Cry1B抗独特型单链抗体的定点突变及生物活性分析
2021-11-19仲建锋李兴奎徐重新张霄刘贤金
仲建锋 李兴奎 徐重新 张霄 刘贤金
(1. 江苏省食品质量安全重点实验室—省部共建国家重点实验室培育基地 农业农村部农产品质量安全控制技术与标准重点实验室 江苏省农业科学院 农产品质量安全与营养研究所,南京 210014;2. 郑州职业技术学院,郑州 450121)
苏云金芽孢杆菌(Bacillus thuringiensis,Bt)是一种广泛存在于各类环境中的昆虫病原菌,对多种农业害虫具有杀虫活性[1]。Cry1B是Bt Cry毒素的一种,其对鳞翅目昆虫的靶标位点包括中肠刷状缘膜囊泡(brush border membrane vesicles,BBMV)上的氨肽酶N(aminopeptidase N,APN)、钙粘蛋白(cadherin)[2-3], 以 及 ABC 转 运 蛋 白[4]等。APN属于锌结合金属蛋白酶家族蛋白,它的碳端结合部位富含N-乙酰氨半乳糖胺(N-acetylgalactosamine,GalNAc),这些区域是Cry毒素的结合位点[5]。烟芽夜蛾(Heliothis virescens)的170 kD的APN可与Cry1Aa、Cry1Ab 和 Cry1Ac结合[6]。卷叶蛾(Epiphyas postvittana)BBMV上的120 kD的APN可与Cry1Ac和Cry1Ba结合[7]。我们前期研究发现Cry1B抗独特型(anti-idiotypic,anti-Id)单链抗体(single chain fragment variable,scFv)C7可以模拟Cry1B的结构和功能[8],因此推测C7与昆虫BBMV上的APN可能存在结合关系。但是C7的结合和生物活性跟原毒素Cry1B相比有差距,需要改进提升。
单链抗体由重链可变区(heavy chain variable,VH)和轻链可变区(light chain variable,VL)通过一条单一肽链(linker)组成,大小为完整抗体分子的1/6,制备流程较简单,易进行分子改造[9-10]。然而,基因工程抗体一般存在亲和力低等问题,以计算机模拟为基础的定点突变可以预测分子间结合区域并确定关键氨基酸残基[11],关键氨基酸的有利突变可以提高抗体的亲和力,在体外通过定点突变等基因工程手段可以构建限定性突变抗体库,进而筛选到高亲和力抗体[12]。Barderas等[13]在研究人类抗胃泌素TA4 scFv亲和力成熟的过程中,在限定性突变的基础上采取定点突变等方法,最终得到将TA4的亲和力提高了454倍的突变scFv。Zhang等[14]在构建了抗成纤维细胞活化蛋白scFv E3突变库的前提下,对CDR区进行了定点突变,其中重链33位氨基酸Asp变为Gly,107位Tyr变为His效果最好,此E3 Mut2亲和力提高了4倍。此方法目的明确,避免了随机突变编码可变区基因片段法随意性大的缺点。
分子模拟技术在抗体结构与功能关系分析、蛋白质分子设计与改造等过程中得到了广泛开发与应用。本研究通过计算机辅助设计指导抗体亲和力成熟,借助分子模拟技术获得对接复合物的关键结合区域并预测热点,利用热点构建饱和突变抗体库,筛选出亲和力和生物活性提高的突变体。以期进一步对基因工程抗体结合区域改造指明方向。
1 材料与方法
1.1 材料
抗Cry1B毒素Ab2β 型scFv-C7来源于本实验室,使用噬菌体展示技术从Tomlinson I+J库中筛选获得,具杀虫活性[10],还得到了与C7结构相似的scFv-D5,但无杀虫活性(图1)。CrylB购于上海佑隆生物科技有限公司;大肠杆菌(Escherichia coli)TG1购自英国剑桥MRC实验室;辅助噬菌体M13K07、Nco I酶、Not I酶、T4 DNA连接酶和Dpn I酶均购于NEB公司;使用的其余试剂均为分析纯。

