州 510642)
落地生根(kalanchoedaigremontiana),原产马达加斯加,在我国主要分布在福建、台湾、广东、广西、云南等地,喜温暖潮湿的气候,宜种植在排水良好的肥沃砂质土壤上。落地生根在我国一直作为一种常见的中草药,用于治疗烫伤、急性中耳炎、热性胃痛等。落地生根叶片宽大肥厚,在叶片周围形成一圈类似蝴蝶的胎生苗,所以它也作为一种常见的园艺植物应用在生活中。落地生根的胎生苗不仅具有园艺观赏的价值,而且还是研究体细胞胚和器官繁殖的理想模型(H. et al. 2009)。我国目前对胎生苗的研究大部分集中对其形态解剖学进行研究(尹秀玲等. 2005;袁强 等. 2006 ),对胎生苗产生的原因及影响因素的研究较少。国外对胎生苗产生的研究也存在争议,Heide(Heide 1965)认为生长素强烈地抑制了落地生根叶片上胎生苗的产生。而其他实验却认为生长素的抑制作用很弱或者几乎不产生抑制作用(Karpoff 1982;Yazgan and Vardar 1977)。最近,Kulka(Kulka 2005)重新试验了各种植物激素对白姬之舞(Kalanchoemarnierianum)产生胎生苗的作用,认为细胞分裂素是为维持胎生苗原基休眠的关键激素。白姬之舞是属于胁迫诱导型产生胎生苗的物种之一,而大叶落地生根属于半自发产生胎生苗的类型。目前还没有研究探讨过植物激素对大叶落地生根胎生苗的作用。
所以,本文以大叶落地生根为植物材料,对影响胎生苗产生的激素进行了深入分析,探讨激素对胎生苗产生过程所扮演的角色,为进一步研究胎生苗提供理论依据和实践支持。
1 材料和方法
1.1 植物材料的准备
实验中所用的植物材料均是由同一株大叶落地生根繁殖而成,将大叶落地生根叶片上长出的胎生苗播种在装有草炭、蛭石和沙(按4∶2∶1混合)的育苗盘中于培养箱中培养,培养条件为14 h光照,10 h黑暗,光照度2 000 Lx。待胎生苗长到约5 cm时,取材待用。另将一部分长到约5 cm的落地生根幼苗,进行干旱处理,获得胎生苗。
1.2 植物激素的配置
2,4-D、6-BA、生长素抑制剂TIBA(2,3,5-triiodobenzoic acid)、细胞分裂素抑制剂PR (purine riboside)、乙醇、DMSO(二甲基亚砜)等试剂均购于科百奥公司。2,4-D溶于乙醇,配制为2 mg/mL的贮存液,放置于-20 ℃冰箱中待用。6-BA用DMSO溶解,配制为2 mg/mL的贮存液,放置于-20 ℃冰箱中待用。TIBA 在乙醇中溶解后,配制为10 μm/mL 的贮存液,放置于-20 ℃冰箱中待用。PR 溶于水后,配置为100 μm/mL的贮存液,放置于-20 ℃冰箱中待用。
1.3 实验处理
对大叶落地生根叶片、胎生苗进行表面消毒,依次放入70%的酒精中30 S,2%的次氯酸钠中10 min,并用无菌水清洗6次。叶片消毒完成之后,取其中一部分叶片切割成约为0.5 cm×0.5 cm的正方形小块。将3种植物外植体材料(完整叶片、切割叶片、胎生苗)放入培养板中,培养板中含MS培养基(Murashige and Skoog,1962)、8 g/L的琼脂、3 g/L的蔗糖及2 mg/L的2,4-D和6-BA或10 μm/L TIBA 以及250 μm/L PR。将培养板放入培养箱中,培养条件同上。
