草地贪夜蛾不同季节取食玉米叶和大豆叶生活史特性比较
2021-11-18黄丽莉薛芳森何海敏
刘 伟,徐 强,黄丽莉,薛芳森,何海敏*
(1.江西农业大学 昆虫研究所,江西 南昌 330045;2.上饶职业技术学院 管理工程系,江西 上饶 334109;3.豫章师范学院 生态与环境系,江西 南昌 330103)
【研究意义】草地贪夜蛾Spodoptera frugiperda(J.E.Smith)是起源于美洲热带和亚热带地区的多食性害虫,也是重大的迁飞性害虫[1-3]。2019年5月上旬草地贪夜蛾首次迁飞到江西省,2020年4月中旬在江西南部信丰县(25.4°N,114.9°E)首次发现为害,发生期比2019年提早了20 d。2020年8月下旬在南昌市乐化镇玉米地调查草地贪夜蛾时发现,在没有采取任何防治措施的田块,其玉米植株100%受害。幼嫩组织,叶、茎、穗和花丝均遭受严重危害,几乎看不到一片完整的叶片。在江西大部分地区,玉米与大豆邻作较为普遍,即有玉米种植的地方附近也种植了大豆,且两种作物在全年出现的时间基本一致。然而,在玉米与大豆邻作的田块和其他田块中调查发现,草地贪夜蛾仅趋向于玉米上产卵和为害,没有在大豆上产卵和为害。探明新入侵草地贪夜蛾对大豆是否具有潜在风险,为该虫发生为害的预测和防治对策的制定提供一定的理论依据。【前人研究进展】2016 年1 月草地贪夜蛾首次发现在西非的尼日利亚和加纳暴发为害[4],随后迅速扩散到撒哈拉以南非洲的大部分地区,给玉米生产造成了巨大的损失[5]。草地贪夜蛾没有滞育能力,在北美温带地区不能以任何虫态越冬。在美国草地贪夜蛾越冬地区是佛罗里达南部和德克萨斯州南部[1,6]。遗传分析表明,草地贪夜蛾可分为玉米型和水稻型两个种群,这两个种群在遗传上出现明显分化,但形态上仍相同,且两个种群均能取食玉米[7-8]。2019年1月草地贪夜蛾玉米型种群首次迁入我国云南省江城县,在玉米和甘蔗上发生危害[9]。尽管目前报道草地贪夜蛾的寄主植物种类有353 种,分属76 科,禾本科占106 种,豆科和菊科占31 种[10],但在江西受草地贪夜蛾为害的作物主要是玉米,一些地方的甘蔗也受其为害。然而在非洲加纳和肯尼亚的大豆田,大豆受到草地贪夜蛾严重为害[5]。昆虫的生活史特性主要包括发育历期、生长速率、体质量、寿命和繁殖力等,这些生活史特性是衡量一种昆虫对环境适应性的重要参数,也能反映了入侵昆虫对当地环境的适应及其生存能力[11]。在田间自然条件下,不同季节具有不同的气候因子,气候因子被认为是影响昆虫生长发育、存活、繁殖等生命活动最重要的因子之一,而温度是影响昆虫生活史特性最重要的气候因子。一般来说,在适合昆虫生存的温度范围内,温度的升高会增加代谢速率,从而导致更高的生长速度、更短的发育时间和更小的成虫体型,亦称之为温度-体型大小法则(the temperature-size rule)[12-13]。