图1 CrylB毒素的anti-Id scFv C7和D5的氨基酸序列分析Fig. 1 Amino acid sequence alignment of anti-Id scFv C7 and D5 against Cry1B toxin
稻 纵 卷 叶 螟(Cnaphalocrocis medinalis) 源自扬州绿源生物化工有限公司。试虫饲养温度为(27±1)℃,相对湿度80%±5%,光照时间14 h,以水稻(Oryza sativa)叶片续代饲养。
1.2 方法
1.2.1 ScFv-C7与稻纵卷叶螟APN的同源建模和分子对接 ScFv-C7的同源建模从Protein Data Bank(PDB)数据库分别搜索VH、VL的同源序列,根据一致性确定模板。在分别构建VH和VL的同源模型后,通过最小二乘法对蛋白质主链结构重叠以构建C7三维结构。稻纵卷叶螟APN(CmAPN,NCBI登录号:ADZ05466.1)的建模则从PDB数据库搜索一致性较高的模板即可。采用Ramachandran plot对C7和APN三维结构进行评估,评价其合理性。建模采用的Modeller程序由EasyModeller 4.0软件[15]完成,Ramachandran plot分析由PROCHECK模块评估(https://saves.mbi.ucla.edu)完成,蛋白三维结构输出使用 PyMOL 1.3 软件[16]。
使用ZDOCK在线程序(http://zdock.umassmed.edu/)对scFv-C7和CmAPN进行蛋白对接。采用KFC热点预测(http://kfc.mitchell-lab.org/)计算分析scFv-C7和CmAPN对接复合物,获得二者接触界面上互作的热点残基。
1.2.2 ScFv-C7定点突变抗体库的构建 采用大引物PCR法(megaprimer PCR)[17-18]进行饱和突变,分析scFv-C7位于对接界面上的热点残基,选择了scFv- C7的 H-CDR3区 A123、Y124和 L-CDR2区S204、S207作为突变位点。LMB3和pHENseq是用来扩增全长序列的引物;NNK_VH是引入H-CDR3区A123、Y124饱和突变的引物;NNK_VL是引入L-CDR2区S204、S207饱和突变的引物(表1)。所有引物均由上海生工生物合成。

表1 饱和突变抗体库构建使用的引物序列Table 1 Primer sequences for the construction of sitedirected mutagenesis library
普通 PCR 扩增条件:95℃ 5 min,94℃ 45 s,56℃ 45 s,72℃ 1 min,共30个循环,72oC 延伸10 min。“大引物”PCR 扩增条件:95℃ 5 min,94℃ 45 s,58℃ 45 s,72℃ 1 min,共35个循环,72℃ 延伸10 min。为去除PCR产物中未突变的的模板质粒,使用Dpn I酶消解处理。
分别将载体pIT2质粒与定点饱和突变产物使用Nco I、Not I双酶切处理,再用T4 DNA连接酶连接。使用BTX ECM 830电转仪(Harvard Apparatus公司)将连接产物转化至TG1感受态细胞中,电转产物快速加入1 mL 2×TY培养基,37℃,200 r/min复苏1 h;10倍梯度稀释后涂布于TYE-AG平板,置于37℃过夜培养,计算库容;然后将剩余复苏产物离心后用100 μL 2×TY重悬沉淀,并均匀涂布于TYE-AG平板,37℃过夜培养。次日,随机挑取10个单克隆,并用菌落PCR验证突变库的准确性,然后向TYEAG平板中加入1 mL 2×TY,用涂布棒刮起平板上的菌体细胞,并彻底混匀,所得即为定点饱和突变抗体库。