1.4 数据统计
本实验采用随机设计,记录不同处理下的胎生苗发生时间和发生数量,以及胎生苗的叶幅宽的变化情况。利用SPSS软件对数据进行单因素方差分析,P<0.05代表差异显著。
2 结果与分析
2.1 不同类型外植体在不同处理情况下诱导产生胎生苗的表型差异

图1 不同类型大叶落地生根外植体在不同处理情况下诱导产生的胎生苗的表型
A)完整叶片在(1)MS培养基上诱导产生的胎生苗表型;(2)MS培养基附加2,4-D (2 mg/L)诱导产生的胎生苗表型;(3)MS培养基附加 6-BA (2 mg/L)诱导产生的胎生苗表型;(4)MS 培养基同时附加 2,4-D (2 mg/L) 和 6-BA (2 mg/L)诱导产生的胎生苗表型; (5) MS 培养基附加 TIBA(10 μm/L) 诱导产生的胎生苗表型; (6)MS培养基附加 PR(10 μm/L)诱导产生的胎生苗表型; (7)MS培养基同时附加 2,4-D 和 PR 诱导产生的胎生苗表型; (8) MS培养基同时附加 6-BA 和 TIBA 诱导产生的胎生苗表型。B)切割叶片在(1)MS培养基上诱导产生的胎生苗表型;(2)MS培养基附加2,4-D(2 mg/L)诱导产生的胎生苗表型;(3)MS培养基附加 6-BA(2 mg/L)诱导产生的胎生苗表型;(4)MS 培养基同时附加 2,4-D(2 mg/L) 和 6-BA(2 mg/L)诱导产生的胎生苗表型; (5) MS 培养基附加 TIBA(10 μm/L) 诱导产生的胎生苗表型; (6)MS培养基附加 PR(10 μm/L)诱导产生的胎生苗表型; (7)MS培养基同时附加 2,4-D 和 PR 诱导产生的胎生苗表型; (8) MS培养基同时附加 6-BA 和 TIBA 诱导产生的胎生苗表型。
通常情况下,胎生苗的发生部位是叶片边缘的缺刻处,从叶片顶端向叶柄基部逐渐发生。生长素(2,4-D)诱导产生的胎生苗类似于愈伤组织,叶片发育不完全,许多叶片呈一个圆形,叶柄基部膨大,呈白色,出现类似愈伤组织白色透明的组织,仅有少量的能分化出根,大部分分化出的胎生苗就面临着死亡。细胞分裂素(6-BA)诱导产生的胎生苗,叶片与在对照中一样,发育完整。同时添加生长素和细胞分裂素,诱导产生的胎生苗与只添加生长素中的胎生苗生长情况类似,胎生苗下部膨大,而且叶片发育不完整,这说明在控制胎生苗表型上,细胞分裂素(6-BA)对生长素(2,4-D)的拮抗作用不明显。使用生长素抑制剂(TIBA)对完整叶片上诱导产生的胎生苗表型没有影响,但是使在切割叶片诱导的胎生苗表型出现叶片发育不完整和不对称的情况,这说明生长素抑制剂在切割叶片中制约相当一部分生长素的作用。细胞分裂素抑制剂(PR)对完整叶片上诱导产生的胎生苗表型没有影响,但是在切割叶片中,施用细胞分裂素抑制剂(PR)完全抑制了胎生苗的产生。同时使用生长素和细胞分裂素抑制剂,完整叶片上仍然能诱导胎生苗产生,而且不像单独施用生长素的胎生苗,这些胎生苗的发育情况较为完整。