不同光境下多花黄精光合特性及形态结构比较
2021-11-05贺安娜伍贤进李胜华何述金
贺安娜,伍贤进※,李胜华,何述金
(1.怀化学院 生物与食品工程学院,民族药用植物资源研究与利用湖南省重点实验室,湘西药用植物与民族植物学湖南省高校重点实验室,湖南 怀化 418008;2.湖南新汇制药股份有限公司,湖南 长沙 410200)
多花黄精(PolygonatumcyrtonemaHua)为百合科黄精属多年生草本植物,具补气养阴、健脾、润肺、益肾等功效,为传统大宗珍贵中药材[1],也是国家批准的药食同源植物,其肥厚的根状茎富含黄精多糖、类固醇皂苷、黄酮苷等化合物[2],具有降血糖、降血脂、提高免疫力的作用[3],在保健品及功能性食品开发等方面具有广阔的发展前景。
随着多花黄精药用特别是食用的不断增加,野生资源正日益枯竭,人工种植则方兴未艾。光照条件是植物生长发育最重要的外部条件,对栽培多花黄精光合生理特性研究表明,多花黄精不耐强光照射,人工种植需拉遮阳网,遮光率40%~50%时光合速率最高[4-8]。野生的多花黄精常分布于林下及林缘,喜阴湿的环境,林下种植是其非常合适的栽培模式[9]。研究表明,多花黄精对光强的适应能力非常强,从全光照至郁闭度90%的林下都能生长发育[10]。这些研究结果显示,人工种植多花黄精,光照强度的选择仍然需要进一步论证。而且,植物生长发育不仅受光照强度影响,光质的影响也不可忽视[11-12]。这是因为植物叶绿体色素对光的吸收集中在可见光的蓝色(400~500 nm)和红色(600~700 nm)区域,相同的光强下,光质不同可引起叶绿素含量及比例发生变化[13],植物形态建成也受光质的影响,紫外光抑制茎的生长,使植株变得矮小,红光则能促进茎的生长,增加地上部分的生物产量[14]。人工遮阴可以改变光照强度,但基本不能改变光质。相反,林下不仅光照强度较低,而且光质也因上层树叶对大部分红光和部分蓝光的吸收造就了与遮阳网不同的光质。但目前对不同光环境下,多花黄精生理特性和生长发育所受的影响还缺乏足够的研究。为此,本文通过对不同光照环境下多花黄精生理特性的研究,可为多花黄精的人工栽培选择适合的光环境提供理论和实践指导,也可为类似喜阴作物栽培提供理论参考依据。
1 材料与方法
