东海中南部秋季游泳动物多样性及群落结构
2021-11-19张琥顺宋普庆刘世刚王雪华郑进顺林龙山
张琥顺,宋普庆,李 渊,刘世刚,王雪华,郑进顺,林龙山*
(1.上海海洋大学海洋科学学院,上海 201306;2.自然资源部第三海洋研究所,福建 厦门 361005;3.福建省宁德市霞浦县海洋与渔业局,福建 宁德 355100)
东海为太平洋西部边缘海,较浅的水深使得本海区光合作用充分,又因受到黑潮分支台湾暖流、东海沿岸流等水系的影响,饵料生物丰富,是多种渔业生物繁殖、索饵和越冬的良好场所,也是我国渔业资源生产力最高的区域之一[1]。近年来,随着捕捞强度的增加以及海洋污染的加剧,以大黄鱼(Larimichthyscrocea)、小黄鱼(Larimichthyspolyactis)等为代表的东海生物资源退化严重,即便在国家实施严格的休渔、禁渔、护渔措施后,一些传统的经济种类仍未见明显的资源恢复迹象[2]。在这一背景下,我国学者对东海渔业生物的多样性和群落结构开展了广泛研究,但其研究区域多集中于长江口[3]、东海中北部[4-5]及黄、东海交界海域[6-7],少有涉及到东海中南部海域。
东海中南部海域形成了鱼山渔场、温台渔场、闽东渔场等数个优良渔场,是我国重要的渔业生产基地。朱文斌等(2014)分析了东海南部头足类生物多样性和群落结构特征与海域主要环境因子的关系[8],但未涉及其他渔业生物;李圣法等(2005)根据1997—2000年的底拖网数据分析了东海中南部鱼类的群聚特征[9],但研究时间已相对较为久远。在自然环境变化以及人类活动的综合影响下,东海中南部游泳动物的群落结构可能已经发生了较大的变动,但近十年内对此海域游泳动物生物多样性和群落结构等方面的研究却鲜有报道,因此,对该海域游泳动物的相关研究是十分必要的。
本研究根据2018年11月东海中南部渔业资源调查资料,分析了该海域游泳动物的种类组成、优势种、种类多样性与群落结构等,并对该海域群落结构与环境因子的关系进行了初步探讨,以期为东海中南部渔业资源的可持续利用及科学管理提供依据。
1 材料与方法
1.1 样品采集与处理
2018年11月对东海中南部海域进行了游泳动物调查,设计7个断面共布设了49个站位(图1)。底拖网网具长127 m,网口宽27 m,网囊网目15 mm。调查时每个站点拖网时长为1 h,拖速约3 kn。渔业资源调查与采样方法按照《海洋调查规范》[10]进行,记录每个站位取样比例,将样品冷冻保存带回,在实验室根据文献资料[11-13]进行分类鉴定以及生物学数据的测量。