但是在切割叶片中,没有发现胎生苗的产生。同时施用细胞分裂素和生长素抑制剂(TIBA),在完整叶片和切割叶片上都能诱导产生发育完全的胎生苗。另外,我们把相同处理的落地生根放在黑暗情况下,发现没有光照这个条件,所有外植体都死亡,不能诱导产生胎生苗 (照片没有展示)。由于不定芽产生比较一致,不定根产生的时间比较复杂,不定根不一定从时间最久的不定芽上最先发生,有时在时间短的不定芽上反而先长出不定根。我们的研究集中在对不定芽发育影响的探讨上,激素对胎生苗不定根产生及发育的影响将会在以后的研究中进行叙述。
2.2 以完整叶片作为外植体在不同处理情况下诱导产生胎生苗的时间和数量差异

图2 以大叶落地生根完整叶片作为外植体在不同处理情况下诱导产生胎生苗的时间和数量差异
A)不同处理条件下,以大叶落地生根完整叶片为外植体诱导产生胎生苗的时间差异。B)不同处理条件下,以大叶落地生根完整叶片为外植体诱导产生胎生苗的数量差异。
6-BA和2,4-D都对胎生苗的产生有抑制作用。其中,使用2,4-D后,胎生苗的产生时间推迟2.8 h;2,4-D对胎生苗产生的抑制作用不太明显,使用TIBA后,胎生苗产生时间推迟2.4 h,对胎生苗产生时间的影响不是很大,说明生长素可能在控制胎生苗产生这个过程中没有起到主要的作用。使用6-BA后,胎生苗的产生时间推迟13.2 h。PR处理的胎生苗产生时间提前2.4 h。说明在胎生苗产生过程中,细胞分裂素起到了一个相对重要的作用。同时添加2,4-D和PR,胎生苗的产生时间推迟10.8 h;与单独施用2,4-D相比,对胎生苗产生时间的抑制增加了8 h。说明完整叶片中细胞分裂素和生长素存在着拮抗作用,在抑制细胞分裂素的运输后,生长素就完全的表现出对胎生苗产生的抑制作用。同时施用6-BA和TIBA,胎生苗的产生时间推迟14.4 h;与单独施用6-BA相比,对胎生苗产生时间的抑制增加了1.2 h。说明在完整叶片中,生长素对胎生苗产生影响不太,细胞分裂素决定着胎生苗产生的时间。同时施用2,4-D和6-BA,胎生苗的产生时间推迟了9.2 h,与单独施用6-BA和2,4-D比较,对胎生苗的抑制的作用介于单独施用细胞分裂素和生长素之间。这说明在完整叶片中细胞分裂素和生长素存在着拮抗作用,同时细胞分裂素在控制胎生苗产生的过程中起决定性作用。
PR处理的外植体,胎生苗产生所需的时间最短,在图2B中,在2~5 d这段时间内,用PR处理产生的胎生苗是最多的。TIBA处理外植体产生的胎生苗总数,在6~11 d这段时间内产生的胎生苗数最多。这说明了在完整叶片中,细胞分裂素和生长素对胎生苗的诱导产生都存在抑制作用,在施用对应的抑制剂PR、TIBA后,胎生苗的产生不再受到抑制,胎生苗的数量也得到了提高。使用6-BA处理和6-BA、TIBA处理的外植体胎生苗产生所需的时间相对较长,所以,所得到的胎生苗总数相对较少。
2.3 以切割叶片作为外植体在不同处理情况下诱导产生胎生苗的时间和数量差异
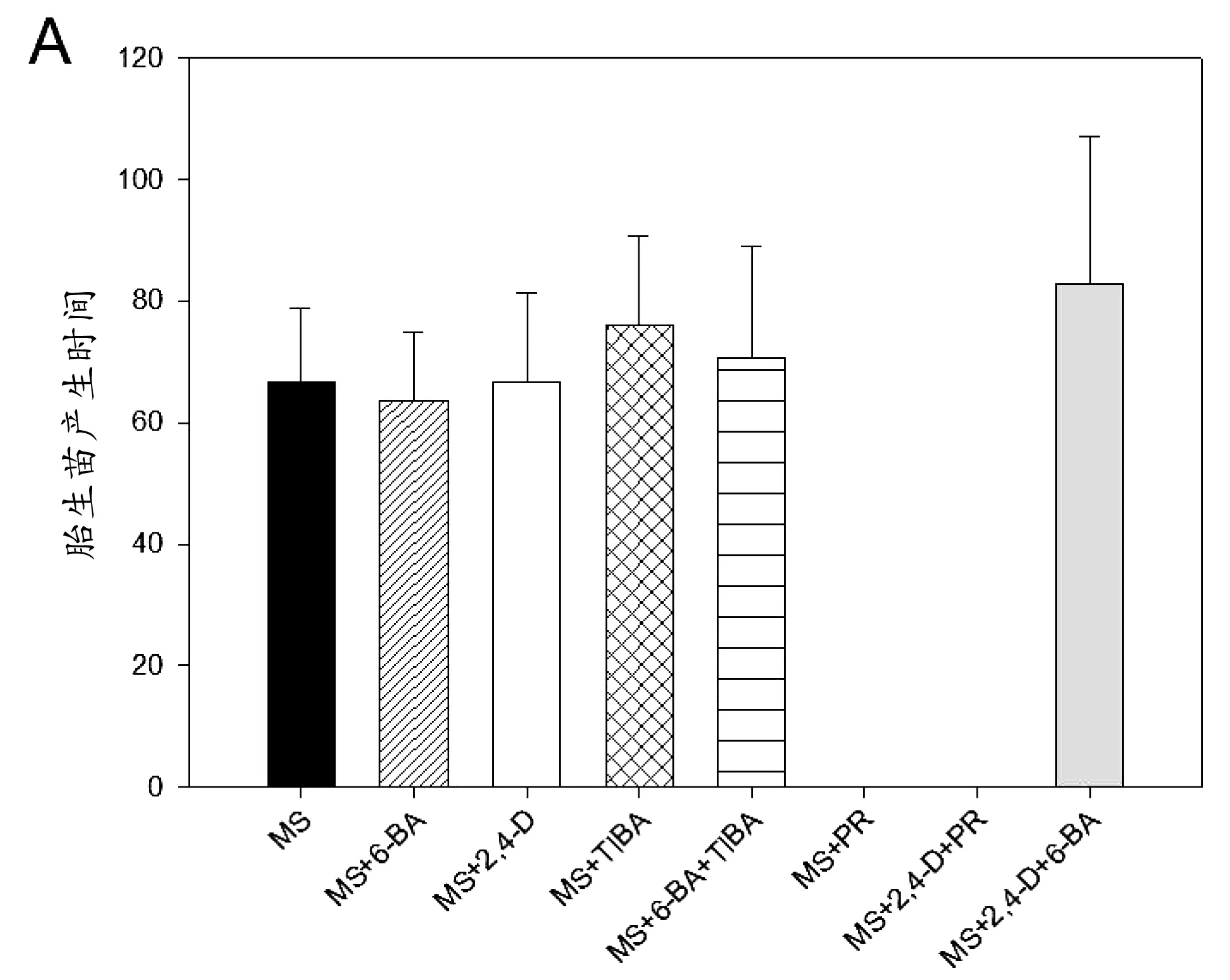
图3 以落地生根切割叶片作为外植体在不同处理情况下诱导产生胎生苗的时间和数量差异