动物在雌雄体型间常常存在差异,这种现象称为性体型二型性(sexual size dimorphism)。现有研究表明,88%的昆虫雌性大于雄性,只有7%的昆虫是雄性大于雌性[14]。寄主植物也是影响昆虫生活史特性的重要因子。对于可取食为害多种植物的昆虫来说,由于寄主植物所含营养成分或次生代谢物质的差异,取食不同寄主植物可以对其生长发育、繁殖力和种群增长产生影响[15-17]。【本研究切入点】目前,国内有关草地贪夜蛾取食玉米、甘蔗、水稻、烟草、花生、荞麦、生姜、小麦、高粱、烟草、臂形草等不同寄主植物对其适应性的影响已有不少报道[18-25]。这些研究为了解草地贪夜蛾取食不同寄主植物的适应性提供了参考资料。然而上述研究均未分雌雄进行幼虫个体发育的观察(仅吴正伟等[20]报道了雌雄蛹质量存在显著差异),观察的样本数也较少,观察到的产卵量也相对较低。本研究对草地贪夜蛾的生活史特性(life-history traits)进行了较细致和全面的研究,测定了每个雌雄个体从卵孵化到化蛹和化蛹至成虫羽化的时间、蛹和成虫的质量、生长速率,记录了幼虫和蛹的存活率以及性比,计算了蛹发育到成虫丢失的质量比率,观察了已交配雌虫的产卵前期、产卵期、产卵量和卵的孵化率。【拟解决的关键问题】通过系统比较草地贪夜蛾在不同季节取食玉米和大豆的生活史特性,明确季节和寄主植物对草地贪夜蛾生活史特性的影响,探明该入侵害虫草地贪夜蛾的食物适应机制,为该虫种群发生动态的预测预报及其防治策略的制定提供基础资料。
1 材料与方法
1.1 供试虫源和饲养材料
草地贪夜蛾于2020年采自江西南昌乐化镇奶牛场玉米地(28°68′N,115°89′E)。供试寄主植物玉米Zea mays和大豆Glycine max品种分别为‘甜糯182’和‘中黄37’。在江西农业大学试验田种植,均为常规水肥管理且不施用农药。
1.2 饲养方法
所采集草地贪夜蛾在通风透光的养虫室(面积20 m2)自然条件下继代饲养繁殖,收集卵块供试验用。在本试验中,将夏初6月14日、夏中7月7日和秋中9月4日孵化的幼虫分别置于盛有湿润卫生纸的塑料盒(16 cm×11 cm×5.5 cm)中,在自然条件下用已完全展开的新鲜玉米叶和大豆叶饲养,每个处理接虫3~5 盒,每盒不少于50 头。待幼虫发育至2 龄后转为每个培养皿(高2.0 cm,直径9.0 cm)单头饲养直至化蛹和羽化。根据取食情况及时更换新鲜叶片。养虫室内用温度记录仪(鲜盾1000B)每隔1 h记录温度。在夏初、夏中和秋中这3个不同季节的试验中,生活史特性的日平均气温是指幼虫孵化到成虫羽化的温度(夏初、夏中和秋中孵化的幼虫所经历的日平均气温分别为28.6,29.1,24.0 ℃),繁殖特性的日平均气温是指成虫羽化到成虫死亡的温度(夏初、夏中和秋中所经历的日平均气温分别为28.2,30.9,21.4℃)。
1.3 生物学特征观察