1.2.3 定点饱和突变抗体库的筛选
1.2.3.1 突变抗体库的扩增 取500 μL饱和突变库加入到200 mL 2×TY-AG培养基中,37℃、250 r/min培养至对数期(OD600=0.4),将2×1011PFU M13KO7辅助噬菌体加入到上述50 mL培养液中,37℃孵育30 min。剩余的150 mL培养液继续培养2 h,10 800× g离心10 min,去上清,并重悬至10 mL的2×TY(含15%甘油)中,最后分装保存至超低温冰箱。含辅助噬菌体的饱和突变库菌液孵育后,3 000 × g离心10 min,弃上清,再用100 mL 2×TY-AKG将菌体沉淀重悬,30℃过夜培养。次日,3 300 × g离心30 min,上清加入1/4体积的PEG/NaCl并充分混匀,冰浴1 h。3 300× g离心30 min,再用4 mL PBS重悬沉淀,最后11 600× g离心10 min,上清即为扩增好的饱和突变抗体库,可用于后续筛选。
1.2.3.2 稻纵卷叶螟BBMV对突变抗体库的富集和筛选 使用稻纵卷叶螟4龄末期幼虫的中肠提取BBMV,采用Mg-EGTA沉降法制备[19]。筛选策略参考课题组前期方法[20],略有改动。采用逐渐降低包被浓度的固相筛选方法对扩增好的饱和突变抗体库进行3轮筛选,稻纵卷叶螟BBMV(CmBBMV)的浓度分别是 100 μg/mL、50 μg/mL 和 25 μg/mL,以每孔4 mL包被6孔板。经过3轮“吸附-洗脱-扩增”的筛选,使特异性结合的噬菌体得到有效富集。
1.2.3.3 单克隆ELISA鉴定 参考剑桥MRC和课题组前期的方法[20],稍有改动,从第3轮筛选后的平板中随机挑取单菌落转接至96孔板中培养;次日,从中取2 mL菌液转移至新的96孔板中,培养2 h后,每孔加入25 mL滴度为109的辅助噬菌体M13KO7,培养、离心、去上清,每孔加入200 mL 2×TY-AKG重悬细胞,培养过夜;次日离心,取上清液备用。取30 mg/mL CmBBMV包被96孔板,每孔100 mL。次日封闭后加入每孔100 mL单克隆表达上清。其余均为ELISA标准操作步骤,使用酶标仪(Thermo)测定OD450值,阳性/阴性值大于2.1,即可判断为阳性克隆。
1.2.4 突变抗体基因的测序与分析 选取在上述单克隆噬菌体ELISA分析中OD450值高于野生型scFv-C7的突变单克隆,按标准流程培养后提取质粒、酶切、PCR鉴定和测序。序列比对分析使用ClustralW 2.0.10程序,图像输出使用Jalview 2.11软件。
1.2.5 突变单链抗体与CmBBMV的结合能力分析 表面等离子体子共振(surface plasmon resonance,SPR)技术是一种快速监测生物分子互作的方法。各样品菌液的制备方法同抗体库扩增方法一致,使用20 mL 2×TY-AG培养体系,次日PEG/NaCl充分混匀后冰浴、离心后弃上清,再用10 mL PBS重悬沉淀,最后11 600 × g离心10 min,上清即为待测样品,滴度为3.2×107CFU/mL。阳性对照Cry1B毒素浓度20 μg/mL。
使用BIAcore X100 实验平台(GE Healthcare),所用材料购自GE公司。CmBBMV先用PBS稀释至500 mg/L待用,进样反应再用NaAc稀释到50 mg/L。各待测样品均使用HBS-EP稀释到指定浓度。
