诱导子对雷公藤悬浮细胞中雷公藤甲素积累的影响
2021-11-18朱留刚吴承祯
张 强,宋 萍*,朱留刚,封 磊,洪 伟,3,吴承祯
(1.福建农林大学 林学院,福建 福州 350002;2.福建农林大学 资源与环境学院,福建 福州 350002;3.福建省高校森林生态系统过程与经营重点实验室,福建 福州 350002;4.武夷学院,福建 武夷山 354300)
【研究意义】雷公藤(Tripterygium wilfordiiHook.f.)隶属卫矛科雷公藤属,广泛用于治疗类风湿性关节炎、肺病、肾病、皮肤病、白血病等疾病,是珍贵的药用植物[1]。雷公藤提取物中的二萜三环氧化物雷公藤甲素(triptolid)是其主要的药用活性成分之一。1972年,Kupchan等[2]首次发现雷公藤甲素具有明显的抗白血病作用,此后雷公藤甲素又被陆续发现可对多种肿瘤细胞株产生明显的抑制作用;此外,雷公藤甲素还具有免疫抑制和抗炎作用,对多种促炎细胞因子和介质以及内皮细胞粘附分子的表达具有抑制作用,并具有诱导自身或其它细胞凋亡的能力[3]。因此,雷公藤甲素具有广阔的开发前景与应用市场。但由于其结构十分复杂和特殊,目前尚无法有效地进行大量人工合成,加之其在植株中的天然含量低、分离困难,使得对于雷公藤甲素的研究和应用受到很大限制。植物细胞悬浮培养技术由于可以人工控制实现细胞生长、分化和次生代谢产物积累的最佳条件,从而可以有效地促进细胞增殖并定向诱导次级代谢产物的合成和积累。因此,研究如何利用细胞悬浮培养技术生产雷公藤甲素具有重要意义。【前人研究进展】利用诱导子来提高次生代谢物的产量,是目前植物悬浮细胞培养中常用的方法。诱导子可分为生物诱导子与非生物诱导子两种。生物诱导子包括来自真菌、细菌或草食动物诱导子、植物细胞壁碎片、植物被病原体或草食动物入侵时释放的化学物质等,其功能与受体偶联,通过激活或失活许多酶或离子通道起作用[4]。以酵母提取物及榆黄蘑作为生物诱导子,添加至怀牛膝悬浮细胞培养物中,能明显促进牛膝多糖的生成[5];利用青霉和黑曲霉作为生物诱导子,可分别将大豆悬浮细胞中异黄酮的积累量提升2.56倍与1.32倍[6]。近年来,植物内生菌被发现可以作为生物诱导子用于提高植物细胞培养过程中次生代谢产物的积累。如高媛等[7]通过添加内生真菌诱导子使苦豆子中喹诺里西啶类生物碱达到了对照组的2.65 倍;李群等[8]将6 株内生菌作为生物诱导子添加于灰毡毛忍冬悬浮细胞培养体系内,其对细胞生物量及次生代谢产物绿原酸含量产生了不同程度的促进作用。非生物诱导子是指本身并不是植物细胞中的天然成分,却可以通过化学或物理途径使植物产生防御反应,进而触发植物次生活性物质的合成。其种类主要包括植物生长调节剂、水杨酸、茉莉酸甲酯、重金属盐、稀土元素、紫外线辐射、高低温和溶氧量等[4]。如利用茉莉酸甲酯和水杨酸作诱导子使半夏悬浮细胞中的生物碱含量分别达到对照组的3.6和2.5倍[9]。