对草地贪夜蛾上述3 个时期孵化的每个个体,测定了其卵孵化到化蛹和化蛹至成虫羽化的时间、蛹和成虫的质量、生长速率。记录了每种温度下幼虫和蛹的存活率以及性比。计算了蛹发育到成虫丢失的质量比率。幼虫化蛹后,第2 天置于电子天平(AUY120 日本岛津公司)称量蛹质量,成虫于排出蛹便后称量。成虫羽化后,立即进行配对,置于充入空气的塑料袋(20 cm×30 cm)中交配产卵,每袋只放1对,袋中放入用10%蜂蜜水湿润的棉球供成虫补充营养,每种温度下雌雄配对不少于26 对。每天收集卵块,详细记载已交配雌虫的产卵前期、产卵期、产卵量和卵的孵化率。交配成功率是指配对的雌虫产下可育卵。孵化率是指每种温度下雌虫产下的所有卵孵出幼虫的比例。每天更换塑料袋和用蜂蜜水湿润的棉球,直至成虫死亡。
1.4 数据分析
生长速率的计算方法:生长速率=ln 蛹质量/幼虫期[26]。蛹和成虫间的失重比计算方法:失重比=l-(成虫质量/蛹质量)[26]。
试验数据采用SPSS 17.0 软件进行分析。运用混合线性模型分析了寄主、季节和性别对生活史特性的影响。寄主、季节和性别间生活史特性差异比较作为混合单因素,采用one-way ANOVA 和Duncan 氏多重比较检验,寄主间繁殖特性的差异比较采用独立样本t检验。不同季节下采用非参数检验中的二项分布检验方法,比较其性别间是否显著偏离1∶1。
2 结果与分析
2.1 与食料、季节和性别相关的发育历期
食料、季节和性别显著影响了草地贪夜蛾幼虫的发育历期,食料×季节、食料×季节×性别的互作对幼虫的发育历期也有显著影响(表1)。幼虫的发育历期显著受到季节的影响(F11,1059=652.755,P<0.05)(图1A)。在不同季节,取食玉米叶的幼虫发育历期均显著短于取食大豆叶的幼虫发育历期(P<0.05)。在日平均气温28.6 ℃的夏初,雌性幼虫期缩短了5.4 d,雄性幼虫期缩短了5.1 d;在日平均气温29.1 ℃的夏中,雌性和雄性幼虫期均缩短了2.6 d;在日平均气温24.0 ℃的秋中,取食玉米叶的雌性幼虫的发育历期缩短了4.8 d,雄性缩短了4.6 d(表2)。在夏初和夏中,取食玉米叶雌幼虫期显著短于雄幼虫期(P<0.05),在秋中取食玉米叶雌雄幼虫期无显著差异(P>0.05);在夏中和秋中,取食大豆叶雌幼虫期显著短于雄幼虫期(P<0.05),在夏初取食大豆叶雌雄幼虫期无显著差异(P>0.05)(表2)。
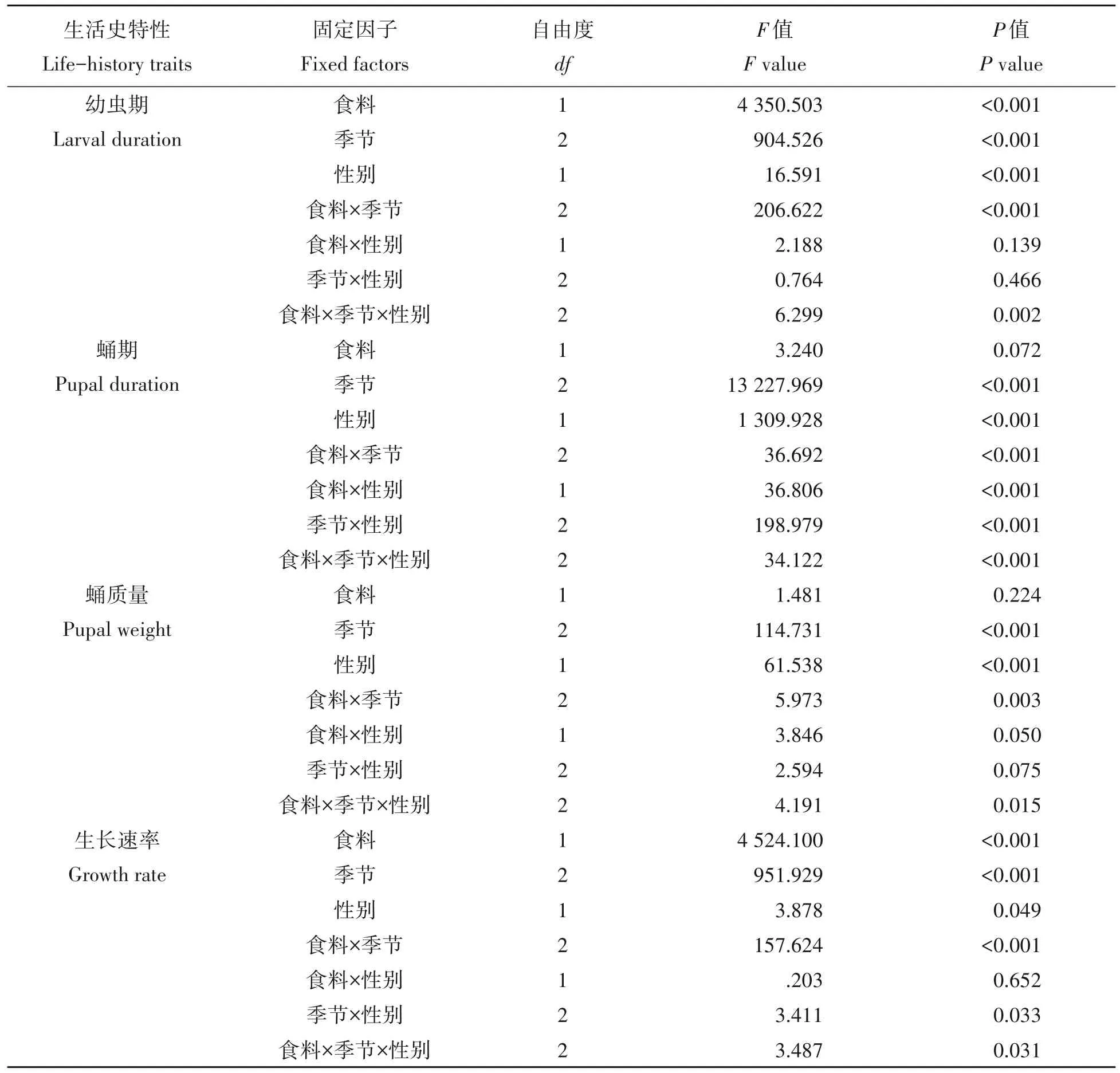
表1 食料、季节和性别对草地贪夜蛾幼虫期、蛹期、蛹质量和生长速率影响的多因素方差分析Tab.1 Multi-factor ANOVA analysis of food,season and sex on larval duration,pupal duration,pupal weight and growth rate in Spodoptera frugiperda