与在以大叶落地生根完整叶片为外植体不同,以大叶落地生根切割叶片为外植体,施用6-BA使胎生苗的产生时间提前了3.1 h。说明在切割叶片中,细胞分裂素对胎生苗的产生起到了促进作用。而且用细胞分裂素抑制剂PR处理的切割叶片,胎生苗完全受到抑制。说明在切割叶片中,细胞分裂素对诱导胎生苗产生是至关重要的,并且有利于胎生苗的产生。 施用2,4-D进行处理,对胎生苗产生时间影响不大,但是施用生长素抑制剂TIBA的处理,胎生苗的产生时间推迟了9.3 h。这些结果说明在切割叶片中,生长素对胎生苗产生有一个正向的影响。同时用2,4-D和PR处理切割叶片,不能诱导胎生苗产生,说明2,4-D不能挽救无细胞分裂素的情况。同时用6-BA和TIBA处理,胎生苗的产生时间推迟4 h。与单独施用TIBA相比,6-BA促进了胎生苗的产生;与单独施用6-BA相比,TIBA的加入使得胎生苗的产生受到抑制,这说明在切割叶片诱导胎生苗产生的过程中,生长素起到了一个不可忽视的作用,抑制生长素的运输后,胎生苗的产生也受到抑制。
6-BA促进切割叶片中胎生苗的产生,因此6-BA处理过的切割叶片产生的胎生苗总数是最多的。2,4-D处理的切割叶片,诱导产生的胎生苗总数比对照多。细胞分裂素和生长素都诱导产生更多的胎生苗,说明在切割叶片中,这两种激素都起到了促进胎生苗产生的作用。与同时用6-BA和2,4-D处理的切割叶片相比,生长素抑制剂处理诱导产生的胎生苗总数相对较少,说明胎生苗的长期发育过程中,两种激素的共同作用有利于胎生苗的发育。
2.4 不同处理对胎生苗发育的影响
在0~5 d,凡是添加6-BA处理的胎生苗叶片直径大于其他处理的。其中,同时用2,4-D和PR处理后的胎生育,叶片直径是最小的。在6~11 d,相对于对照,单独用6-BA处理后的胎生苗叶片直径稍大于对照;同时用6-BA和TIBA处理的胎生苗叶片直径稍小于对照。同时添加2,4-D和PR处理的胎生苗叶片直径是最小的。在11~17 d,对照的胎生苗叶片直径是最大的。6-BA、6-BA+TIBA处理的胎生苗叶片直径大小相似,小于对照的胎生苗直径。2,4-D+PR处理的胎生苗叶片直径在整个发育阶段一直是最小的。而且,单独用PR处理的胎生苗叶片直径在整个发育阶段,也是一直落后于其他处理。

图4 胎生苗在不同处理下的叶片直径生长变化情况
3 讨论
在完整叶片中,2,4-D和6-BA都表现出抑制胎生苗产生的作用,施用生长素抑制剂TIBA能略微缓解这种抑制作用,而施用细胞分裂素PR抑制剂能完全阻碍这种抑制,并能促进胎生苗的产生。而在切割叶片中,6-BA表型出促进胎生苗产生的作用,而2,4-D作用不明显。相应地,施用生长素抑制剂TIBA,抑制了胎生苗产生。特别是施用细胞分裂素抑制剂PR完全地阻止胎生苗的产生。不同外植体诱导产生的胎生苗总数也与胎生苗产生时间呈现类似的趋势。在不同的外植体中,相同浓度的两种激素表现出的作用完全不同,甚至相反。