1.1 实验材料及处理
2019年12月,挑选大小一致、无病虫害的多花黄精种茎种植于塑料花盆内,每盆种植一株,用土量保持一致。花盆分别置于全日照下(100%透光率)、香樟树下(40%透光率)及遮阳网致的阴网棚下(40%透光率),每种处理放置15盆,三种处理的直线距离不超过100 m,处理期间保证一致的浇水和施肥量。2020年8月待多花黄精进入果熟期时进行测定。
1.2 方法
1.2.1 气体交换参数及叶绿素荧光参数测定
选择晴朗的天气,随机选取生长正常的多花黄精中段叶片,于8∶30—10∶00使用Li-6400便携式光合仪(Li-COR,USA)对瞬时气体交换参数进行测定。光照、二氧化碳浓度等均采用自然条件,记录净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)等参数。每组处理测定3株约10片叶。
用锡箔纸包裹好待测的中段叶,使其暗适应30 min后,用Li-6400便携式光合仪的荧光探头测定原初荧光(Fo)及暗中最大荧光(Fm)。打开叶室,让叶片在光下活化30 min后,测定测稳态荧光(Fs)、最大荧光(F´m)及最小荧光(F´o)。计算光化学猝灭系数:qP=(Fm'-Fs)/(Fm'-Fo'),非光化学猝灭系数:qN=(Fm-Fm')/(Fm-Fo'),实际光化学效率:ΦPSⅡ=(Fm'-Fs)/Fm',电子传递速率:ETR=ΦPSⅡ×0.5×0.84×400。每组处理测定10片叶。
1.2.2 叶绿素含量测定
取健康的中段叶片,去除主脉称量0.2 g,用95%的乙醇提取并定容至25 ml,分别在665,649,470 nm波长下测定光吸收值。叶绿体色素的含量(mg/g·FW)=(C×V×N)/m×1 000,式中:C为色素浓度(mg/L),V为提取液体积(25 ml),N为稀释倍数,m为样品质量(0.2 g左右),1000是表示1 L=1 000 ml。每种处理重复3次。
1.2.3 叶片显微结构观察
选取健康的中段叶,FAA固定液固定24 h以上。叶横切观察采用石蜡切片的方法,经染色、脱水、透明、浸蜡、包埋、切片、脱蜡等程序后,封片拍照;叶表皮结构的观察采用徒手切片的方法,制作临时装片拍照。采用Olympus BX-60型光学显微镜观察叶片显微结构并拍照,采用PhotoShop13.0.1图片处理软件进行图片合并及有关数据的测量,每组处理重复3~5次。