蛹的发育历期也受到季节和性别及它们互作(季节×性别)的显著影响;蛹期没有受到食料的显著影响,但食料×季节、食料×性别和食料×季节×性别的互作显著影响了蛹期(表1)。蛹发育历期在不同季节存在显著差异(F11,1059=2 605.328,P<0.05)(图1B)。在不同季节,取食玉米叶和大豆叶的蛹期没有呈现有规律的变化。在夏初日平均气温28.6 ℃时,取食玉米叶的雌雄蛹期均短于取食大豆叶的(P<0.05);在夏中日平均气温29.1 ℃时,取食玉米叶的雌雄蛹期显著长于取食大豆叶的(P<0.05);在秋中日平均气温24.0 ℃时,取食玉米叶的雌蛹期显著长于取食大豆叶的(P<0.05),但取食玉米叶的雄蛹期显著短于取食大豆叶的(P<0.05)(表2)。无论取食玉米叶和大豆叶,雄性的蛹期均显著长于雌性(P<0.05),导致了雌性先熟现象(protogyny phenomenon)。

图1 草地贪夜蛾在夏初、夏中和秋中取食玉米叶和大豆叶的发育历期Fig.1 Development time of Spodoptera frugiperda feeding on corn leaves and soybean leaves in early summer,mid-summer and mid-autumn
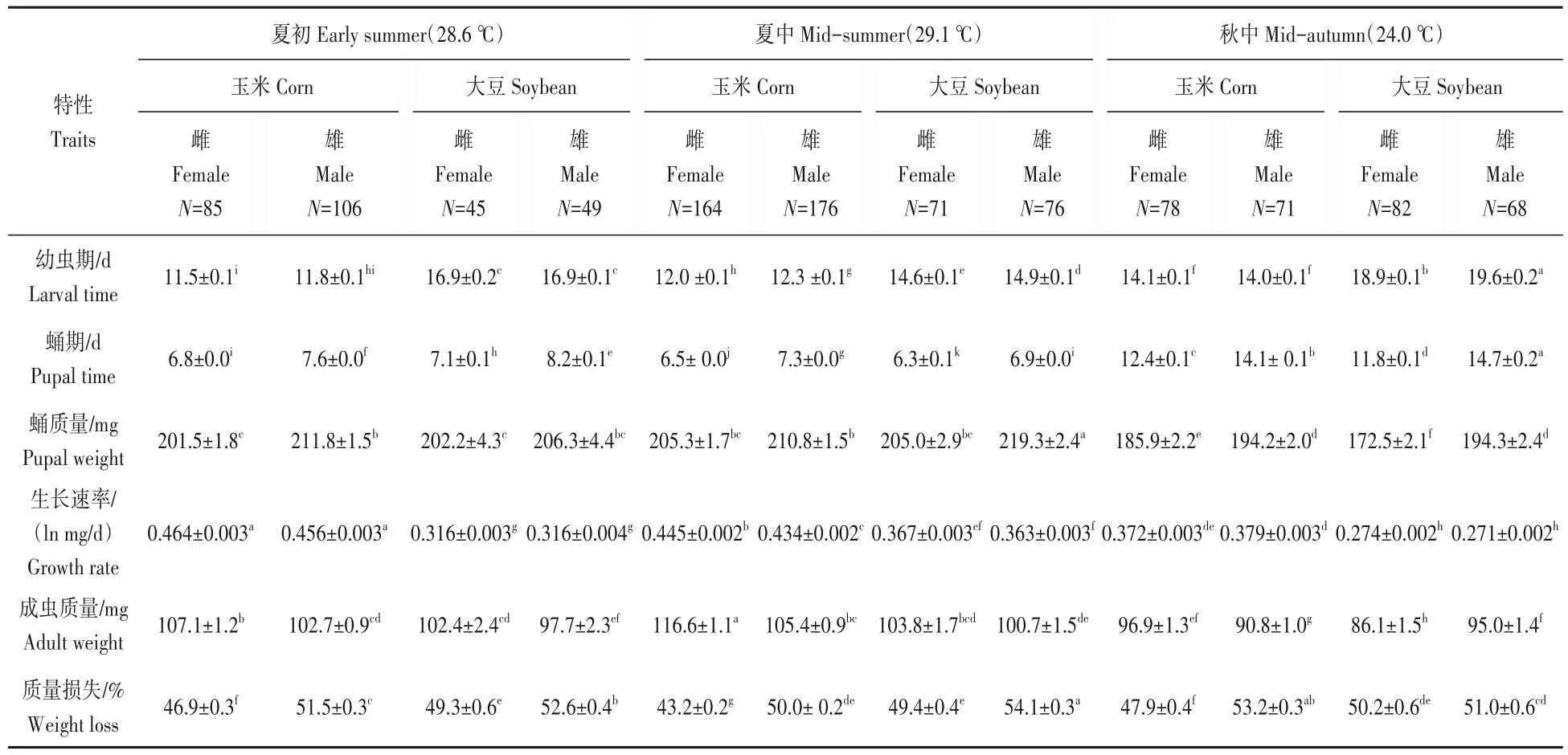
表2 草地贪夜蛾Spodoptera frugiperda在不同季节取食玉米叶和大豆叶的生活史资料Tab.2 Life history data(mean±SE.)for male and female S.frugiperda fed on the corn leaves and the soybean leaves at different seasons
2.2 与食料、季节和性别相关的蛹质量和生长速率
蛹质量受到季节和性别及它们互作(季节×性别)的显著影响;蛹质量没有受到食料的显著影响,但食料×季节、食料×性别和食料×季节×性别的互作显著影响了蛹质量(表1)。蛹质量在不同季节存在显著差异(F11,1059=31.606,P<0.05)(图2A)。在秋中中性温度下,取食玉米叶和大豆叶的蛹质量显著小于夏初和夏中高温时的蛹质量(P<0.05)。例如,秋中取食玉米叶和大豆叶的雌性蛹质量分别为185.9 mg 和172.5 mg,显著低于夏初的201.5 mg 和206.3 mg 及夏中的205.3 mg 和205.0 mg(表2)。在夏初和夏中,取食玉米叶的雌性蛹质量与取食大豆叶的雌性蛹质量没有显著差异(P>0.05);在秋中,取食玉米叶的雌性蛹质量显著高于取食大豆叶的雌性蛹质量(P<0.05),而取食玉米叶的雄性蛹质量与取食大豆叶的雄性蛹质量没有显著差异(P>0.05)。在各季节,雄性蛹质量均显著大于雌性蛹质量(P<0.05),显示了性体型二型性(sexual size dimorphism)。