【本研究切入点】截止目前,已有多种诱导子被用于包括人参、雪莲、红豆杉、黄芪等在内的药用植物悬浮培养体系中,并在细胞生长及活性成分积累方面起着重要作用,然而有关雷公藤悬浮细胞中雷公藤甲素积累的研究鲜少涉及。前期研究发现,雷公藤甲素可以在雷公藤愈伤组织[10]和悬浮细胞[11]中积累。本研究拟通过雷公藤细胞悬浮培养,以水杨酸(salicylic acid,SA)、稀土元素La3+以及雷公藤内生真菌为诱导子,研究其对雷公藤悬浮细胞中雷公藤甲素积累的影响。【拟解决的关键问题】本研究通过制备和添加不同的非生物和生物诱导子,揭示诱导子对雷公藤悬浮细胞产雷公藤甲素的影响,以期提高悬浮细胞中雷公藤甲素含量,为雷公藤甲素的扩大化生产提供基础。
1 材料与方法
1.1 雷公藤细胞悬浮培养
以雷公藤植株嫩叶为外植体诱导愈伤组织,连续继代培养3 代以上。选取质地疏松、生长良好的愈伤组织,用镊子夹成小块,称取0.4 g进行接种[14]。应用White基础培养基(青岛海博生物技术公司),附加激素2,4-D 1.0 mg/L、NAA 0.1 mg/L和KT 0.1 mg/L,培养基经121 ℃灭菌20 min。将愈伤组织接种到装有50 mL培养基的三角瓶中,置于恒温振荡摇床于120 r/min、(25±1)℃下悬浮暗培养。
1.2 非生物诱导子的添加
雷公藤细胞悬浮培养至第3 天向培养液中添加非生物诱导子。使用医用注射器分别吸取SA 和LaCl3原液,经0.22 μm 的灭菌水系微孔滤头过滤除菌后添加至细胞培养液中。SA 添加后的质量浓度分别控制在0.1 mg/L(低浓度)、1.0 mg/L(中等浓度)和10 mg/L(高浓度);LaCl3添加后的浓度分别控制在20 mg/L(低浓度)、40 mg/L(中等浓度)和60 mg/L(高浓度)。对照不添加任何诱导子。
1.3 内生真菌诱导子的制备
利用本实验室保藏的两株雷公藤内生真菌Pochonia suchlasporiaNS-17和Penicillium spinulosumNS-5 进行生物诱导子的制备。将菌株接种于PDA 平板进行活化,取菌饼接种于PDA 液体培养基,于28 ℃、120 r/min 下摇床震荡培养6 d,对培养物进行抽滤分别收集菌丝体和发酵液滤液。菌丝体用蒸馏水充分清洗后,加入蒸馏水进行匀浆,再次抽滤,得到菌丝提取物,于121 ℃灭菌20 min,即得菌丝体诱导子。将发酵液滤液高压灭菌制备成培养液诱导子。在植物细胞悬浮培养至第4、8天(细胞分别处于生长前期和对数生长期)时,分别添加各内生真菌诱导子。以诱导子的糖浓度来确定其添加量,糖含量应用苯酚硫酸法测定。
1.4 细胞活力的测定
采用氯化三苯四氮(2,3,5-Triphenyltertrazolium Chloride,TTC)还原法测定悬浮细胞活力。
1.5 细胞干质量的测定