1.2.4 株叶形态及生物产量测定
用软尺测株高、叶长、叶宽,用游标卡尺测茎粗(茎基部离地面1 cm处的直径)、用Li-3000C叶面积仪(Li-COR,USA)测定单株的功能叶面积。每组处理取15株。
材料处理完毕后,挖出地下部分洗净,测定地上、地下部分鲜重,计算根冠比。黄精多糖含量按照《中华人民共和国药典》规定的方法测定[1]。
2 结果与分析
2.1 不同光境下多花黄精叶片气体交换参数比较
表1表明,8月中旬多花黄精地上部分的生长进入缓慢期,各光境下的净光合速率(Pn)都较低,Pn最高的为香樟树下处理组,为4.02μmol·m-2·s-1,全光照下最低。气孔导度及蒸腾速率也是香樟树下的最高,也正是因为有较高的蒸腾速率,香樟树下的水分利用率(WUE)最低。阴网棚下处理的多花黄精叶片气孔导度(Gs)及胞间二氧化碳浓度(Ci)显著低于其他两项处理,说明此时光合速率受气孔因子限制,气孔关闭也使蒸腾速率(Tr)维持在较低的水平。
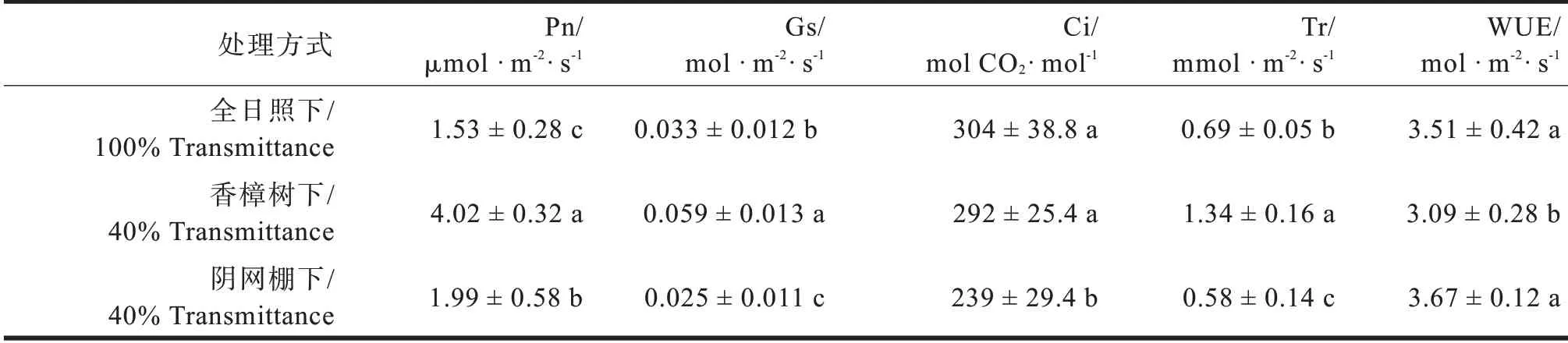
表1 不同光境下多花黄精叶片气体交换参数比较
2.2 不同光境下多花黄精叶片叶绿素荧光参数及叶绿素含量比较
表2显示,三种光境处理下的多花黄精叶片Fv/Fm及Fv/Fo没有显著差异,说明三种光境下叶片光系统Ⅱ反应中心原初光能转换效率及从Cha/b蛋白复合体LHCP到光系统Ⅱ的光能传递能力没有显著差异,但香樟树下的非光化学淬灭(qN)最低,且实际光化学反应速率(ΦPSⅡ)和电子传递效率(ETR)最高,且与另两种处理达到显著水平。因此,香樟林下叶片对光能的利用能力较强。

表2 不同光境下多花黄精叶片叶绿素荧光参数比较
从表3结果看,香樟树下的多花黄精叶片叶绿素a含量高于阴网棚下的处理,叶绿素b则低于阴网棚下的,但差异均未达显著水平。类胡萝卜素能抗光氧化,有利于抵御不良环境,该含量以全日照下最高,香樟林下次之,阴网棚下最低且与全日照和香樟林下有显著差异。
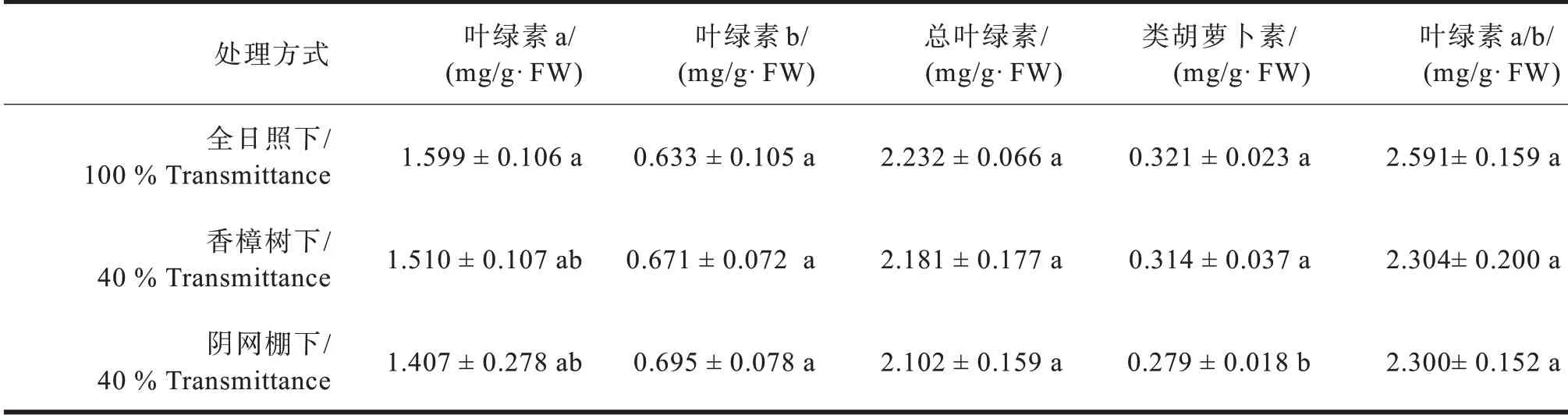
表3 不同光境下多花黄精叶片叶绿素含量比较