生长速率受到食料、寄主和性别及它们的互作(食料×季节、季节×性别和食料×季节×性别)显著影响(表1)。取食玉米叶和大豆叶的生长速率在季节间存在显著差异(P<0.05),秋中取食玉米叶和大豆叶的生长速率显著低于在夏初和夏中时的生长速率(P<0.05)(图2B)。取食玉米叶的生长速率均显著高于取食大豆叶的生长速率(P<0.05)。在夏中取食玉米叶和大豆叶的生长速率雌性显著大于雄性(P<0.05),在秋中取食玉米叶的生长速率雄性显著大于雌性(P<0.05),其他处理的生长速率雌雄间没有显著差异(P>0.05)。
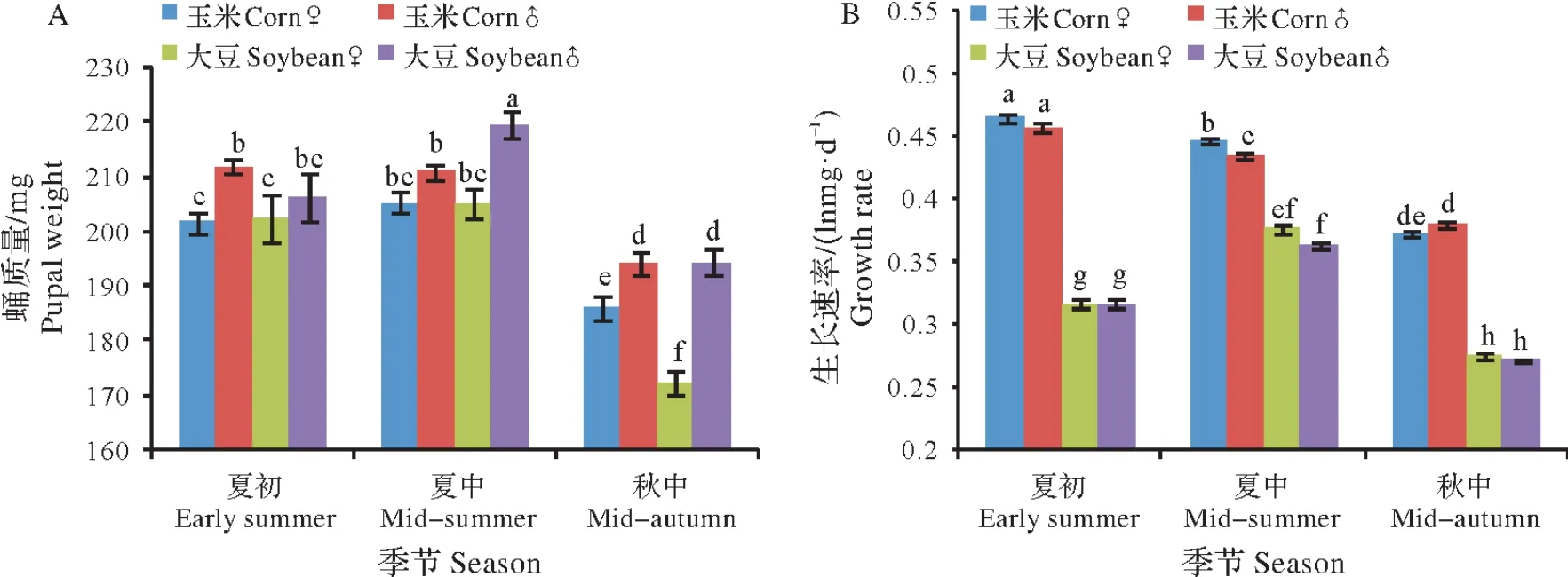
图2 草地贪夜蛾在夏初、夏中和秋中取食玉米叶和大豆叶的蛹质量和生长速率Fig.2 Pupal weight and larval growth rate of Spodoptera frugiperda feeding on corn leaves and soybean leaves in early summer,mid-summer and mid-autumn
2.3 与食料、季节和性别相关的成虫质量和失重比
食料、季节和性别及它们的互作显著影响了草地贪夜蛾的成虫体质量和失重比(表3)。从图3A可以看出,雌雄成虫质量在季节间存在显著差异(P<0.05),在秋中中性温度下,取食玉米叶和大豆叶的成虫质量显著小于夏初和夏中高温时的成虫质量(P<0.05)。在夏初和夏中,取食玉米叶的雌性成虫质量均显著高于取食大豆叶的雌性成虫(P<0.05),取食玉米叶的雄性成虫质量也显著高于取食大豆叶的雄性成虫(P<0.05);在秋中,取食玉米叶的雌性成虫质量均显著高于取食大豆叶的雌性成虫(P<0.05),而取食玉米叶的雄性成虫质量却显著低于取食大豆叶的雄性成虫(P<0.05)。除了在秋中取食大豆叶的雌性成虫质量低于雄性成虫外,其他所有的处理都是雌性成虫质量显著高于雄性成虫(P<0.05)。从图3B 还可以看出,无论是取食玉米叶还是大豆叶,雄性从蛹发育到成虫丢失的体质量比均显著大于雌性(P<0.05)。

图3 草地贪夜蛾在夏初、夏中和秋中取食玉米叶和大豆叶的的成虫质量和失重比Fig.3 Adult weight and proportionate weight loss of Spodoptera frugiperda feeding on corn leaves and soybean leaves in early summer,mid-summer and mid-autumn

表3 食料、季节和性别对草地贪夜蛾成虫质量和失重比影响的的多因素方差分析Tab.3 Multi-factor ANOVA analysis of food,season and sex on adult weight and weight loss rate in Spodoptera frugiperda