用超纯水将悬浮细胞充分清洗,去除残留的培养基,抽滤,60 ℃烘干至恒量,用电子天秤称重,记为干质量。
1.6 总糖测定
悬浮细胞培养液中总糖的含量采用苯酚硫酸法测定。
1.7 苯丙氨酸解氨酶(PAL)活性的测定
细胞PAL的活性依据PAL与L-苯丙氨酸的反应产物肉桂酸在290 nm处的吸光值的变化进行测定。
1.8 雷公藤甲素的测定
雷公藤甲素应用高效液相色谱仪(HPLC)测定。通过过滤分离培养液和悬浮细胞,其中细胞样品烘干至恒重,用研钵碾成粉末。称取粉末0.500 0 g。以甲醇为浸提液,超声处理30 min,4 ℃自然浸提12 h。然后4 000 r/min 离心,将上清液定容于10 mL 容量瓶,经0.45 μm 滤头过滤后作为待测液。HPLC 色谱柱为XB-C18 柱(5 μm,4.6 mm×250 mm);流动相为水/甲醇(30/70);流速是1.0 mL/min;波长为218 nm;柱温是室温。进样量为10 μL[13]。每个样品重复3次。
2 结果分析
2.1 不同诱导子对雷公藤甲素产量的影响
由图1,SA处理下的雷公藤细胞悬浮培养体系中雷公藤甲素含量与诱导子浓度有关,0.1 mg/L的SA明显促进了雷公藤甲素的产量,在培养12 d时培养液中雷公藤甲素含量最高,约是对照的3.5倍。而1.0 mg/L和10 mg/L的SA对雷公藤甲素积累的影响很小。

图1 诱导子对雷公藤甲素含量的影响Fig.1 The effect of elicitors on the content of triptolide
相比对照,3种浓度的LaCl3诱导子均明显提高了培养液中雷公藤甲素的产量,且诱导子浓度越低其诱导效应越强;但诱导子浓度越低,雷公藤甲素产量峰值出现的越晚。20,40,60 mg/L 的LaCl3诱导下的雷公藤甲素浓度峰值分别是对照的2.0、1.7和1.2倍。由此可见,低浓度的SA和不同浓度的LaCl3均能促进雷公藤悬浮细胞向外释放雷公藤甲素,而LaCl3的诱导效应随浓度的增加而减弱。
与对照相比,在细胞培养第4天添加内生真菌诱导子,NS-5菌丝体和NS-17菌丝体均显著促进了悬浮细胞培养体系中雷公藤甲素的积累,培养12 d 时,雷公藤甲素产量最高,分别是对照的1.9 和1.7 倍。NS-17培养液和NS-5培养液对雷公藤甲素产量的影响则相对较小。
在细胞培养第8 天添加内生真菌诱导子,在诱导初期各诱导子便提高了细胞中雷公藤甲素的释放;随后,NS-17培养液和NS-5菌丝体诱导下雷公藤甲素在培养第15天达到峰值,其产量分别是对照的2.0和2.4 倍;NS-17 菌丝体、NS-5 培养液诱导下的雷公藤甲素产量分别在培养第9、12 天时最大,分别是对照的2.1和1.8倍。
2.2 不同诱导子对细胞活力的影响