某些研究认为,由于培养基中的蔗糖与细胞分裂素存在拮抗作用(Kulka 2005;Hartig and Beck 2010;Rolland,Baena-Gonzalez,and Sheen 2006;Moore et al,2003),所以高浓度的蔗糖(3%)会阻碍细胞分裂素的作用,造成实验结果不同,但低浓度的蔗糖(0.3%)则不会对细胞分裂素产生拮抗作用。本实验中,所有实验都是在含低浓度蔗糖(0.3%)的培养基上进行的,而且培养的条件完全一致,因此,我们推测相同浓度的激素表现出不同作用的原因可能是由于切割叶片破坏了叶片中存储的植物激素的平衡和运输,也有可能是破坏了叶片中信号传导途径。
本实验中,细胞分裂素和细胞分裂素抑制剂对完整叶片和切割叶片两种外植体类型都相应地产生明显的抑制或者促进作用,特别是在切割叶片中,施用细胞分裂素抑制剂PR完全地抑制了胎生苗的产生。其他一些学者认为细胞分裂素对胎生苗的作用可以分为两条途径,一条途径,细胞分裂素直接抑制胎生苗的产生,使胎生苗原基保持休眠状态。另一条途径,细胞分裂素间接的刺激胎生苗原基的发育,这种情况在叶片出现伤口或者细胞分裂素运输受阻的情况下很明显(Kulka 2005)。这个模型类似于Ferreira和Kieber(Ferreira et al. 2007)提出的钟形模型,根中的细胞分裂素在高浓度时抑制根的生长,在低浓度时促进根的生长。这两个模型都能解释细胞分裂素在完整叶片中抑制胎生苗产生,而在切割叶片中促进胎生苗的产生,而我们认为在胎生苗的产生过程中,细胞分类素直接作用胎生苗原基,处于调控途径中的下游,而生长素则处于上游。在这个过程中,推测生长素和细胞分裂素存在拮抗作用,若其中一种激素过量会使另一种激素含量降低(Moubayidin,Di,and Sabatini 2009;Fukaki and Tasaka 2009;Shimizu-Sato,Tanaka,and Mori 2009;Hwang,Sheen,and Müller 2012;Staffan et al. ;Zhi et al. 2013;SU et al.,2011),所以生长素和生长素抑制剂处理对完整叶片中胎生苗的产生时间和数量影响不大。
在胎生苗发育过程中,首先起作用的激素也是细胞分裂素,6-BA处理的胎生苗叶片直径明显大于其他处理。而细胞分裂素抑制剂处理的胎生苗,叶片直径一直都是所有处理中较小的。接着,激素的作用逐渐减小,对照中的胎生苗直径相对是最大的。所以,在胎生苗的发育过程中激素的作用不是一成不变的,6-BA在胎生苗发育前期刺激其生长,但是在后期作用逐渐减小。虽然,细胞分裂素在胎生苗的产生和发育过程中起到了主要作用,但是仍然不能忽略生长素的作用。在胎生苗总数中,2,4-D处理的总数在统计后期都不同于前期,说明在胎生苗发育的后期生长素的作用逐渐重要。
作者贡献
刘华是本研究的实验设计者和实验研究的执行人,完成数据分析,论文初稿的写作,参与实验设计和试验结果分析;钟天秀是项目的构思者及负责人,指导实验设计、数据分析、论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由国家自然基金(31802116)和贵州省科技计划项目(LH[2017]7027)共同资助。参与本研究的全体人员向资助方表示衷心感谢。