2.3 不同光境下多花黄精叶片显微结构比较
多花黄精叶没有明显栅栏组织与海绵组织之分,属等面叶(见图1)。全日照和香樟树下的叶上表皮排列整齐,紧靠上表皮的叶肉细胞呈椭球型,排列也较紧密,阴网棚下的叶片叶肉细胞排列则较疏松,全日照和香樟树下的叶片较厚,叶脉明显,维管组织发达。
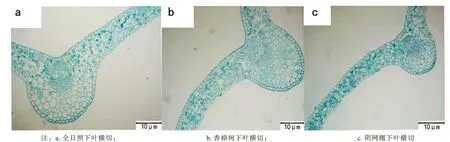
图1 不同光境下多花黄精叶横切
多花黄精叶片上表皮细胞呈长方形,排列紧密,未见气孔分布;下表皮细胞呈不规则型,气孔均匀的分布在下表皮,未见护卫细胞(见图2)。多花黄精叶表皮的显微结构在不同光境下有差异,表现为香樟树下的上表皮细胞最长、最宽;气孔密度最高是全日照下的,有94.3个/mm2,气孔指数也显著高于其他两项处理;阴网棚下叶的气孔密度最低,仅57.4个/mm2(见表4)。

图2 不同光境下多花黄精叶表皮结构
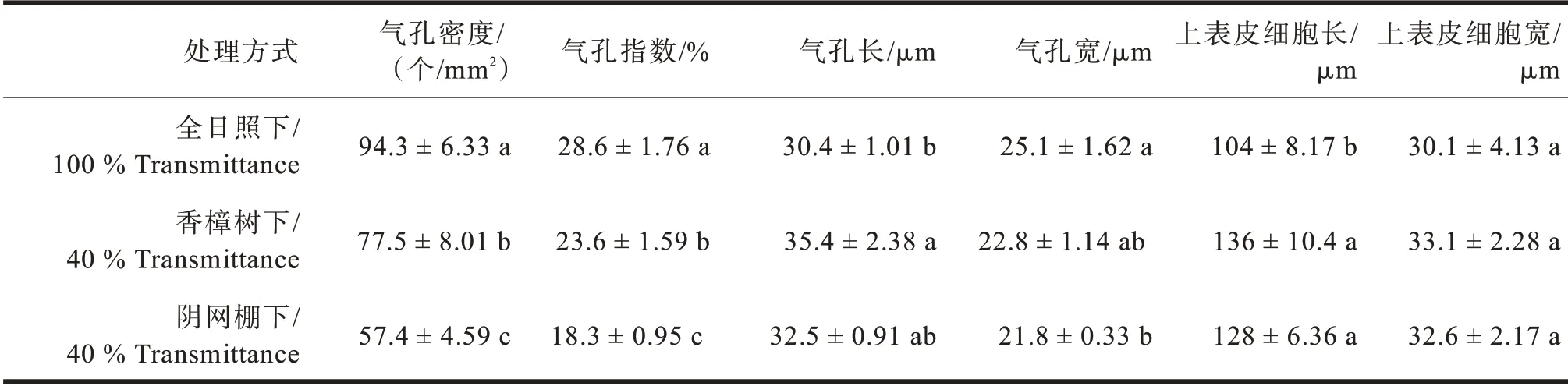
表4 不同光境下多花黄精叶表皮细胞特征
2.4 不同光境下多花黄精株叶形态及生物产量比较
多花黄精的株高在不同光境条件下存在显著差异(见表5),阴网棚下最高,可达85.4 cm,全日照下最低,只有49.1 cm;茎粗、叶长、叶宽等以阴网棚下最高,但三种处理之间无显著差异;单叶叶面积以全日照最低,且与阴网棚和香樟林下有显著差异。
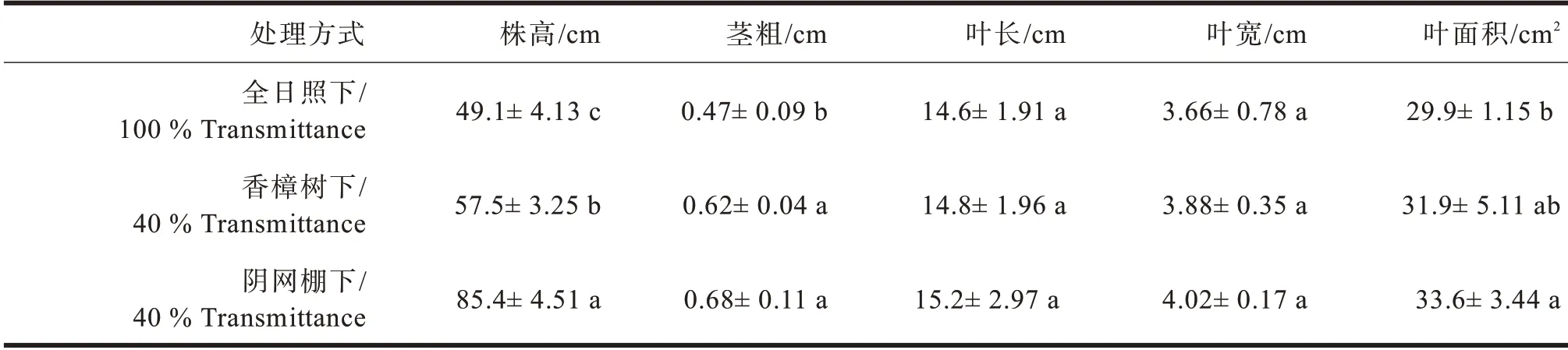
表5 不同光境下多花黄精株叶形态比较
不同光境下多花黄精的生物量也有很大差异(见表6),全日照条件下单株鲜重最低,仅73.9 g,但根冠比最高,说明强光照有利于地下茎的积累;香樟树下的单株鲜重最高,多糖含量也最高,阴棚内的单株鲜重、根冠比适中,但多糖含量最低。
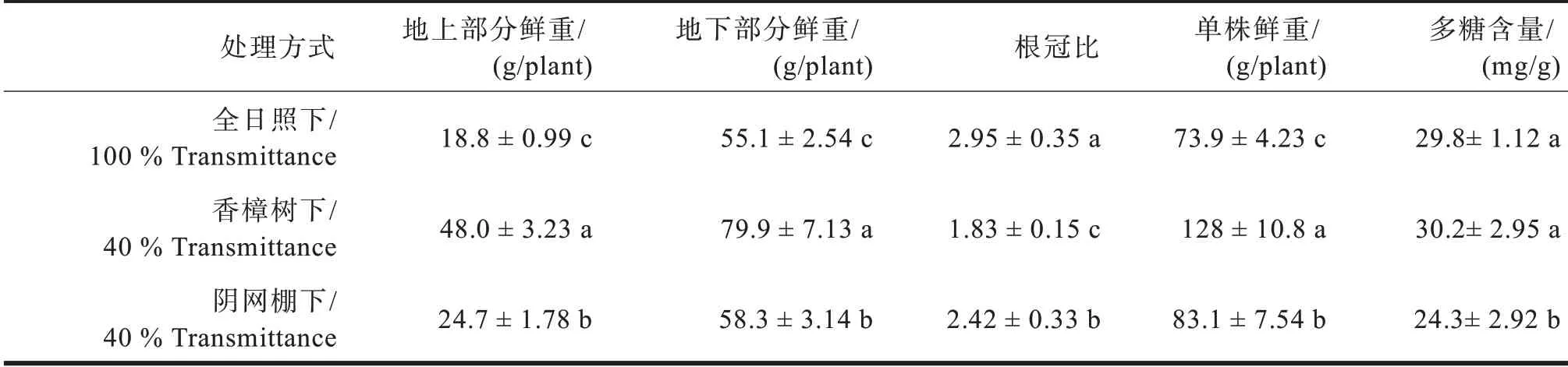
表6 不同光境下多花黄精生物产量比较
3 讨论