如图2,将不同浓度的非生物诱导子SA 和LaCl3添加至培养3 d的雷公藤细胞悬浮培养体系后,细胞活力随着培养时间的增加呈现先增强后减弱的趋势。与对照相比,两种非生物诱导子的添加均使细胞活力的峰值提前。中、低浓度的SA(1.0 mg/L、0.1 mg/L)和LaCl3(40 mg/L、20 mg/L)在悬浮培养前期对细胞活力具有明显的诱导效应,诱导第3 天和第6 天时,细胞活力高于对照;培养12 d 之后,这些诱导子的诱导效应减弱,各处理的细胞活力接近于对照。与20 mg/L 的LaCl3相比,40 mg/L 的LaCl3提高细胞活力的效应更强。高浓度的SA和LaCl3处理的悬浮细胞活力均低于对照,显示出一定的抑制效应。

图2 诱导子对细胞活力的影响Fig.2 The effect of elicitors on the cell viability
在雷公藤细胞悬浮培养前期添加内生真菌诱导子后,在培养前12 d 内,NS-5 培养液显著提高了细胞活力,NS-5菌丝体对细胞活力的影响则不明显;在培养后期,NS-5培养液和菌丝体对细胞活力均产生了抑制作用(图1)。除了诱导初期,从培养至第9天开始,NS-17培养液和菌丝体对悬浮细胞活力均显示出明显的抑制效应。在细胞对数生长期添加内生真菌诱导子后,相比对照,NS-5 培养液在细胞对数生长期(9 d、12 d)提高了细胞活力,但在细胞生长后期对细胞活力产生了抑制作用;其它诱导子在整个诱导期对细胞活力均具有抑制作用。
2.3 不同诱导子对细胞生长的影响
对非生物诱导子和生物诱导子处理下的雷公藤悬浮细胞生长情况测定(图3)发现,各诱导子处理下雷公藤悬浮细胞的生长趋势与对照基本一致。0.1 mg/L SA 处理的悬浮体系细胞干质量大于对照;10 mg/L SA 在细胞生长前期干质量高于对照而进入生长对数期以后则低于对照;1.0 mg/L SA 则明显降低了细胞干质量的积累。与对照相比,20 mg/L 和40 mg/L LaCl3处理对细胞干质量的积累无明显的影响;60 mg/L LaCl3则明显提高了对数生长期和生长后期的细胞干质量。
在细胞生长前期添加内生真菌诱导子能够促进细胞的生长,提高细胞的干质量(图3)。其中,NS-17 培养液诱导子的促进作用最显著,12 d 时的细胞干质量达最大,是对照的2.8 倍;其次是NS-5 培养液诱导子,细胞干质量在9 d时达最大,是对照的1.9倍;NS-5菌丝体和NS-17菌丝体诱导子对细胞干质量呈现出轻微的促进作用。在细胞培养第8 天添加内生真菌诱导子后,NS-5 培养液在整个诱导期均提高了细胞干质量;NS-17 培养液在细胞对数生长期显著促进了干质量的积累;而NS-5 菌丝体和NS17 菌丝体的诱导作用不大,细胞干质量与对照相接近。由此可见,细胞生长前期添加内生真菌诱导子有利于提高细胞生物量,内生真菌培养液比菌丝体的促进作用要强,其中NS-17培养液的诱导作用最强。