图1 东海中南部海域游泳动物调查站位
同时,在每个站位使用CTD同步采集该站位的水深、温度和盐度数据。
1.2 相对重要性指数
东海中南部游泳动物优势度通过相对重要性指数(Index of Relative Importance,IRI)进行计算[14],其公式为:
IRI=(w+n)×F×104
(1)
式(1)中:w为某一种类重量占总重量的比例(%),n为某一种类尾数占总尾数的比例(%),F为某一种类出现的站数占总站数的比例(%)。优势种的划分参考林楠等(2009)采用的标准,将IRI≥500的种类定为优势种[5]。
1.3 数量生物量比较曲线
数量生物量比较曲线(Abundance Biomass Comparison Curve,简称ABC曲线)通过将数量优势度曲线和生物量优势度曲线放在同一坐标系中比较,以两条曲线的分布特征来分析群落的受干扰情况[15]:群落结构稳定、未受到干扰时,生物量优势度曲线分布于数量优势度曲线之上;当两条曲线相交时表明群落受到中等程度干扰;当数量优势度曲线在生物量优势度曲线之上表明群落受到严重干扰。通过W统计值来表示ABC曲线中数量和生物量的相对关系[16]:
(2)
式(2)中:Bi和Ai分别为ABC曲线中种类序号对应的生物量和数量占比(%),s为总种类数。当生物量优势度曲线在数量优势度曲线之上时,W为正值,反之则为负值。
1.4 游泳动物群落多样性分析
运用下列公式[17]对东海中南部游泳动物进行群落多样性分析:
d=(S-1)/log2N
(3)
H′=-∑Pilog2Pi
(4)
J=H′/Hmax
(5)
式(3)至(5)中:d为丰度,H′为种类多样性指数,J为均匀度,S为站位出现种类数,N为总个体数,Pi为第i种渔获物的重量占总的渔获物重量的比例。
1.5 群落结构分析
选取生物量比例超过0.1%且出现频率大于5.0%的游泳动物种类作为分析对象,对其生物量进行4次方根转换,以削弱极端值的影响[9,18]。计算各站位间Bray-Curtis相似性系数(dij)矩阵[19]:
(6)
式(6)中:Yij和Yim分别是第i种类游泳动物在j站位和m站位生物量转化后的数值。采用未加权的组平均法对游泳动物群落进行聚类分析,采用非度量多维度标度(Non-metric Multidimensional Scaling, NMDS)对游泳动物群落进行排序,这两种方法相互补充,可以验证分析结果的准确性[9,20]。以拟合优度(Stress)来衡量NMDS的分析结果[21]:Stress<0.20时认为NMDS的二维点图具有一定的解释意义,Stress<0.10时可以认为是一个好的排序,Stress<0.05时认为此排序具有很好地代表性。