2.4 不同季节取食玉米叶和大豆叶的存活率和性比
表4表明,不管取食玉米叶还是大豆叶,幼虫和蛹的存活率均在90%以上。除在夏中高温下取食玉米叶的蛹存活率显著高于取食大豆叶的外(t=3.186,df=8,P<0.05),其他处理幼虫和蛹的存活率均未达到显著差异(P>0.05)。表4 还表明,性比没有受到季节和寄主植物的显著影响(P>0.05),在所有处理中性比均接近1∶1。

表4 草地贪夜蛾Spodoptera frugiperda在不同季节取食玉米叶和大豆叶的幼虫和蛹存活率及性比Tab.4 Larval and pupal survival rate and sex ratio of Spodoptera frugiperda fed on the corn leaves and the soybean leaves in different seasons
2.5 不同季节取食玉米叶和大豆叶的繁殖生物学
从表5 可以看出,草地贪夜蛾在不同季节取食玉米叶和大豆叶对雌虫的繁殖特性有显著影响。与取食大豆叶的草地贪夜蛾比较,玉米叶饲养的草地贪夜蛾在夏初、夏中和秋中均显示了较高的交配成功率(夏初达到显著水平:t=7.209,df=4,P<0.05)、较短的产卵前期(秋中和夏初下达到显著水平:秋中,t=-3.922,df=45,P<0.05;夏初,t=-2.930,df=46,P<0.05)、较高的产卵量(秋中和夏初达到显著水平:秋中,t=3.091,df=45,P<0.05;夏初,t=3.218,df=46,P<0.05)和显著高的孵化率(秋中:t=5.978,df=157.043,P<0.05;夏初:t=6.670,df=195,P<0.05;夏中:t=1.988,df=122.629,P<0.05)。取食玉米叶的交配成功率在夏初最高,显著高于夏中的交配成功率(F2,6=6.149,P<0.05);产卵前期、产卵期和寿命在季节间存在显著差异(产卵前期:玉米:F2,77=6.676,P<0.05;大豆:F2,49=14.047,P<0.05;产卵期:玉米:F2,77=38.049,P<0.05;大豆:F2,49=38.555,P<0.05;寿命:玉米:F2,77=43.632,P<0.05;大豆:F2,49=51.261,P<0.05)。取食玉米叶的产卵量在秋中最高(1 553 粒),显著高于夏中的产卵量(F2,77=15.503,P<0.05);取食大豆叶的产卵量亦在秋中最高(1 230 粒),显著高于夏初和夏中的产卵量(F2,49=6.250,P<0.05)。卵的孵化率均在秋中最高,显著高于夏初和夏中的孵化率(玉米:F2,335=110.327,P<0.05;大豆:F2,275=244.979,P<0.05)。
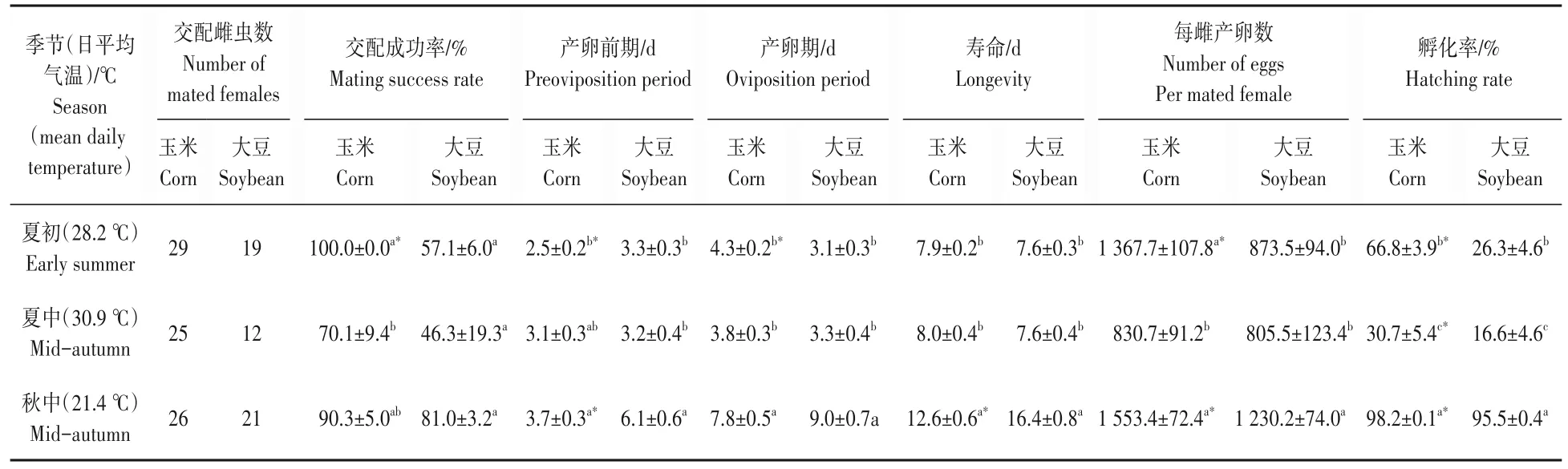