图3 诱导子对细胞生长的影响Fig.3 The effect of elicitors on the cell growth
2.4 不同诱导子对总糖消耗的影响
由图4中SA 处理下的雷公藤悬浮细胞培养液中总糖含量的变化可知,0.1 mg/L 和10.0 mg/L SA 诱导下细胞对总糖的消耗明显高于对照,而1.0 mg/L SA 诱导的细胞总糖消耗则略低于对照。SA 诱导的细胞总糖消耗与生物量的增加基本一致。根据LaCl3诱导的培养液总糖含量变化曲线,20 mg/L LaCl3诱导下细胞对培养液中总糖消耗明显高于对照,这可能与雷公藤甲素的积累有一定关联;60 mg/L 诱导下的细胞总糖消耗也明显高于对照,总糖的消耗与细胞生物量的提高相一致;40 mg/L LaCl3诱导下的细胞总糖消耗则稍低于对照。
在细胞生长前期添加内生真菌NS-5、NS-17 诱导子后,培养液中总糖含量均低于对照(图4)。其中,NS-17培养液诱导的细胞总糖消耗最大。总体上,各内生真菌诱导子作用下的培养液总糖消耗与细胞生物量增长相一致,而两者与雷公藤甲素的积累没有明显关联。在细胞培养至对数生长期添加内生真菌诱导子与生长前期添加的培养液总糖含量变化十分接近。

图4 诱导子对培养液中总糖含量的影响Fig.4 The effect of elicitors on the total sugar content in the culture broth
2.5 不同诱导子对细胞PAL活性的影响
非生物诱导子处理下的雷公藤悬浮细胞PAL 活性变化如图5 所示,细胞PAL 活性随诱导时间的增加明显增强,在对数生长期(第9 天)达到峰值,之后逐渐减弱。相比对照,3 种浓度的SA 诱导子均明显剌激了细胞PAL 活性,并且随着SA 诱导子浓度的增加,剌激作用越强。3 种浓度的LaCl3在细胞对数生长期对PAL活性具有增强作用,在细胞生长后期作用不明显,其中,40 mg/L的LaCl3处理的细胞PAL活性峰值最大。
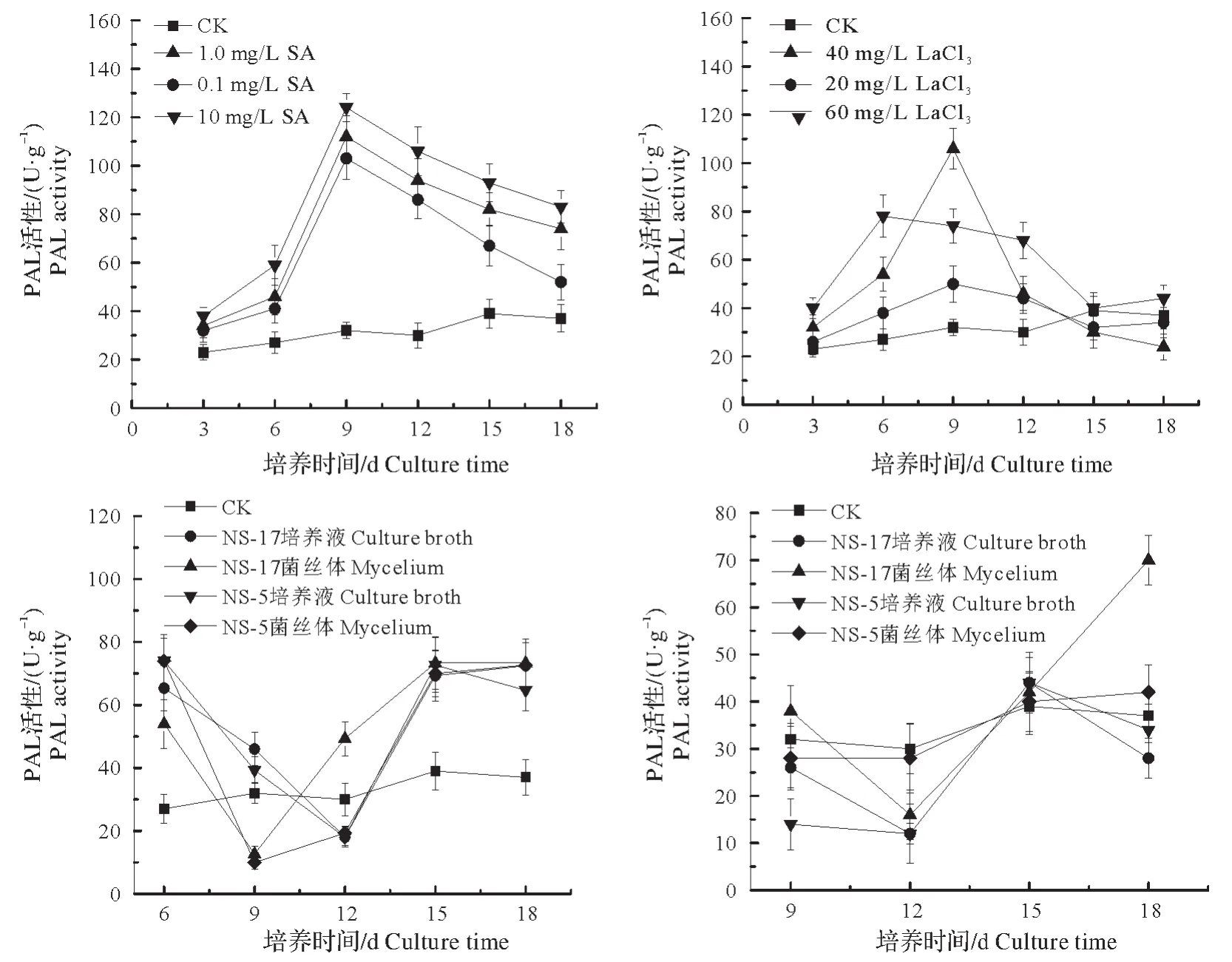
图5 诱导子对PAL活性的影响Fig.5 The effect of elicitors on the PAL activity
细胞生长前期添加内生真菌诱导子后,雷公藤悬浮细胞PAL活性随培养时间的增长呈现先减弱后增强的趋势,细胞对数生长期PAL达到低谷。与对照相比,各内生真菌诱导子在细胞生长前期和后期提高了PAL 活性,而在对数生长期则降低了PAL 活性。在细胞对数生长期添加内生真菌诱导子后,相比对照,NS-17菌丝体诱导子在细胞生长前期和后期提高了PAL活性,而在对数生长期抑制了PAL活性;NS-17和NS-5培养液诱导子在整个诱导期对细胞PAL显示了抑制作用;NS-5菌丝体诱导子对PAL的影响不明显。
3 讨论与结论