不同游泳动物组群的显著性检验通过单因子相似性分析进行。以相似性百分比分析的计算结果划分造成各群组内群落结构相似的典型种以及造成各群组内群落结构差异的分歧种,并分析它们对组内相似性及组间差异性的贡献率[22]。通过生物-环境变量相关性分析(Biota-environment Matching, BIOENV)找出与群落结构相关性最密切的环境因子组合[23],模型中使用的环境因子为水深(D)、表层水温(ST)、底层水温(BT),表层盐度(SS),底层盐度(BS)。
2 结果与讨论
2.1 种类组成及优势种
2018年11月东海中南部底拖网调查共捕获游泳动物种类181种,隶属于25目82科142属(表1),其中:鱼类25目65科111属133种,甲壳类2目9科21属34种,头足类3目8科10属14种,分别占种类总数的73.48%、18.78%、7.74%。鱼类中,鲈形目的种类数最多,有26科51属63种,占鱼类种类总数的47.37%;其次为鳗鲡目7科10属11种,占比为8.27%。头足类中,枪形目的种类数最多,有4科5属6种,占头足类种类总数的42.86%;甲壳类中十足目的种类数最多,有8科20属31种,占甲壳类种类总数的91.18%。

表1 东海中南部游泳动物种类分类阶元组成
从游泳动物种类数分布来看,处于调查海域北部的几个站位出现种类数较多,达50种以上,A4站位游泳动物种类数最多,有76种;浙江、福建沿海海域与台湾海峡北部海域次之,种类数在30~50种之间;位于27°00′N以南,远离大陆的几个站位游泳动物种类数较少,其中E5站位最少,仅有5种渔获物;其余站位渔获物种类数在10~30种之间(图2)。
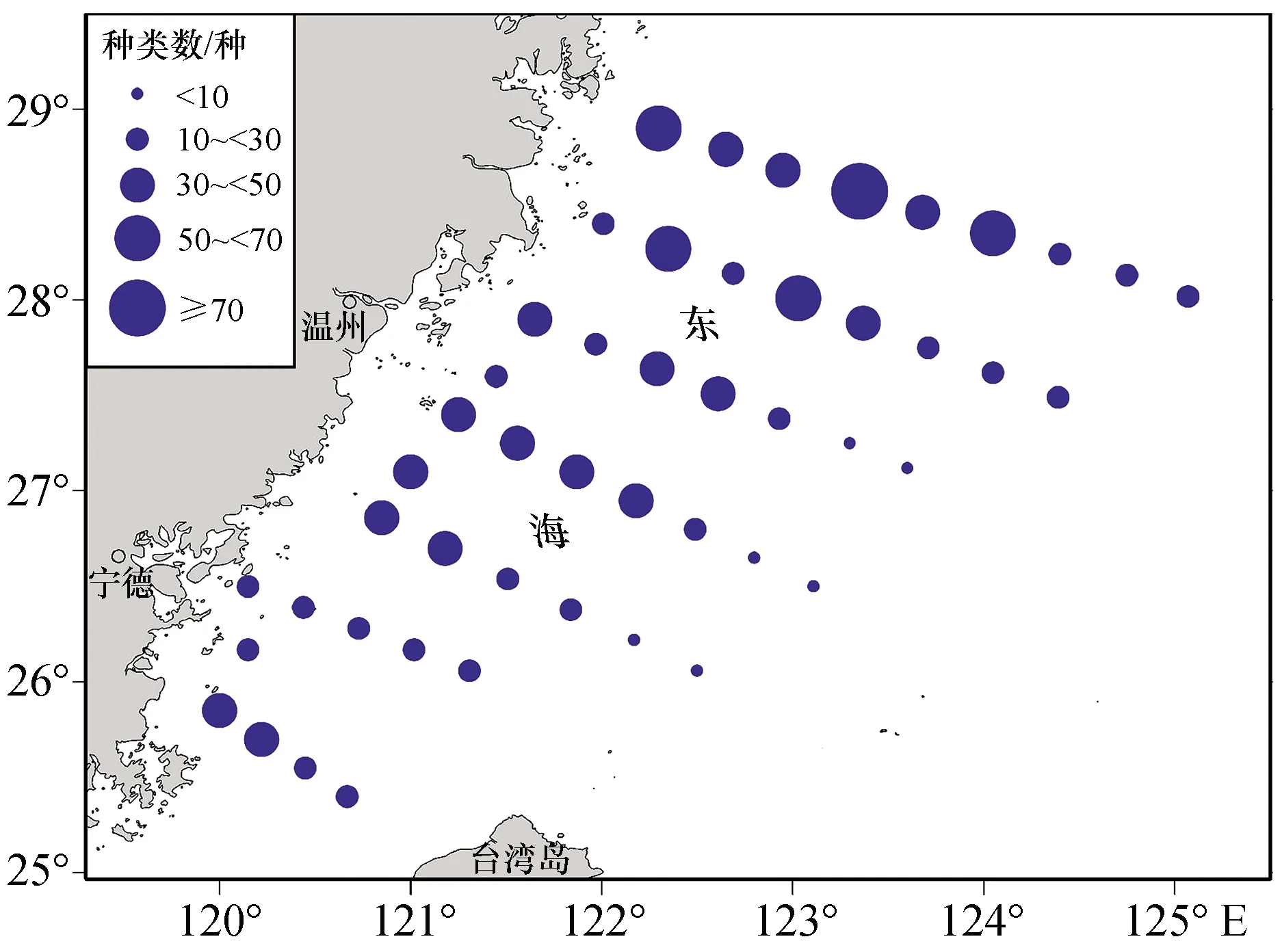
图2 渔获物种类数分布
对本研究中的游泳动物进行相对重要性指数分析,其中IRI值大于10的物种如表2所示。本研究优势种有5种,依次为七星底灯鱼(Benthosemapterotum)、带鱼(Trichiuruslepturus)、麦氏犀鳕(Bregmacerosmcclellandi)、龙头鱼(Harpadonnehereus)和日本发光鲷(Acropomajaponicum)。