表5 草地贪夜蛾Spodoptera frugiperda在不同季节取食玉米叶和大豆叶雌成虫繁殖生物学Tab.5 Reproductive biology of mated females of Spodoptera frugiperda fed on the corn leaves and the soybean leaves in different seasons
3 结论与讨论
草地贪夜蛾入侵江西省两年以来,已成为江西玉米上的头号害虫。田间调查发现,在没有采取任何防治措施的玉米地,为害最严重的田块其玉米植株100%受害。该虫对玉米的为害是毁灭性的,已对江西的玉米生产构成了严重威胁。田间调查也表明,草地贪夜蛾在江西几乎全部发生在玉米上为害,少量发生在甘蔗上为害,这与草地贪夜蛾侵入我国后多在玉米地暴发为害是一致的[27-28]。研究亦表明,玉米为我国新入侵草地贪夜蛾的最佳寄主植物[18-19,29-30]。本研究系统比较了3 个不同季节草地贪夜蛾取食玉米叶和大豆叶的生活史特性,进一步证实了玉米是草地贪夜蛾最嗜好的寄主。与取食大豆叶的草地贪夜蛾比较,取食玉米叶的草地贪夜蛾显示了显著短的幼虫发育历期,显著高的生长速率、较高的交配成功率、较短的产卵前期、较高的产卵量和显著高的卵孵化率。此研究结果与He等[30]的一些研究结果一致。然而,尽管草地贪夜蛾在大豆上的适合度不如在玉米上那么好,但以大豆叶饲养的草地贪夜蛾能顺利完成个体发育,幼虫和蛹的存活率均在90%以上,与以玉米叶饲养的草地贪夜蛾没有显著差异;以大豆叶饲养的草地贪夜蛾亦表现出较高的繁殖力。研究表明,植食性昆虫可通过植物挥发物等次生代谢物来寻找和定位寄主植物[31]。例如,对马铃薯甲虫Leptinotarsa decem⁃lineata来说,只要有马铃薯叶片气味存在,马铃薯甲虫就会产生寄主定向行为[32];而许多植物挥发性气味成分中的苯乙醛对棉铃虫Helicoverpa armigera、亚洲玉米螟Ostrinia furnacalis、粉纹夜蛾Trichoplu⁃sia ni等多种蛾类具有引诱作用;多种果树挥发物中的一种法尼烯成分对苹果蠢蛾Cydia pomonella具有较强的引诱作用;杨树叶的挥发物则对雌性棉铃虫有很强的引诱作用[33]。目前在江西没有发现草地贪夜蛾为害大豆,可能是大豆叶中的挥发性气味对入侵的草地贪夜蛾不具备引诱作用。但随着草地贪夜蛾入侵时间的增长,暴发频率的增加,当玉米食物匮乏时,可能转向为害大豆,最终形成适应大豆的草地贪夜蛾种群。因为昆虫的寄主转移与进化适应是种群分化的重要机制,桃蚜Myzus persi⁃cae取食烟草后逐渐形成了适应烟草的种群就是一个典型的例证[34]。因此,今后必需密切关注草地贪夜蛾在大豆田的发生动态。