CmBBMV作为受体蛋白,通过氨基偶联法固定到CM5传感芯片上。将CM5传感芯片FC1通道设为对照通道,FC2通道设为反应通道。活化FC2通道待基线平稳封闭,确定固定CmBBMV至芯片上的蛋白量。然后确定芯片再生反应所需再生缓冲液Glycine-HCl的最适pH值。所有待测样品匀速连续流过芯片表面,最后的传感图显示FC2-FC1的响应值(RU)。
1.2.6 突变单链抗体对稻纵卷叶螟的杀虫活性 采用浸叶法,生测样品的制备同SPR分析一致,最终得到PEG/NaCl沉淀的上清(滴度3.2×107CFU/mL),Cry1B浓度也是20 μg/mL。将新鲜水稻叶片分别放入各种生测材料中浸渍后取出晾干,用于实验。将处理后的水稻叶片分别放入培养皿,每皿接入20头3龄幼虫,24、48、72 h后分别记录试虫状态,每处理3次重复。试虫死亡率采用Abbott公式进行校正,并以平均数±标准误表示。各处理时间样品间采用单因素方差分析和Tukey显著性比较,使用SPSS 17.0软件进行数据处理。
2 结果
2.1 ScFv-C7与CmAPN三维结构的同源建模及合理性评估
CrylB毒素anti-Id scFv C7的VH选取人类生殖细胞抗体3-23/B3(PDB:3QOS)作为同源建模的模板[21],序列一致性为89%。C7的VL模板选取人类抗体Vκ域(PDB:2BX5)[22],序列一致性达到了98%。分别构建VH和VL的同源模型后,通过蛋白质主链结构重叠获得C7的结构模型(图2-A)。CmAPN的模板选择人类 APN Cd13(PDB:4FYQ)[23],序列一致性为31%,CmAPN的三维结构模型则如图2-B所示。
通过Ramachandran 图对C7和CmAPN结构模型主链构型的Φ-Ψ二面角的分布进行检查。结果C7的98.5% 残基落在最佳区域(图2-C);CmAPN中97.3%残基落在最佳区域(图2-D),说明模型结构合理可信。

图2 ScFv-C7和CmAPN三维结构的同源建模Fig. 2 Three-dimensional structure models of scFv-C7 and CmAPN
2.2 ScFv-C7与CmAPN的分子对接及热点预测
ScFv-C7和CmAPN对接后的空间结构如图3-A所示,使用KFC2热点预测系统分析得到scFv-C7上 的 热 点 有 :Y54、A123、Y124、Y186、Y203、S204、S207。一般来说,单链抗体氨基酸序列变化主要发生在VH和VL的CDR2、CDR3区,按照Kabat规则,上述预测的热点残基位于H-CDR3区(A123、Y124)和 L-CDR2 区(S204、S207)(图 3-B),可以选择这两个区域进行饱和突变。

图3 scFv-C7和CmAPN的分子对接及互作热点分析Fig. 3 Molecular docking and the interaction hot spot analysis between scFv-C7 and CmAPN
2.3 ScFv-C7饱和突变抗体库的构建
使用大引物PCR法对scFv-C7模板进行突变,以LMB3和NNK_VH为引物扩增引入H-CDR3区A123、Y124的饱和突变(图4-A),并当做下一轮PCR的上游引物(大引物1),加下游引物pHENseq扩增即为带有H-CDR3饱和突变位点的PCR产物(图4-B)。再以scFv-C7作为模板,NNK_VL和pHENseq为引物,引入VL区S204、S207的饱和突变(图4-C),作为下一轮的PCR下游引物(大引物2),与上游引物LMB3和模板(H-CDR3区突变位点的PCR产物)进行扩增,本次扩增的PCR产物即为引入H-CDR3和L-CDR2的饱和突变区域(图4-D)。