5个优势种的主要分布范围分别为:七星底灯鱼主要分布在90 m等深线以内的海域,在27°00′—29°00′N,123°00′E以西的近海区域生物量分布最高;带鱼在整个采样海区都有分布,无明显的生物量低值区;麦氏犀鳕的生物量在26°00′—28°00′N,123°00′E以西的近海海域分布较高;龙头鱼在25°00′—28°00′N,122°50′E以西的近海区域生物量较高;日本发光鲷的生物量主要分布在28°00′N以南,123°00′E以西,无明显的高值区。
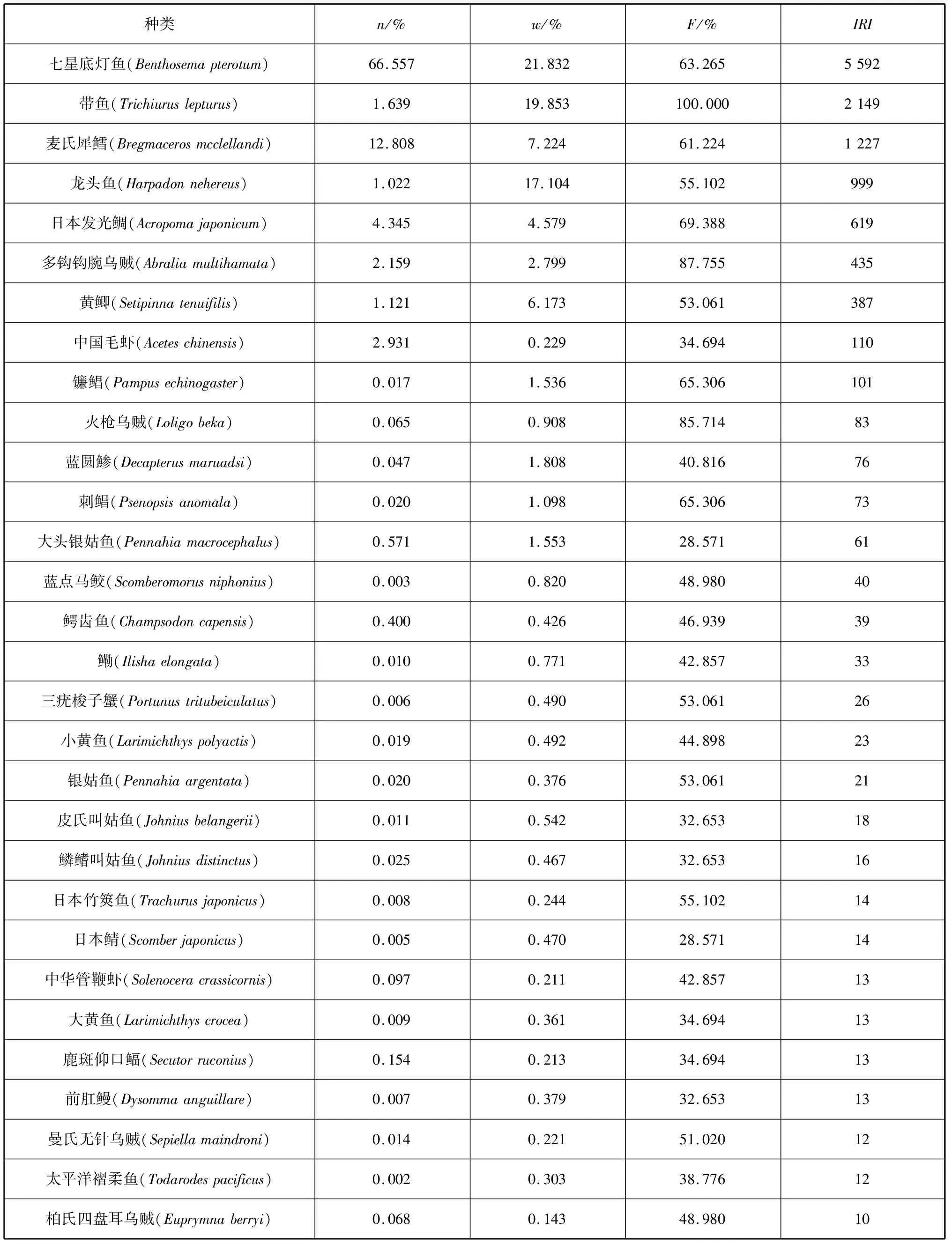
表2 东海中南部主要游泳动物相对重要性指数
优势种在生态系统中占据重要地位,对维持生态系统的稳定及渔业的可持续发展具有重要意义[24]。从表3可以看出,与历史资料相比,本研究优势种类存在一定异同:①2001—2018年之间的3次调查中,带鱼始终作为一种优势经济种出现;②前两次调查中作为优势经济种的小黄鱼和银鲳(Pampusargenteus)在此次调查中出现数量较少,生物量较低,被次要经济种类龙头鱼取代;③与以往相比,本研究优势种中经济价值较低、年龄结构简单的饵料鱼类(如七星底灯鱼等)比例较高,优势种存在小型化、低值化的特征。

表3 不同年份东海游泳动物优势种比较
带鱼是东海主要经济鱼类之一,也是本研究中唯一在全部站位都有出现的种类,且渔获率较高。宓崇道(1997)认为东海数种资源量较高的底层鱼类先后衰退,为带鱼的生长和繁殖提供了必需的饵料及空间条件,因而导致了带鱼产量的增加[25]。张寒野等(2005)分析了东海带鱼与7种小型饵料鱼类的空间异质性及空间关系,认为日本发光鲷等3种鱼类是带鱼秋季偏好摄食的鱼类,同时发现在带鱼的胃含物中,七星底灯鱼数量在冬季时明显多于其他季节[26]。由于捕捞强度的增加,带鱼和东海区其他经济鱼类一样在过去的十几年中出现了小型化和低龄化的趋势,但其资源量却没有明显的下降,在几次调查中均作为优势种出现[4,6,27]。结合以上研究结果,造成本研究带鱼资源量现状的原因可能为以下几个方面:一是作为凶猛的广食性鱼类,带鱼在秋季摄食强度达到最大[28],而调查区域日本发光鲷、七星底灯鱼等带鱼偏好的饵料鱼类的高资源量,保证了其在秋冬季节的摄食需求;二是同海区其他高级捕食者资源的衰退[29-30],导致生态竞争减少,或许也是使带鱼资源量常年居高的原因之一;三是常年的捕捞压力迫使带鱼群体对此做出适应性改变,如性成熟体长变小、初次性成熟年龄提早[25],逐渐由K-选择性向r-选择性转变。