光合作用是为地球生命提供动力的绿色引擎,能直接影响植物的产量及品质。实验中,8月下旬多花黄精进入果熟期,三种光照条件下净光合速率都不高,尤其是全日照条件下的净光合速率仅1.53μmol·m-2·s-1,说明全日照下的多花黄精衰老速度最快,适当遮阴有利于延长多花黄精叶光合功能的持续时间,有利于促进生长,这与生物量差异也很一致。相同的光强下,不同的光质对植物光合作用的影响不同,在能量较高的蓝光照射下,植物的非光化学淬灭NPQ更高[15]。与全光照相比,香樟树下的散射光和蓝紫光更多,该光境下多花黄精叶片的非光化学淬灭(qN)最低,叶片实际光化学反应速率(ΦPSⅡ)和电子传递效率(ETR)最高,说明多花黄精对这类高能的蓝紫光利用能力较强。三种光境下,这些光合作用有关参数的变化与他们生物量的积累也是一致的。叶绿素是光能转化的关键,其含量的适当增加有利于捕捉光量子,总叶绿素含量的增加与PSI的易感性有关[16],多花黄精在三种光境下,总叶绿素含量没有显著差异,表明这些光境条件下叶绿素的含量都能满足多花黄精光量子的捕捉。类胡萝卜素是一种重要的抗氧化剂,全光照下的高含量是植物适应环境的表现,能减少强光对叶片造成的损伤。
决定光合作用功能的叶片结构在不同光境下,也会作出适应性的改变,如蓝光可使油菜叶片栅栏组织排列更紧凑[17]。实验中,全日照和香樟树下的多花黄精叶片细胞排列紧密,维管组织发达,气孔密度、气孔指数也较大,且三组处理之间的差异都达显著水平,说明光照黄精对多花黄精也的发育产生了实质性影响。香樟树下与阴网棚下的光强虽然一样,但香樟树下的多花黄精叶片细胞排列更紧密,形成更多的气孔以适应高能的蓝紫光环境。这些结果说明,香樟树下虽然光照强度降低了,但因光质的改变,其叶的结构发育仍然能达到较强光照的程度,而单纯通过遮阳网降低光强,其光环境不如林下更适合多花黄精生长。
光照强度减少能促使植物株高增加,相同光照强度下,林下比遮阳网下光的蓝紫光成分更多,也更利于黄精这类阴生植物的生长发育。因此,阴网棚下的多花黄精株高虽然最高,但茎粗、叶长、叶宽和叶面积与香樟林下并无显著差异,而且从叶的细胞组织结构看,香樟林下细胞组织更致密,故香樟林下地上部分鲜重比阴网棚下更高。不仅是地上部分鲜重,香樟林下地下部分鲜重也是三个处理中最高的。这些与香樟林下净光合速率高的数据一致,也说明香樟林下更适合光合产物在黄精地下部分积累。总之,多花黄精为等面叶,属阴生植物,能有效利用短波长的蓝紫光,林下种植多花黄精优于遮阳网遮阳处理。为了提高遮阳网下种植黄精的产量和品质,可考虑增加短波长蓝紫光的辅助设施。