SA 是一种广泛存在于植物体内的酚类化合物,具有多种功能,包括调控植物的生长发育以及在生物和非生物胁迫条件下诱导植物的防御反应等,其作为诱导子可以有效地促进药用植物次生代谢产物的合成和积累。如张媛媛等[12]发现添加200 μmol/LSA 培养30 d 时,半夏中肌苷含量高出对照组161.40%。水杨酸、萘乙酸和硝普钠3种诱导子均可以促进蒙古黄芪有效成分的积累,其中,水杨酸对提高黄酮成分的效果最为明显,萘乙酸与硝普钠则分别对多糖与皂苷的积累具有显著的促进作用[13]。SA不仅对同一细胞系内不同化合物的影响不同,对同一细胞系同一化合物的效应大小也受其添加浓度的影响。本研究中,低浓度的SA(0.1 mg/L)明显提高了悬浮细胞培养体系中雷公藤甲素的产量,最高为对照的3.5倍。同时,低浓度SA的添加提高了细胞活力、细胞生物量积累和总糖消耗量。而高浓度SA(10 mg/L)对雷公藤甲素积累的影响很小。可见,SA 可以有效诱导雷公藤甲素的积累,而雷公藤甲素的积累与细胞生长呈现一定的偶联。较低浓度的外源SA 处理可通过上调防御相关基因表达来诱导各种重要的次生代谢产物[14-15]。低浓度SA(25 μmol/L、50 μmol/L)诱导的颠茄毛状根东莨菪碱质量分数较对照显著增加[16]。而9 mmol/L SA 和12 mg/L 壳聚糖诱导下的宽丝獐牙菜(Swertia paniculata)茎叶培养物中裂环烯醚萜和黄酮类酣含量最高,更高或更低的诱导子浓度则会使这些化合物的含量降低[17]。
镧作为一种重要的稀土元素,对植物的生长发育和抗逆性等都具有一定的影响,其常作为非生物诱导子用于植物次生代谢产物的合成和积累。如添加20.00 mg/L的LaCl3溶液可使铁皮石斛中石斛碱含量增加24.53%[18]。稀土元素对植物生理生化的正、负效应取决于剂量和其它条件,通常,在较低浓度下可刺激细胞生长和酶活性,但在较高浓度下对细胞有毒[19]。本研究中,不同浓度的LaCl3均提高了悬浮细胞培养物中雷公藤甲素含量,随着LaCl3浓度的降低诱导作用增强,低浓度LaCl3诱导下的细胞总糖消耗明显增加,但低浓度的LaCl3对雷公藤细胞活力和细胞生长的影响较小,反映雷公藤甲素的积累可能不是细胞增长的结果,但可能与细胞糖代谢有一定的关联。本研究的结果与其它研究有着相似之处,在云南红豆杉细胞培养中,硝酸镧浓度为1.15~23 μmol/L时,促进了胞外紫杉醇的生产和胞内紫杉醇的合成,其中5.8 μmol/L 硝酸镧的诱导效应最强,在某些低浓度下硝酸镧对紫杉细胞生长显示出轻微的正效应,而更高浓度的硝酸镧(23.1~46.2 μmol/L)诱导作用较弱[19]。可见,镧可以用于诱导雷公藤悬浮细胞培养体系中雷公藤甲素的生产,但应对其剂量和其它条件进行有效控制。