2.2 ABC曲线特征
秋季东海中南部游泳动物数量优势度曲线始终位于生物量优势度曲线之上(图3),ABC曲线计算的W统计值为负数(-0.044),表明此时群落处于严重被干扰状态。数量优势度曲线中占比前三位分别为七星底灯鱼(66.6%)、麦氏犀鳕(12.8%)和日本发光鲷(4.3%);生物量优势度曲线占比前三位分别为七星底灯鱼(21.8%)、带鱼(19.8%)和龙头鱼(17.1%)。
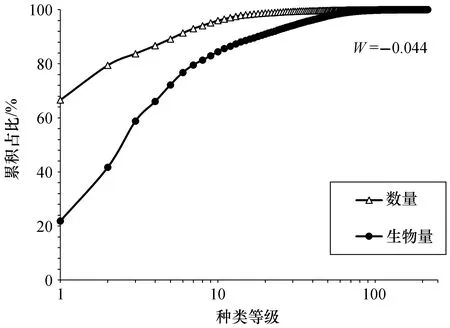
图3 东海中南部游泳动物ABC曲线
ABC曲线最早由Warwick(1986)用于评估海洋底栖无脊椎动物群落的受污染情况[15],近年来逐渐应用到海洋中其他生物群落的受干扰程度研究中。Blanchard等(2004)认为鱼类群落ABC曲线的特征是群落中大个体种类和小个体种类相对数量变化的反映,即群落中优势种的个体大小决定了ABC曲线中数量优势度曲线和生物量优势度曲线的位置[31]。李圣法(2008)在通过ABC曲线分析东海鱼类群落时,也认为群落种类组成中中小型个体的比例较高是造成ABC曲线特征的主要原因[32]。本研究渔获物组成多以小型游泳动物为主,优势种中七星底灯鱼和日本发光鲷等都属于小型饵料种类,其数量和生物量的占比差异是造成ABC曲线特征的主要原因,这也验证了Blanchard等[31]的结论。造成不同海域W统计值为负的原因是不尽相同的,这可能与群落中各种类的补充和生长、繁殖群体的迁移等内在因素以及过度捕捞、水域污染等人为因素有关[32-34]。对比同季节南海中部渔业生物ABC曲线W统计值[34],东海中南部游泳动物群落受到的干扰程度更高,作为我国渔业生产力最高的海域之一,东海具有悠久的捕捞历史,其W统计值为负可能受人为因素影响更大,过度捕捞使得大型个体所占比例减少,一方面幼体比例增多,另一方面对饵料生物的摄食强度下降;从内在因素来看,东海海域大多数鱼类在春末夏初产卵而在秋季初长成[35],这可能也是导致本研究小型鱼类居多的原因之一。
2.3 群落多样性指数
对各调查站位群落多样性指数分析显示,秋季东海中南部游泳动物d值范围为0.37~5.94,平均值为2.52,基本呈现北面高、南面低,近海高、外海低的分布趋势;H′值范围为0.38~2.64,平均值为1.51;J值范围为0.14~0.71,平均值为0.46。如图4所示,H′与J的高值分布具有明显的相关性:在25°00′—28°00′N之间是H′和J的高值区。