本研究首次发现在不同季节取食玉米和大豆叶片的草地贪夜蛾,其雄蛹发育历期显著长于雌蛹,导致雌性先羽化的雌性先熟现象,何莉梅等[35]在25 ℃下也观察到取食花生的草地贪夜蛾的雄蛹发育历期显著长于雌蛹。这种雌性先熟现象已在一些鳞翅目昆虫物种中得到证实,例如迁飞性害虫斜纹夜蛾Spodoptera litura[36]、棉铃虫H.armigera[37-38]和小菜蛾Plutella xylostella[39],但出现这种现象的原因还不清楚。它可能代表一种进化策略,以促进不同种群个体之间的交配[39-40],也可能是一种减少近亲繁殖的策略[36]。本研究还发现草地贪夜蛾的雄蛹质量显著大于雌蛹质量(P<0.05),显示了性体型二型性,这与吴正伟等[20]的报道一致。然而草地贪夜蛾的成虫体质量,除了在秋中取食大豆叶的雌性成虫质量小于雄性成虫外,其他所有的处理都是雌性成虫质量显著大于雄性成虫(P<0.05)。造成这种差异的主要原因是因为雄性从蛹发育到成虫丢失了更多质量所致。这一结果表明了性别间失重比的差异对昆虫性体型二型性起到了重要的调节作用[41]。雌蛹在变态过程中丢失的质量少于雄蛹应该是一种进化的适应,因为雌性更趋向通过增加体质量来提高繁殖力[42-43]。
本研究还首次揭示了以玉米叶和大豆叶饲养草地贪夜蛾均在高温的夏初和夏中获得了更高的蛹质量和成虫质量,没有遵循温度-体型大小法则。究其原因可能是入侵的草地贪夜蛾的体型在原产地长期经历了高温选择所致。这种反温度-体型大小法则(the reverse temperature-size rule)也出现在红萝卜甲虫Entomoscelis americana[44]、4 种水生的蜉蝣昆虫[45]、4 种英国蚱蜢[46]、热带蝴蝶Bicyclus anynana[47]、有迁飞能力的小菜粉蝶Pieris rapae[48]、亚洲玉米螟Ostrinia furnacalis[49-51]和二化螟Chilo suppressalis[52-53]中,这些昆虫种类均显示了其体型或蛹质量随着温度的升高而增加或在较高的温度下成熟的个体体型更大。
需提及,相对于9 月4 日孵化在中性温度(24.0 ℃)下生长发育到成虫的个体,6 月14 日和7 月7 日孵化在高温度下(28.6 ℃和29.1 ℃)生长发育到成虫的个体显示了明显短的发育历期和获得了更大的体质量,但是在这两种高温下成虫的产卵量和孵化率却显著降低,是什么原因导致的呢?9 月1 日从野外玉米上采回3 头雌成虫置于室内产卵,每雌平均产卵量达(1 343.0±67.8)粒,卵孵化率为(97.7±0.5)%。根据推算,这3 头雌虫幼虫期所经历的日平均气温为30.6 ℃,蛹期所经历的温度为28.9 ℃。因此推测在高温下将刚羽化雌雄成虫置于带有用10%蜂蜜水湿润的棉球的充入空气的密封塑料袋中配对可能对成虫的交配、受精和产卵有不利影响。如将配对成虫置于通风透气的纱网笼中交配,雌虫的产卵量和卵孵化率应该有较大提高。为此,下一步的研究工作将通过改善高温下的交配环境提高成虫产卵量和卵孵化率。