植物内生真菌诱导子能够快速、高效和专一地活化细胞特定次生代谢途径,已广泛用于提高植物次生代谢产物的合成和积累,如苦豆子内生真菌菌液浓缩物使苦豆子组培苗中氧化苦参碱含量得以提高,最高为对照组的20.9倍[20]。齐凤慧等[21]在茶条槭悬浮细胞培养的对数期加入内生真菌(Phomopsissp.)诱导子,使茶条槭细胞中没食子酸的含量得到明显提升,进一步研究发现该内生真菌诱导子可能通过增大细胞膜透性,促进细胞核有丝分裂,提高无机盐离子吸收等多个方面调控没食子酸的合成。真菌诱导子中具有诱导活性的成分主要是其所含有的多糖类物质,能够刺激细胞代谢产物的合成。本研究中,在悬浮细胞培养前期添加内生真菌诱导子,NS-5菌丝体和NS-17菌丝体诱导子均明显促进了雷公藤甲素的生产,两者对细胞生长和总糖消耗呈现轻微的正效应,雷公藤甲素的积累与细胞生长没有明显的关联;在细胞培养至对数生长期添加各诱导子均提高了雷公藤甲素的含量,最高是对照的1.8~2.4倍。无论是细胞培养前期添加还是对数生长期添加,NS-5 菌丝体提取物的诱导作用均是最强的。可见,内生真菌可作为诱导子用于悬浮细胞雷公藤甲素的生产,而诱导效应的大小即与内生真菌诱导子和添加时间有关。不同添加时间对活性产物合成的影响可能与细胞的代谢活动有关。李萍等[5]指出,在细胞生长前期细胞还没有形成足够的前体用于合成代谢需要的各种酶;在对数生长期,细胞代谢活动旺盛,对诱导子比较敏感;在生长迟缓期,细胞代谢水平下降,诱导子对代谢产物的合成影响很小,因此在细胞对数生长期添加诱导子往往诱导效应更强。
PAL是苯丙烷类途径中第一个酶,也是连接初生代谢和次生代谢的关键酶,对植物的生理状态十分敏感,PAL活性的激活是植物细胞受到生物和非生物胁迫常见的反应[22]。在红豆杉细胞培养物中,各种生物和非生物因子,如重金属离子、真菌诱导子、茉莉酸甲酯和机械应力等的诱导提高了PAL活性[23-25]。本研究中,SA及LaCl3的添加明显提高了雷公藤悬浮细胞PAL活性,在对数生长期PAL活性最大;而内生真菌诱导子的添加则使PAL活性呈现先增后降再增加的变化,对数生长期PAL活性呈现抑制状态。SA及LaCl3添加对PAL活性明显的促进作用表明,SA 及LaCl3诱导的PAL活性直接或间接上调可能是悬浮细胞培养体系中雷公藤甲素产量得以提高的原因之一。而内生真菌诱导子促进雷公藤甲素在悬浮细胞培养体系内积累的同时,PAL活性并未明显升高,由此推测内生真菌诱导的雷公藤甲素的合成可能与PAL酶无关。
综上所述,不同诱导子对雷公藤悬浮细胞中雷公藤甲素积累均具有一定的促进作用。本研究中0.1 mg/L SA提高雷公藤悬浮细胞活力、细胞生物量、总糖消耗量及PAL活性的同时促进了雷公胶甲素的积累,最高含量为对照的3.5 倍;LaCl3对悬浮细胞培养物中雷公藤甲素积累的诱导作用随着其浓度的降低而增强,最高含量为对照的2.0倍;内生真菌诱导子中的NS-5菌丝体提取物对雷公藤甲素积累的诱导效果最好,最高含量是对照2.4倍。本研究的结果可为雷公藤甲素的扩大化生产奠定基础。