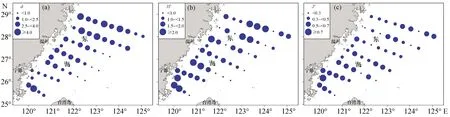
图4 东海中南部群落多样性指数的分布
对比同季节长江口、南海及台湾海峡游泳动物群落多样性指数可以发现,本研究各多样性指数值范围跨度较大,d及H′的平均值低于台湾海峡及南海海域而高于长江口海域,J值则低于同季节其他3个海域(表4)。

表4 不同海域游泳动物群落多样性指数比较
已有研究结果表明,在大陆架海域,生物群落的多样性随着水深的增加而增大,在大陆架边缘海域达到最高值,然后沿斜坡逐渐下降[37-39]。不同于上述结论,本研究各多样性指数范围跨度大,近海与外海差异明显:近海海域群落多样性水平相对较高,随着深度的增加多样性水平逐渐降低。也有研究发现鱼类群落多样性的区域变化除了与水深有关外,还与海域温度等环境因素有关[40]。本研究海域分布着两种不同的水系,海水温度、盐度等环境因素存在较大差异,这可能是造成本研究游泳动物群落多样性区域分布特点的原因之一。同时还有观点认为渔业调查采样设计会对群落结构产生影响:赵静等(2014)研究了不同采样设计对鱼类群落评估效果的影响,认为定点采样的效果要弱于简单随机采样和分层采样[41]。本研究采用了定点采样的方法,作业方式为底层拖网,而随着水深的增加,可能对中上层鱼类的捕获相对较低,同时,很多鱼类都有着昼夜垂直移动的习性[42-43],作业方式、作业时间的限制都有可能影响采样调查时渔获物的种类组成,进一步影响游泳动物的真实物种多样性水平。
2.4 群落结构特征
聚类分析结果将东海中南部秋季游泳动物生物群落分为两个群组(图5):近海群组和外海群组。NMDS排序分析的Stress值为0.08,表明NMDS的结果是一个好的排序(图6)。近海群组主要位于浙江南部以及福建北部沿海;外海群组主要位于26°00′—28°50′N,121°50′—125°00′E之间的海域(图7)。单因子相似性分析显示两个群组间游泳动物群落组成差异极显著(R=0.794,p<0.01)。
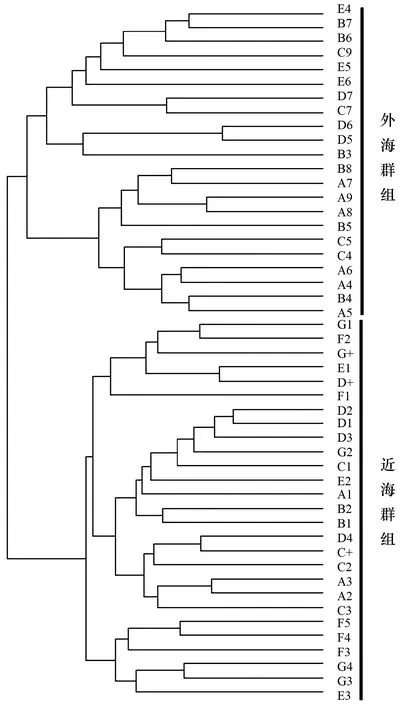
图5 东海中南部游泳动物群落聚类分析
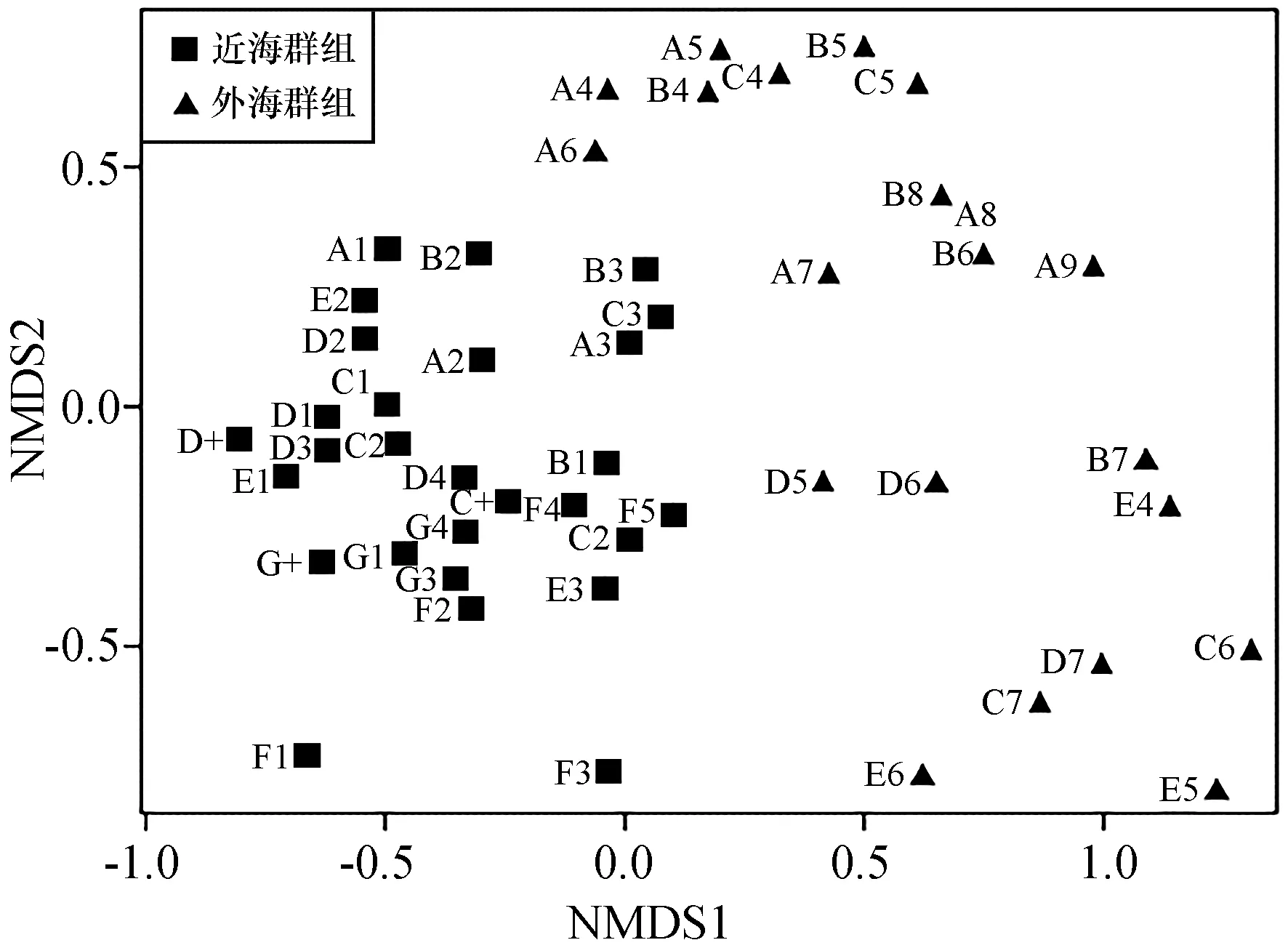
图6 东海中南部各站位NMDS二维点图
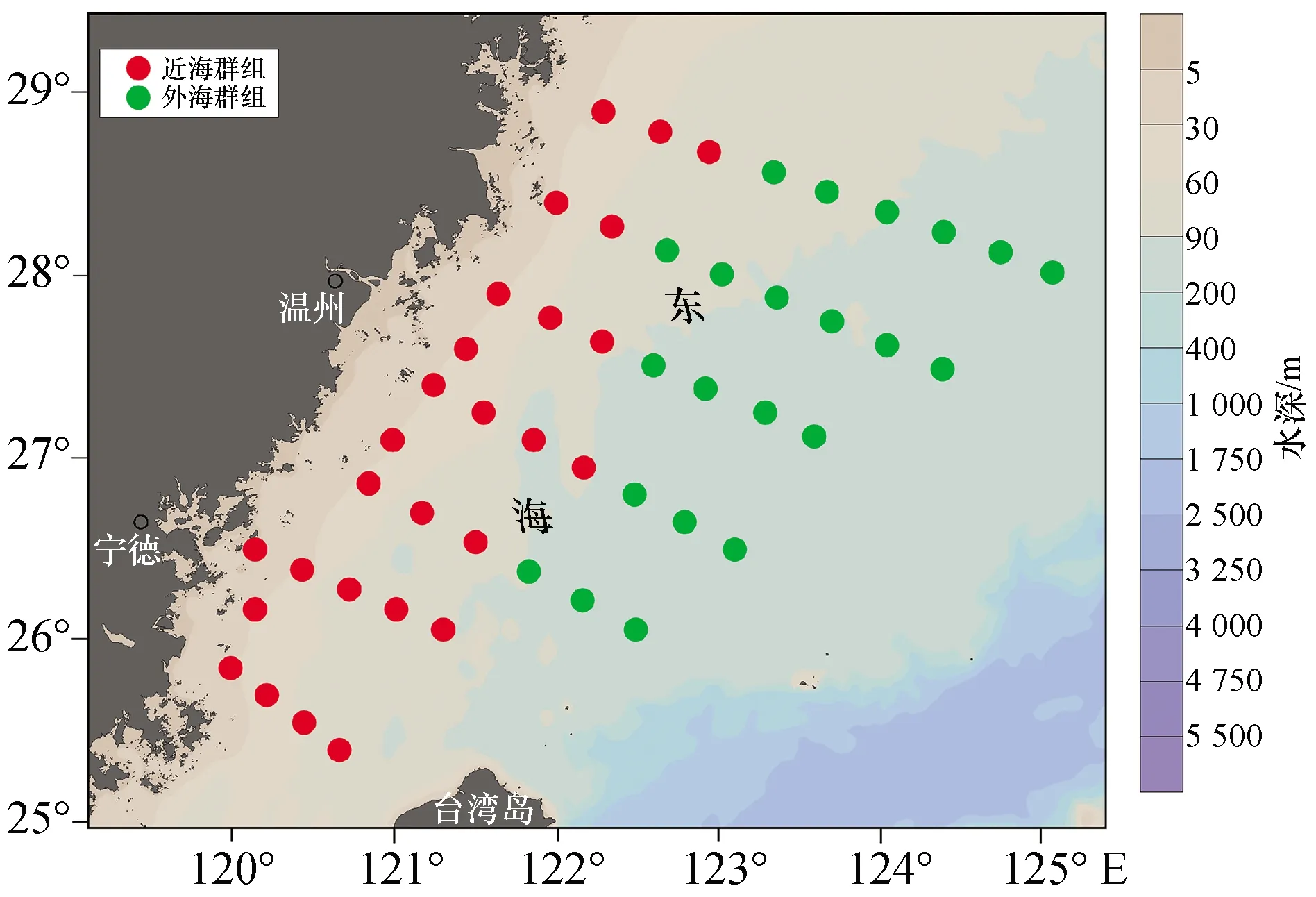
图7 东海中南部游泳动物群组的空间分布