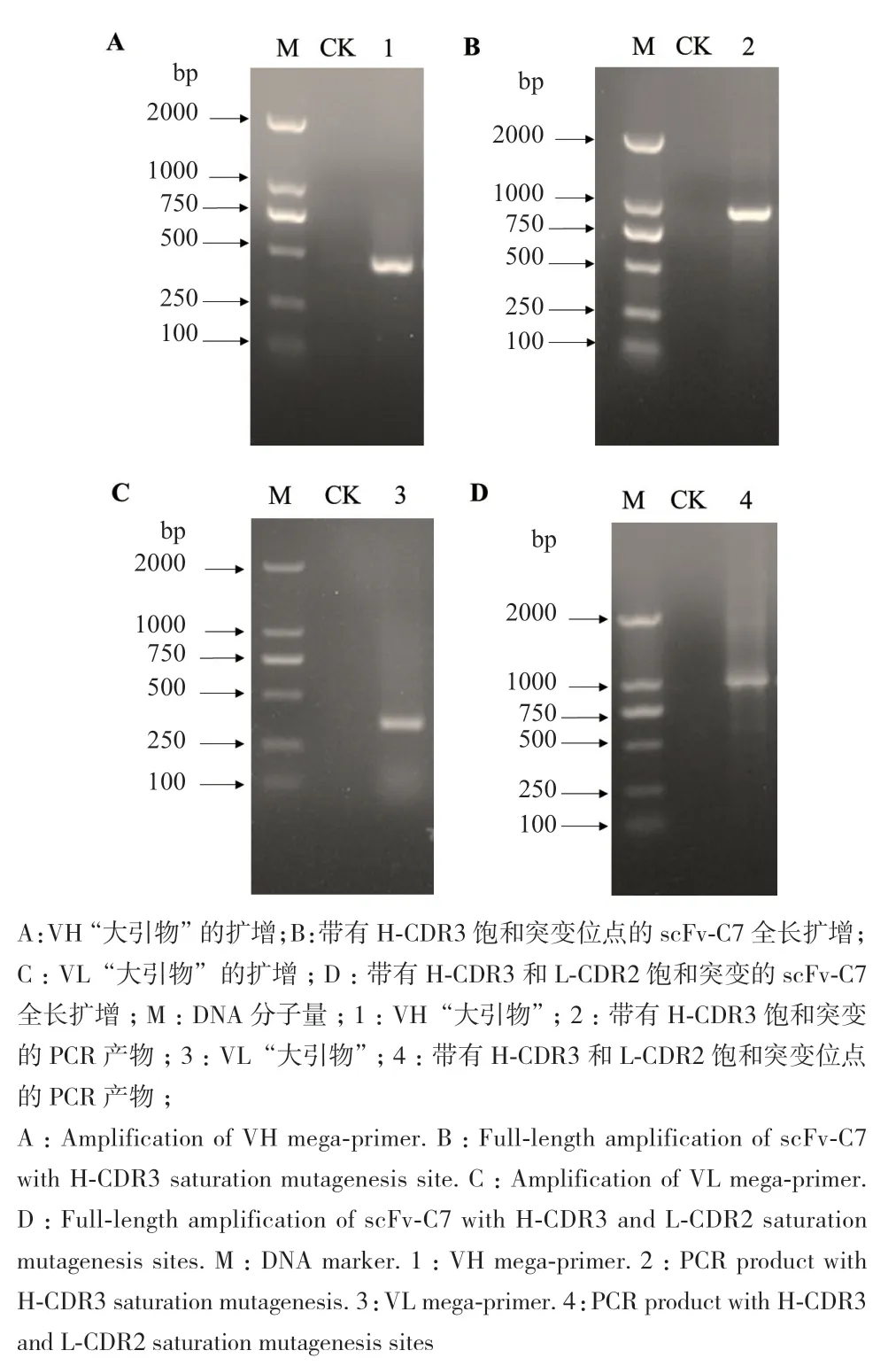
图4 基于scFv-C7对接界面热点氨基酸的饱和突变Fig. 4 Saturation mutagenesis of hot spot amino acids on docking interface based on scFv-C7
将scFv-C7饱和突变的PCR产物纯化后克隆至pIT2噬菌粒载体,随后电转化至大肠杆菌TG1感受态细胞,再将转化产物涂布于TYE-AG平板,构建定点饱和突变抗体库,经菌落计数法计算库容为2×103。
为了验证突变抗体库所含基因是否正确,随机挑取10个单克隆并利用噬菌体载体pIT2自带引物LMB3和pHENseq进行PCR验证,结果发现9个克隆在950 bp左右有目的条带(图5),符合噬菌粒基因大小,只有1个突变克隆PCR未成功。构建的饱和突变抗体库基因正确率90%,可见适合后续筛选工作。

图5 构建的饱和突变抗体库正确性验证Fig.5 Verification of correction from constructed saturation mutant antibody library
2.4 ScFv-C7定点突变抗体库的筛选
分析每轮筛选之后整体次级库的亲和力变化趋势,结果发现产出/投入随着筛选轮数而增高,第3轮比第1轮的产出/投入提高了12倍(表2),可见筛选3轮后特异性突变株得到了有效富集。

表2 scFv-C7突变抗体库的富集和筛选Table 2 Enrichment and screening of scFv-C7 mutant antibody library
2.5 单克隆ELISA筛选阳性重组噬菌体
随机挑取第3轮筛选的单菌落用于单克隆噬菌体ELISA鉴定,从OD450值大于C7的阳性克隆中选取了3个最高的单克隆:3A5、3G2和2F7(图6),用于进一步序列比对分析。
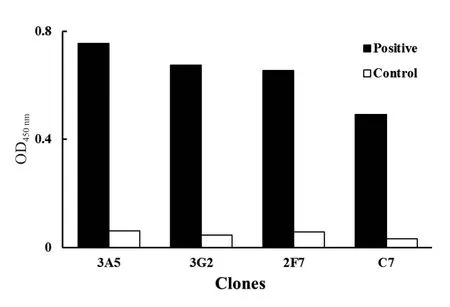
图6 突变抗体的单克隆噬菌体ELISA分析Fig. 6 ELISA analysis of monoclonal phage for mutant antibodies
2.6 筛选获得突变基因的序列分析
使用pIT2载体引物对3A5、3G2和2F7进行PCR检测,条带在950 bp左右(图7-A),匹配噬菌体完整外源基因大小,可用于进一步的测序分析。
多重序列比对显示3A5的H-CDR3区124位发生了突变,Tyr变成了Gly(Y124G);而3G2则无变化,跟C7序列完全一致,2F7序列的H-CDR2区76位出现了终止子(图7-B)。最终选择3A5即Y124G作进一步分析。
图7 突变单链抗体的基因克隆(A)与氨基酸序列比对(B)Fig.7 Gene cloning(A)and amino acid sequence alignment(B)of mutant scFv antibodies
2.7 突变单链抗体与CmBBMV的结合能力
SPR检测单链抗体与CmBBMV的结合过程,上 样 缓 冲 液 选 择 pH 4.0的 NaAc(10 mmol/L),CmBBMV偶联到芯片上的量为1 267 RU,再生反应选择pH 2.5的Glycine-HCl(10 mmol/L)和5 mmol/L NaOH。
各待测样品分别通过CM5芯片,它们同CmBBMV的结合是动态过程,先结合再解离。ScFv-D5同 CmBBMV基 本 没 有 结 合(3.57 RU),Cry1B的结合能力最强(197.39 RU),突变型Y124G的结合能力(155.49 RU)将野生型scFv-C7的结合能力(72.77 RU)提高了2.1倍(图8)。