BIOENV结果(表5)显示,与游泳动物群落结构相关性最密切的单个环境因子为水深(rw=0.55);与组合的环境因素的关系中,水深与表层水温两种环境因子的组合与群落结构相关性最好(rw=0.46)。综合比较,水深与游泳动物群落结构的关系最为密切。
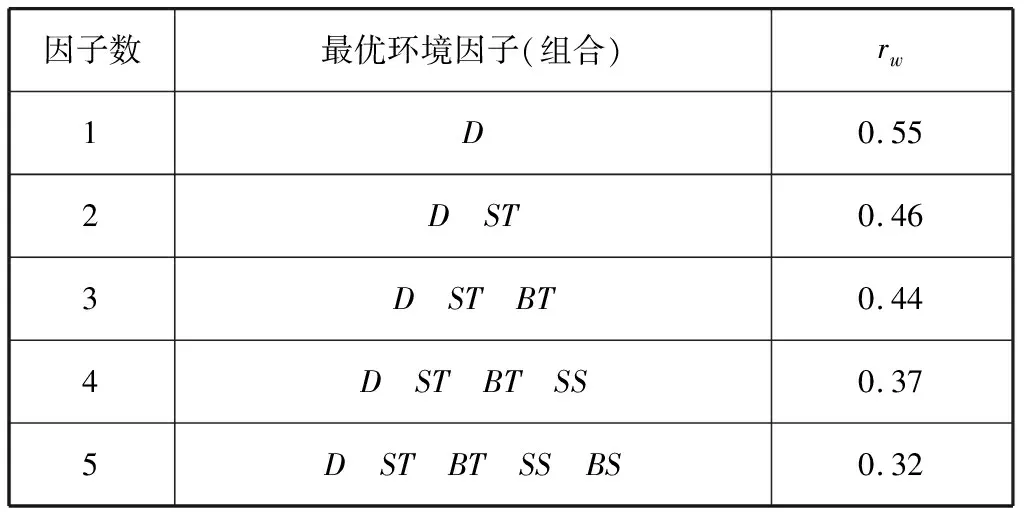
表5 BIOENV计算结果
本研究聚类结果与李圣法等对1997—2000年东海中南部鱼类群组的分析结果相一致[9],此结果验证了在大陆架及斜坡海域,水深是影响群组种类组成主要非生物环境因子的结论[44-46]。BIOENV结果显示水深是与游泳动物群落结构关系最密切的环境因子,这也验证了本次聚类分析结果的准确性。李圣法等将100~110 m水深作为近海群组与外海群组的过渡带[9],本次研究结果中两个群组之间大致以90 m等深线为界,两次调查中近海、外海两个群组的分界线变化不明显。除单一因素水深外,BIOENV显示水深与表层水温的环境因子组合也与群落结构表现出较强的相关性(rw=0.46)。刘尊雷等(2016)对台湾海峡海域鱼类群落结构与环境因子的关系进行了探究,发现底层盐度、底层温度、底层无机氮、表层无机氮、表底层温度差和水深对群落结构的影响较为显著[47];史赟荣等(2018)应用GAMs模型证实了长江口鱼类群落的物种时间分布格局主要受水温、径流及盐度等环境因子的控制[48]。由于本研究采集的环境因子有限,我们只选取了水深、表层水温、表层盐度、底层水温、底层盐度等5种环境因子来分析其与群落结构的关系,表层水温这一因素的背后可能隐藏了更多环境因子(如光照、温跃层、叶绿素a浓度等)的综合作用。
相似性百分比分析结果显示:①近海群组各站位的平均相似性为51.40%,对此贡献率较高的种类有带鱼、七星底灯鱼、镰鲳(Parnpusechinogaster)、龙头鱼、黄鲫(Setipinnatenuifilis)、多钩钩腕乌贼(Abraliamultihamata)、刺鲳(Psenopsisanomala)、火枪乌贼(Loligobeka)、鳓(Ilishaelongata)等;②外海群组各站位平均相似性为41.16%,带鱼、多钩钩腕乌贼、火枪乌贼、太平洋褶柔鱼(Todarodespacificus)、鳄齿鱼(Champsodoncapensis)、日本发光鲷、日本竹筴鱼(Trachurusjaponicus)等对此平均相似性贡献较高;③近海群组、外海群组的平均相异性为70.84%,对组间相异性贡献较大的种类主要有七星底灯鱼、龙头鱼、黄鲫、镰鲳、麦氏犀鳕等(表6)。
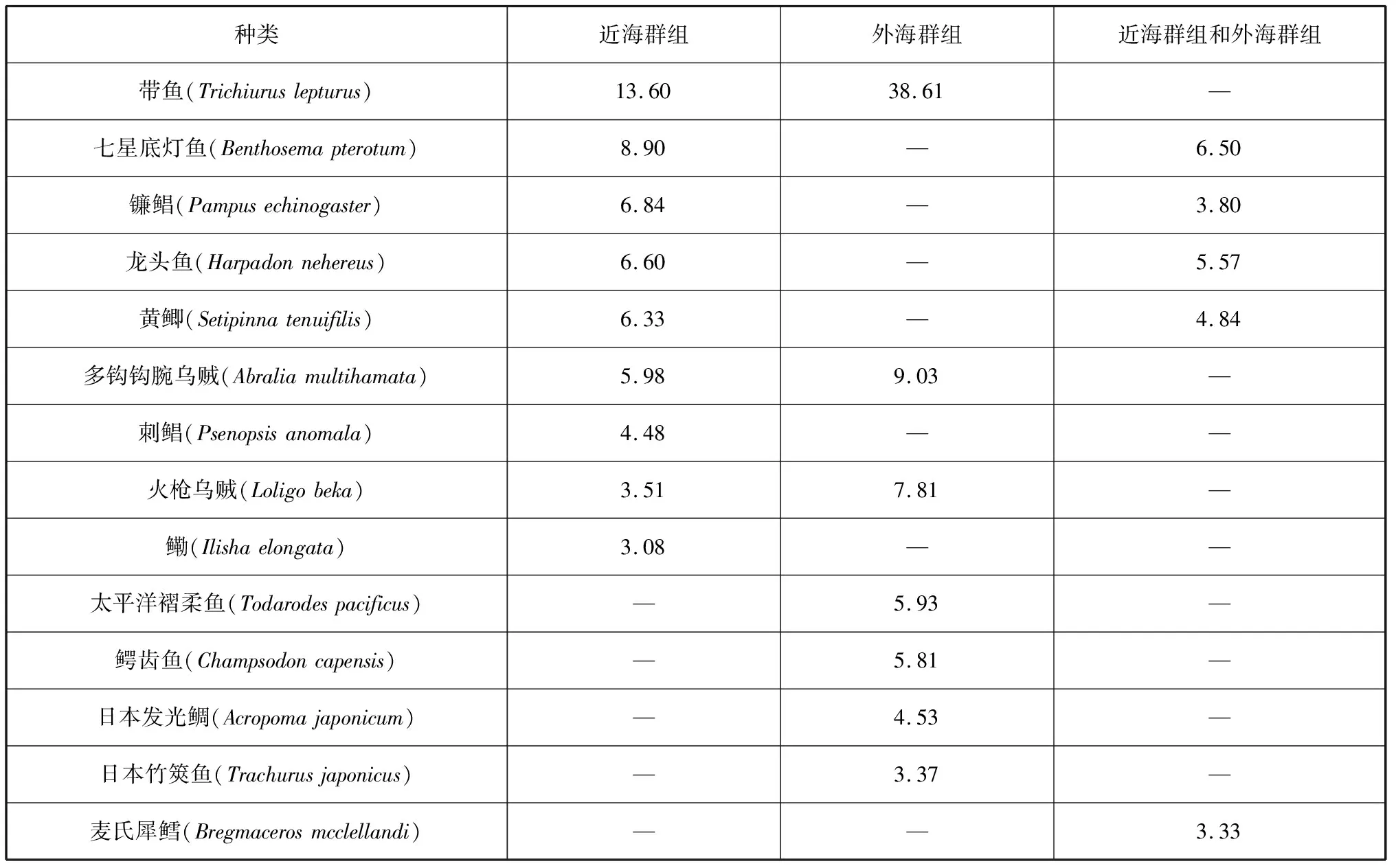
表6 典型种对组内平均相似性及差异种对组间平均相异性的贡献百分比
有研究显示渔业生物群落的分布规律与生态环境的变化密切相关,如离岸距离、河流入海等;不同群组的主要贡献种与物种的栖息环境、洄游习性等也有较强相关性[3, 49-51]。本研究海域主要分布着两种不同性质的水系:①由钱塘江、闽江等江河入海径流与周围海水混合形成的,具有低盐、高营养盐的特征东海沿岸流;②由黑潮分支台湾暖流组成的东海外海水,具有高温高盐的特征[1]。本研究中,近海群组受沿岸流影响较大,淡水、海水混合形成高营养盐水系为该近海海域提供了较高的初级生产力,保障了如七星底灯鱼,多钩钩腕乌贼、麦氏犀鳕等基础饵料生物种群的生长,确保了不同水层、不同摄食偏好的高等级游泳动物群体的食物来源充分,因此近海海域的游泳动物群落偏向于浮游动物食性(黄鲫、鳓、镰鲳等)以及游泳动物食性(龙头鱼、带鱼等)的种类在此聚集;而外海群组受黑潮分支台湾暖流影响更多,底层温度、盐度的季节变化不明显,饵料生物相对单一,造成了游泳动物种类数相对偏少,除了带鱼等本研究的广布性种类外,外海群组主要以近底层种类(日本发光鲷等)、大洋性洄游种类(太平洋褶柔鱼等)和偏好高盐暖温种类(日本竹筴鱼等)为主。
3 结论
本研究通过分析东海中南部海域游泳动物的生物多样性与群落结构特征,获得了如下结论:
(1)在海洋环境变化与人为捕捞压力等多重影响下,东海中南部游泳动物的群落结构已经发生了较大的变化,一方面表现为主要经济种类的更替:龙头鱼取代小黄鱼、银鲳等成为一种新兴的优势经济种类;另一方面表现为优势种构成中呈现出的小型化和低龄化特征。ABC曲线的特征也表明东海中南部的群落结构处于严重干扰状态,而群落中小个体种类的比例较高是造成ABC曲线特征的主要原因。面对长期的捕捞压力,带鱼群体做出了相应的适应性改变,同时海域内其他高级捕食者的资源衰退以及丰富的饵料生物资源为该种群的发展提供了必要的空间条件和饵料条件,这也是其在多次调查中均作为优势种出现的原因之一。
(2)研究还发现海洋环境对东海中南部物种的分布及群落结构的组成起着重要的作用,对调查区域影响最为显著的环境因子为水深,其次为表层水温。由于本研究采集数据有限,更多环境因子的综合作用未体现出。今后应当密切关注优势种、群落结构对该海域海洋环境变动的响应,以应对全球气候变化对我国沿海渔业生产的挑战。