图8 SPR分析突变单链抗体与CmBBMV的结合Fig. 8 SPR analysis of mutant scFv antibodies binding to CmBBMV
2.8 突变单链抗体对稻纵卷叶螟的生物活性
稻纵卷叶螟幼虫经过24 h处理,Y124G与scFv-C7杀虫效果无明显差异,但是48 h后,特别是经过72 h处理,Y124G将scFv-C7的死亡率从42.22%提高到56.67%,虽然突变型将杀虫活性提高,但是显著低于阳性对照Cry1B的76.67%(表3),还需进一步改造以达到更佳效果。
表3 突变单链抗体对稻纵卷叶螟的生物测定Table 3 Bioassay of mutant scFv antibodies against C. medinalis larvae
3 讨论
定点突变的改造方法与传统抗体改造方法相比,避免了盲目性,成功率较高,而且以分子模拟为基础的定点突变,在已知蛋白三维结构及功能等方面信息的基础上,预测分子间结合区域并确定关键氨基酸残基,更加准确的反应了抗体蛋白作用界面的特征,大大提高了预测准确度[13,24-25]。本文利用分子对接、预测热点的理论知识,构建饱和突变抗体库,经过固相筛选,准确发现了一株提高功能活性的突变株。
计算机辅助设计所构建的饱和突变抗体库的库容为2×103,库容偏小。在设计饱和突变片段的时候,突变位点设计成 NNK(N为A、G、C 或T,K为G或T),这使每个位点能囊括全部20种氨基酸,有效减少同义密码子的数量,并能有效减少突变抗体库的筛选数量[26-27]。此外,为了提高转化效率,我们使用了电转化来构建抗体库。库容偏小可能的原因是在扩增饱和突变基因过程中得到的PCR产物偏少或者浓度偏低(图4-D),这需要优化PCR程序或设计新的引物。或者可在既有突变体的基础上进行多轮突变,以积累有益突变,从而获得亲和力更高的突变体。
SPR技术是基于金属薄膜的光学耦合作用原理建立的探测生物分子之间相互作用的新技术,可简单快捷地监测生物分子之间的相互作用[28-30]。BIAcore SPR已在许多监测种类中应用,包括“抗原-抗体”和“受体-配体”互作这两种最常见的类型。SPR分析发现Anti-Id scFv与CmBBMV的结合是“结合-平衡-解离”的动态过程,不同于ELISA结合实验只得到最终状态的静态结果。一般只有偶联到CM5传感芯片上蛋白是单一纯化物质才能计算动力学常数[31-32],由于昆虫中肠BBMV是多种受体蛋白混合而成,所以无法算出动力学常数,但是可以计算出结合能力。SPR与ELISA结果相互印证,明确了突变株Y124G对CmBBMV的亲和力比野生型scFv-C7提高(图8)。
虽然突变型Y124G对稻纵卷叶螟的杀虫活性较野生型scFv-C7提高,但还是显著低于Cry1B毒素(表3)。随着蛋白质晶体结构的增加及优化,三维结构数据库的扩充,模型评价系统的优化,分子对接技术的升级,从而可以获得更准确的对接模型[33]。进而在模型结构上可以精确分析接触面残基位点,确定突变氨基酸,为得到活性提高的抗体蛋白材料奠定了坚实的基础。
4 结论
本研究采用计算机模拟方法构建了scFv-C7和CmAPN的结构模型,分子对接并预测对接复合物的关键结合区域,分析位于scFv-C7上的热点残基并构建定点饱和突变抗体库,利用固相筛选方法包被CmBBMV,筛选出亲和力和生物活性提高的突变单链抗体。对基因工程抗体分子对接热点进行定点饱和突变,是一种快速抗体分子改造的有效方法,也为蛋白农药制备改造提供了一个新途径。
